
瘤胃内容物发酵对豆粕蛋白质品质及菌群组成的影响
2011-07-12 00:32:38浙江万里学院生物与环境学院胡程珩李彩燕杨义右黄丽萍
中国饲料 2011年18期
浙江万里学院生物与环境学院 胡程珩 吕 叶 李彩燕* 郭 宇 杨义右 黄丽萍
豆粕是一种优质的植物蛋白质源,价格低且来源广,是畜牧水产饲料中理想的鱼粉替代物。但豆粕中存在多种抗营养因子,如胰蛋白酶抑制剂、植酸等(Francis等,2001),极大地影响了豆粕的饲用价值,需要经过加工处理才能广泛应用。近年来,与加热处理、化学浸提等常规方法相比,微生物发酵处理由于去除效果良好、营养价值提高、工艺环保等多种优点,备受关注。以微生物发酵的方法来降低植物蛋白质抗营养因子的研究已有不少报道 (Hong 等,2004;Egounlety 和 Aworh,2003;El-Batal和 Karem,2001)。
反刍动物的瘤胃是一种特殊的发酵罐,其中栖息着各种微生物(细菌、真菌、原虫等),表现出很强的降解纤维素和消除饲料毒物的作用,包括生氰糖甙、皂甙和植酸等 (赵智力和王思珍,2008)。国内外对瘤胃微生物的研究主要来自反刍动物营养学领域,对其在蛋白质饲料开发方面的报道很少。笔者所在的课题组前期已通过单因素和正交试验,优化了实验室瘤胃液发酵豆粕的条件,确定了最佳发酵模式,使发酵豆粕中主要抗营养因子得到有效去除(李彩燕等,2011)。
本试验在最适条件下用瘤胃液对豆粕进行了发酵,测定了发酵前后豆粕的蛋白质品质(氨基酸含量和蛋白质降解情况)、微生物组成(细菌总数、乳酸菌、大肠杆菌、霉菌)及pH值,以证实瘤胃液发酵豆粕的益处,可进一步为工业化生产发酵植物蛋白质提供参考。
1 材料与方法
1.1 试验材料
1.1.1 豆粕原料与瘤胃液 豆粕原料购自宁波市鄞州区钟公庙集贸市场。称取同批豆粕粉碎,过40目筛,充分混匀,干燥保存备用。瘤胃液取自健康瘘管湖羊瘤胃,使用前用四层纱布过滤。
1.1.2 主要培养基 营养琼脂培养基 (PCA)、马铃薯葡萄糖琼脂培养基(PDA)、乳酸细菌培养基(MRS)、月桂基硫酸盐胰蛋白质胨(LST)、煌绿乳糖胆盐肉汤(BGLB),均购自杭州微生物试剂有限公司。
1.1.3 主要仪器设备 生化培养箱(GZX,宁波东南仪器有限公司)、酸度计 (梅特勒-托利多FE20)、微型发酵瓶(三角瓶)、凯氏定氮仪、培养皿、卤素水分测定仪(MB35)、日立L-8900氨基酸自动分析仪。
1.2 试验方法
1.2.1 体外发酵技术 参考Menke体外产气法(Menke等,1998),利用瘤胃液中的微生物对豆粕进行厌氧发酵处理。瘤胃液接种量为5%,料水比为 1∶1,39 ℃厌氧发酵 72 h(李彩燕等,2010)。
1.2.2 氨基酸含量的测定 发酵前后豆粕样品经盐酸水解后,氨基酸含量用氨基酸自动分析仪进行测定。
1.2.3 豆粕蛋白质的电泳分析 参考马文强等(2008)和 Hoffmann 等(2003)方法。称取粉碎后过60目的豆粕1.00 g,加入20.00 mL 0.03 mol/L Tris-HCl(pH 8.0),在室温下于摇床上 100 r/min浸泡 1 h;然后 10000 r/min,20 ℃离心 20 min,取上清。提取的蛋白质进行SDS-PAGE电泳。
1.2.4 样品pH值的测定 参考那淑敏等(1999)方法。豆粕发酵前后的样品烘干处理后各称取10 g,用 100 mL蒸馏水混合浸提2 min,经四层纱布过滤,使用酸度计测定水溶液的pH值。
1.2.5 样品菌群的测定 分别取发酵前后的风干豆粕样品1 g,加入有9 mL无菌水的试管中,涡旋器混匀,再稀释至 10-2、10-3、10-4、10-5、10-6浓度梯度,每个梯度分别重复3次,按照以下方法进行操作。细菌菌落总数的测定按GB/T4789.2-2008执行;大肠菌群的测定按GB/T4789.3-2008执行(分别采用MPN计数和平板计数法);乳酸菌测定按GB/T4789.35-2008执行;霉菌测定按 GB/T4789.15-2008执行。
1.3 数据统计 数据经Excel初步处理后,采用SAS(1999)广义线性模型(GLM)的单因素方差分析进行统计分析法和显著性检验,平均数之间用Duncan’s法进行多重比较。
2 结果
2.1 发酵前后氨基酸含量的变化 瘤胃微生物发酵前后豆粕中氨基酸含量变化见表1。由表1可知,氨基酸总量由发酵前得47.25%提高到发酵后的48.67%,比发酵前提高了3%,绝大多数氨基酸的含量都有所提高。
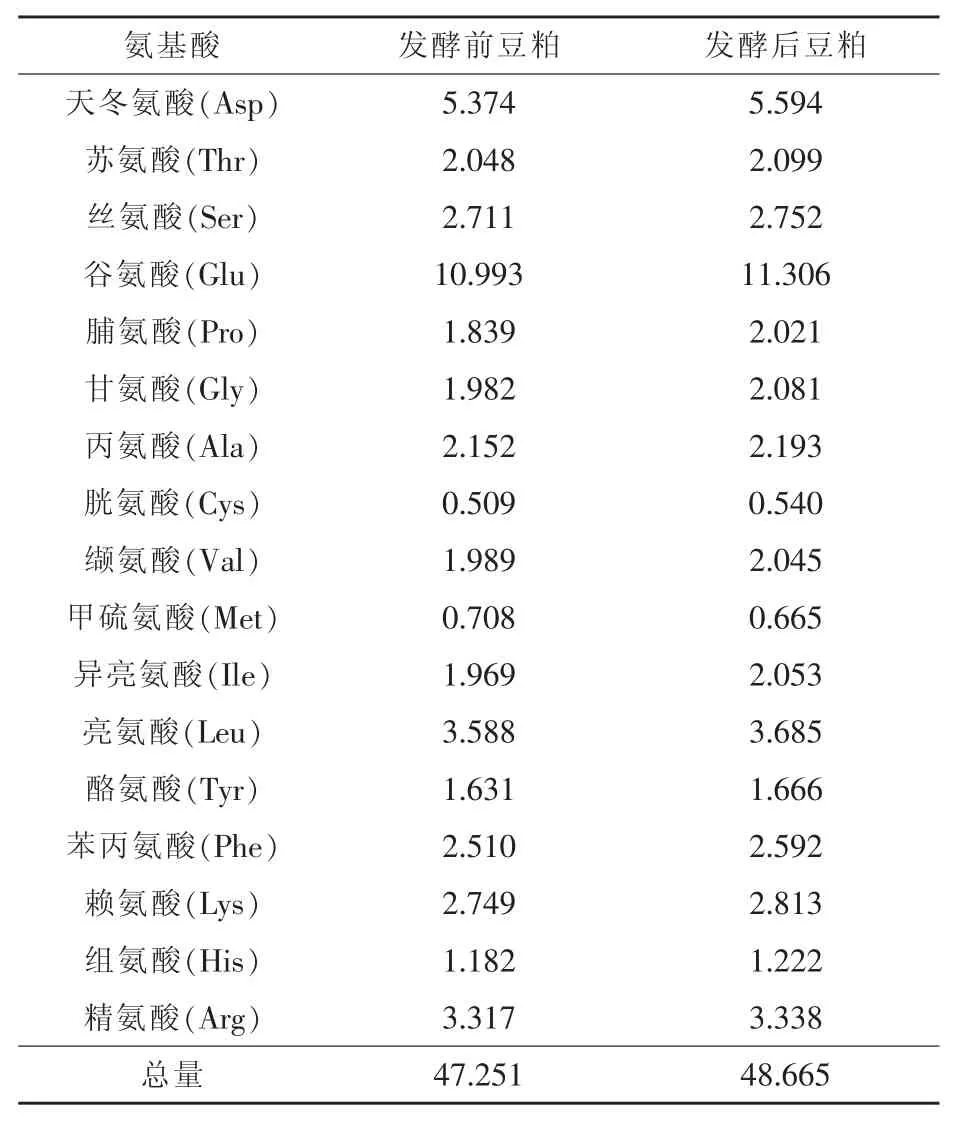
表1 发酵前后豆粕中氨基酸含量的变化%
2.2 发酵豆粕蛋白质的电泳分析结果 对发酵前后的豆粕蛋白质进行SDS-PAGE电泳可得到图1的结果,可见,大分子质量的蛋白质得到降解,分子质量大于31 kDa的蛋白质明显减少,小分子多肽明显增加。
2.3 发酵前后豆粕中pH值和菌群结构的变化瘤胃内容物发酵对豆粕pH值和菌群结构的影响结果见表2。由表2可知,豆粕经瘤胃内容物发酵后pH值显著下降,降低了1.32个单位。
细菌总数显著增加,增加了2个数量级,霉菌和大肠杆菌均显著降低,分别降低了3个和1个数量级。而有益的乳酸菌显著上升,上升了4个数量级。

图1 发酵前后豆粕中的蛋白质电泳图

表2 发酵前后豆粕pH值和微生物指标测定结果
3 讨论
3.1 发酵对豆粕氨基酸组成和蛋白质肽的影响豆粕经瘤胃内容物发酵后,胰蛋白质酶抑制因子和植酸等主要抗营养因子含量大大降低,营养特性得到明显改善(李彩燕等,2011)。本试验期望通过瘤胃微生物对豆粕的发酵,微生物大量繁殖,把部分原料转化为菌体蛋白质,改善蛋白质氨基酸组成,还能积累一些有益代谢产物。试验结果发现,豆粕发酵后,氨基酸含量也升高,与笔者前期对营养特性的研究结论一致。马文强等(2008)通过枯草芽孢杆菌、酿酒酵母菌和乳酸菌对豆粕进行发酵后,其氨基酸的含量比发酵前提高了11.49%。同时本试验对发酵豆粕进行电泳得出,发酵前大分子蛋白质已被降解为小分子质量的肽类,这与马文强等(2008)的结果相一致。发酵后豆粕小肽含量的增加可能是由于发酵过程中微生物分泌的蛋白质酶分解部分大分子蛋白质得到的。蛋白质被微生物分解后,生成动物易吸收的小分子蛋白质或小肽,以提高豆粕利用率与蛋白质的吸收率(马文强等,2008)。
3.2 发酵对豆粕菌群组成的影响 乳酸菌属于有益的肠道微生态调节剂,可对动物肠道微生态平衡进行调整,达到帮助饲料消化吸收,提高饲料利用率,抑制肠道病原菌等多方面的作用 (江萍等,1998)。豆粕经瘤胃液作用发酵后,菌落总数和乳酸菌数量明显增加。这与贾朋辉等(2009)微生态发酵饲料中细菌总数和乳酸菌的变化趋势相一致,虽然采用不同的发酵菌种,本试验中使用瘤胃液,贾朋辉等(2009)使用乳酸菌进行发酵,但是均说明了发酵能促进微生物的大量增殖。此外,由于微生物发酵糖类产酸和厌氧程度的逐渐加强,使物料环境更有利于乳酸菌的增殖。
仅用菌落总数不能作为评价发酵豆粕质量好坏的指标(刘金萍和王士长,2006),需要结合乳酸菌、大肠杆菌、霉菌等微生物指标的变化。大肠杆菌主要来源于人畜粪便,有些是致病菌或条件致病菌,常被用来作为评价食品及饲料的卫生质量指标(贾朋辉等,2009)。本试验中,豆粕发酵后,菌落总数和乳酸菌明显增加的同时,大肠菌群和霉菌数量下降,表示其有害菌的数量下降。
3.3 发酵对豆粕pH值的影响 从表1可以看出,豆粕经过瘤胃液发酵后,pH值显著降低。可能由于发酵后,豆粕中乳酸菌大量增殖,在生长过程中同时产生乳酸,使饲料的pH值降低。刘金萍和王士长(2006)应用植物乳杆菌发酵断奶仔猪料也发现,饲料经过发酵后,pH值都有所降低,最终pH值与乳酸菌含量有一定的关系。
4 小结
豆粕经瘤胃内容物发酵后,氨基酸含量提高,大分子质量的蛋白质得到降解,小分子多肽明显增加,并且显著增加了有益的乳酸菌含量,降低了有害的霉菌和大肠菌群的数量。本试验利用瘤胃微生物较强的消化去毒作用,证实了瘤胃液可作为豆粕发酵处理的有效菌源,为开辟新的植物蛋白质饲料资源进行了有益的探索。
[1]江萍,夏先林,秦礼康,等.魔芋飞粉基质生料发酵[J].饲料研究,1998,9:3~6.
[2]贾朋辉,郭烘新,李国军,等.微生态发酵饲料菌群变化及其应用[J].饲料博览,2009,3:24 ~ 27.
[3]刘金萍,王士长.植物乳杆菌N3发酵断奶仔猪料对饲料pH值及菌群的影响[J].2006,27(16):34 ~ 36.
[4]李彩燕,钱国英,汪财生,等.瘤胃微生物降解豆粕抗营养因子的发酵条件的优化研究[J].中国粮油学报,2011,26(6):73 ~ 77.
[5]马文强,冯杰,刘欣.微生物发酵豆粕营养特性研究[J].中国粮油学报,2008,23(1):121 ~ 124.
[6]那淑敏,贾士芳,陈秀珠,等.嗜酸乳杆菌发酵代谢产物分析[J].中国微生态学杂志,1999,11(5):266 ~ 268.
[7]赵智力,王思珍.反刍动物瘤胃微生物对饲料毒物的作用[J].中国动物保健,2008,112:64 ~ 65.
[8]Egounlety M,Aworh O C.Effect of soaking,dehulling,cooking and fermentation with Rhizopus oligosporus on the oligosaccharides,trypsin inhibitor,phytic acid and tannins of soybean (Glycine max Merr.),cowpea (Vigna unguiculata L.Walp) and groundbean (Macrotyloma geocarpa Harms)[J].Journal of Food Engineering,2003,56(2 ~ 3):249 ~ 254.
[9]El-Batal A I,Karem H A.Phytase production and phytic acid reduction in rapeseed meal by Aspergillus niger during solid state fermentation[J].Food Research International,2001,34(8):715 ~ 720.
[10]Francis G,Makkar H P S,Becker A.Antinutritional factors present in plant-derieved alternate fish feed ingredients and their effects in fish[J].Aquaculture,2001,199: 197 ~ 227.
[11]Hong K J,Lee C H,Kim S W.Aspergillus oryzae GB-107 Fermentation improves nutritional quality of food soybeans and feed soybean meals[J].Journal of Medicinal Food,2004,7(4): 430 ~ 435.
[12]Hoffmann E M,Muetzel S,Becker K.The fermentation of soybean meal by rumen microbes in vitro reveals different kinetic features for the inactivation and the degradation of trypsin inhibitor protein[J].Animal Feed Science and Technology,2003,106:189 ~ 197.
[13]SAS.Procedures guide for personal computers,Version 8.0[M].SAS Institute Inc,Cary,NC,USA.1999.
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23 06:23:18
中国饲料(2022年5期)2022-04-26 13:42:34
当代水产(2019年11期)2019-12-23 09:03:54
当代水产(2019年7期)2019-09-03 01:02:18
当代水产(2019年6期)2019-07-25 07:52:10
中成药(2018年10期)2018-10-26 03:41:28
河南畜牧兽医(2017年8期)2017-11-24 03:21:13
现代检验医学杂志(2016年1期)2016-11-12 13:20:00
兽医导刊(2016年12期)2016-05-17 03:51:35
兽医导刊(2016年12期)2016-05-17 03:51:32
