
Ⅰ型鸭肝炎病毒RNA聚合酶基因的克隆及原核表达
2011-06-08 07:02付玉志张伟伟陈宗艳李传峰杨宗伟刘光清
中国动物传染病学报 2011年2期
王 超,付玉志,张伟伟,陈宗艳,李传峰,杨宗伟,吴 润,刘光清
(1.中国农业科学院上海兽医研究所,上海200241;2.甘肃农业大学动物医学院,兰州730070;3.湖南农业大学动物科技学院,长沙410128;4.浙江师范大学化学与生命科学学院,金华 321004)
Ⅰ型鸭病毒性肝炎属于一种急性、高度致死性的病毒性传染病[1]。该病最早由美国学者Levine报道并分离病原[2],但有关鸭肝炎病毒(Duck hepatitis virus,DHV)的分子生物学信息一直十分欠缺。台湾和韩国学者分别报道,台湾和韩国均存在与DHV-I无交叉中和反应的新型DHV,它们具有典型的DHV-I 基因组结构,与DHV-I属于同一个病毒属的成员[3]。根据衣壳蛋白编码区的序列同源性和进化分析的结果, 可将DHV区分为3个基因型,即A型、B型和C型。血清Ⅰ型、“台湾新型”和“韩国新型”分别对应于基因A型、B型和C型,表明依据衣壳蛋白序列区分的基因型与基于中和试验区分的血清型具有对应关系[4]。
DHV-Ⅰ属于小核糖核酸病毒科,其核衣壳为二十面体对称结构,无囊膜,核心部分为节段的单链正链RNA[5],长约7.7 kb。 依赖于RNA的RNA多聚酶(RNA-dependent RNA Polymerase,RdRp)基因位于其基因组的6015~7373位,RdRp全基因由1359个核苷酸组成,编码453个氨基酸。DHV-Ⅰ在全球出现和流行了半个多世纪,但其分子生物学研究起步很晚。目前,DHV的研究主要集中在DHV不同分离株基因组分析上,分子诊断技术也得到较快的发展,但是,关于上述病原的蛋白功能、病毒与宿主之间的相互作用、分子致病机理和新型疫苗的研发等方面研究非常少,对病毒的复制与调控机理了解的也非常有限。本研究采用原核表达载体对DHV-Ⅰ的RdRp进行了表达和鉴定,并基于RdRp基因的序列比较和分析了不同株DHV的同源性和遗传进化关系,给进一步研究DHV-Ⅰ的复制等方面奠定了基础。
1 材料与方法
1.1 病毒株 DHV-Ⅰ ZJ-Ⅴ株(GenBank登录号:EF382778)由中国农业科学院上海兽医研究所禽传染病研究室禽病毒病课题组保存;质粒pET32a(+)、工程菌JM109、BL21(DE3)感受态细胞均由本实验室保存。
1.2 酶和相关试剂 2xTaq PCR MasterMix 购自TIANGEN公司;T4连接酶、限制性内切酶KpnⅠ、XhoⅠ均购自TaKaRa公司;辣根过氧化物酶标记山羊抗兔二抗购自北京中杉金桥公司;Ⅰ型鸭病毒性肝炎阳性血清由本实验室制备。
1.3 引物设计与合成 根据GenBank中发表的DHV-I ZJ-V株的全基因序列,设计了针对RdRp基因的引物,预期扩增产物为1359 bp,在上游引物中加入KpnⅠ酶切位点,在下游引物中加入XhoⅠ酶切位点,引物由Invitrogen公司合成(下划线部分为酶切位点)。DH RdRp-F: 5'-ACTGGGTTAACCCCKpnIGGGAAAGTAGT AAGCAAG-3'DH RdRp-R: 5'-ACGCTTCCGGAAGGXhoITTAGATCATCA TGCAAGC-3'
1.4 RdRp基因的克隆及其诱导表达1.4.1 目的基因的扩增 用ZJ-Ⅴ株病毒的cDNA为模板按如下程序进行扩增:95℃ 预变性5 min;95℃变性 1 min,55℃退火 1 min,72℃延伸 1 min,共进行30个循环;然后72℃再延伸10 min。扩增结束后用1%琼脂糖凝胶电泳观察结果。
1.4.2 重组质粒的构建及鉴定 PCR产物纯化回收后进行双酶切,胶回收目的片段后,与经过相同的限制性内切酶酶切后的载体pET32a按适当比例用T4连接酶进行连接。连接产物转化JM109感受态,双酶切初步鉴定后送上海基因生物公司测序,验证目的基因的插入方向及阅读框架的正确性。
1.4.3 重组质粒的诱导表达及表达产物的粗纯
经鉴定正确的阳性质粒转化BL21(DE3)感受态,挑取含重组质粒的菌体单斑至4 mL LB(含Amp100μg/mL)中37℃过夜培养。按1:100比例稀释过夜菌,将2 mL菌加入到含200 mL LB培养基的1000 mL锥形瓶中, 37℃震荡培养3 h至OD600≈0.7。取部分液体作为未诱导的对照组,其余部分加入IPTG诱导剂至终浓度1mmol/L作为实验组,两组继续37℃震荡培养3.5 h。分别取菌体2 mL,10 000×g离心2 min,收获沉淀。剩余全部菌液以8000×g离心8 min收集细菌沉淀后,用包涵体粗纯法将表达产物纯化。
1.4.4 重组蛋白的检测 分别取未诱导、诱导、纯化3个样品,加入100μL 2×SDS上样Buffer水浴煮沸10 min,进行SDS-PAGE电泳分析。然后再将诱导蛋白和未诱导蛋白经过SDS-PAGE电泳后,将目的蛋白条带转移到PVDF膜上,用5%脱脂乳封闭,以鼠抗His抗体作为一抗,辣根过氧化物酶(HRP)标记的羊抗鼠IgG二抗与之反应,最后用DAB显色。
1.5RdRp基因序列分析 应用DNAStar软件,对DHV-Ⅰ ZJ-Ⅴ株RdRp基因序列与GenBank上已发表的不同国家和地区的DHV分离株的RdRp基因序列进行核苷酸序列和氨基酸序列的同源性比较,对包括ZJ-Ⅴ株病毒在内的15株不同DHV进行遗传进化分析,并基于RdRp基因序列建立进化树。
2 结果
2.1 重组质粒的构建 用ZJ-Ⅴ株病毒的cDNA为模板扩增得到与预期大小一致的1359bp的特异性RdRp目的条带(图1)。构建的重组质粒pETRdRp经KpnⅠ、XhoⅠ双酶切鉴定,切下约1359 bp的目的条带和约5900 bp质粒载体带。
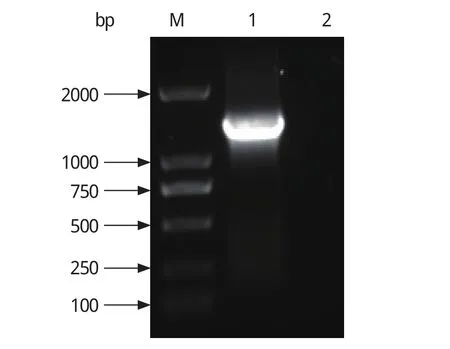
图1 DHV-RdRp PCR扩增Fig. 1 PCR product of DHV-RdRp gene
2.2 表达产物及其免疫原性的检测 重组质粒pET-RdRp以终浓度为1mmol/L的IPTG在37℃进行诱导表达3.5 h后,取诱导菌液与未诱导菌液和粗纯后的蛋白经裂解后用SDS-PAGE电泳检测蛋白表达情况,表达产物分子质量约为65 kDa,与预期结果相符合(图2)。以兔抗DHV-I血清作为一抗,辣根过氧化物酶(HRP)标记的山羊抗兔IgG二抗与之反应,结果出现目的条带(图3)。

图2 RdRp蛋白表达产物SDS-PAGEFig. 2 SDS-PAGE of RdRp recobinant protein

图3 重组蛋白Western blot检测结果Fig. 3 Western blot detection of the recombinant protein
2.3RdRp基因核苷酸序列比较及其推导的氨基酸序列分析 运用DNAStar软件分析DHV-I ZJ-V株RdRp基因序列与GenBank上发表的不同DHV分离株的基因序列,进行核苷酸序列与氨基酸序列的相似性分析。结果表明:ZJ-Ⅴ株与其他DHV毒株基因核苷酸序列的相似性为76.2%~97.1%,与DHV-I R85952株(DQ226541)的相似性最高,为97.1%;氨基酸序列的相似性为87.4%~98.8%,与DHV-I DRL-62株(DQ219396)的相似性最高。表明ZJ-V分离株与其他地方不同的DHV分离株之间,病毒基因具有较高的同源性(表1)。
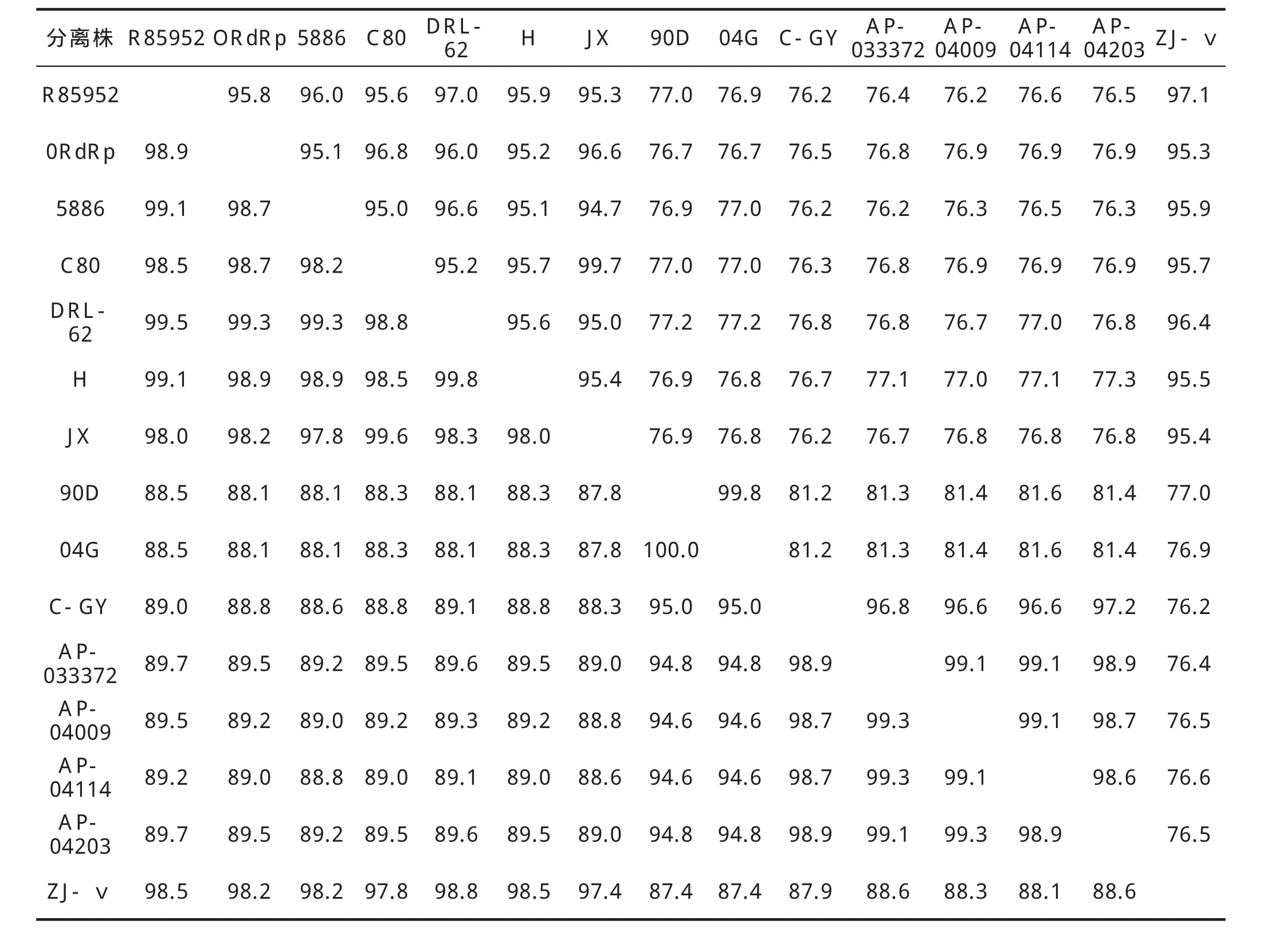
表1 DHV分离株RdRp基因核苷酸(右上)和氨基酸(左下)序列相似性比较Table 1 Pair-wise comparisons between the nucleotide (upper right)and amino acid (lower left)sequences of the RdRp gene of DHV isolates
2.4 遗传进化分析 基于RdRp基因序列构建进化树(图4),从图中可以看出,DHV-Ⅰ ZJ-Ⅴ株与DHV-Ⅰ R85952株(DQ226541)处于同一较小的分枝,亲缘关系最近。而与台湾新型DHV O4G 株 (EF067923)、DHV 90D 株 (EF067924)和韩国新型DHV AP-03337株(DQ256132)、DHV AP-04114株(DQ812093)、DHV AP-04203株(DQ256134)、DHV AP-04009株(DQ256133)处于不同的分支,亲缘关系都较远。
3 讨论
RdRp蛋白为依赖RNA的RNA聚合酶,在病毒RNA复制时主导转录及复制反应。因此,RdRp功能的保守性往往限制了RdRp编码序列的变异性,在氨基酸水平更是如此[6]。此外,RdRp可能具有核酸酶活性和RNA解旋酶活性[7]。 Plotch等[8]认为RdRp催化病毒RNA的复制作用概括起来包括:VPg的尿苷化、RNA聚合酶、末端腺苷转移酶以及与3'UTR和3AB形成RNP复合物。Poly(A)尾也是小RNA病毒复制所必需的,其长度为35~100个A不等,一般认为其长度与RNA的侵染性和复制效率有关[9]。从GenBank公布的基因组序列中,ZJ-V株的序列信息中含有22个A,但估计其实际Poly(A)尾巴应该更长,它可能是RdRp识别的信号,参与病毒基因组的复制[10]。因此,本文对于开展RdRp蛋白的研究且对于如何阐述小RNA病毒的复制及致病机理具有深刻的理论意义。

图4 基于RdRp基因序列建立的进化树Fig.4 Phylogenetic relationships based on the RdRp gene sequence
本实验选用pET32a(+)载体成功表达了带有His标签的RdRp融合蛋白,原核表达产物大约为65 kDa。在免疫印迹试验检测中,出现了特异性目的条带,证明本实验表达的RdRp蛋白具有良好的反应原性。RdRp蛋白作为一种复制酶,在病毒的生命周期发挥着重要作用,深入研究该蛋白的生物学功能对于揭示DHV-I的复制调控机理等无疑将具有重要意义。在研究DHV-I的复制与调控机理过程中,我们需要表达并纯化RdRp蛋白以及相应的抗体,以便研究RdRp与其他病毒蛋白之间以及与宿主细胞因子之间的相互作用,此外还要鉴定RdRp与病毒基因组RNA之间的相互作用,最终达到阐明DHV-I复制的分子机理以及调控过程等。因此,本文的研究内容对于我们下一步的研究提供了扎实的物质基础。另外,保证体外表达的RdRp具有蛋白酶活性对于下一步研究十分重要,我们将对此展开相关研究,本文的目的是获得正确表达的RdRp并制备相应的抗体,为将来研究RdRp的复制酶活性提供良好的物质材料。
应用软件DNAStar MegAlign程序包的Jotun Hein Method分析表明,DHV-I ZJ-V株(EF382778)与DHV-I R85952株 (DQ226541)和 DHV-Ⅰ DRL-62(DQ219396)的RdRp核苷酸同源性较高,分别为97.1%和96.4%,氨基酸的同源性分别为98.5%和98.8%。遗传进化分析,基于RdRp基因序列建立进化树,结果表明DHV-I ZJ-V株(EF382778)与DHV-I R85952株(DQ226541)处于同一分支,其亲缘关系最近。而与2株台湾新型和4株韩国新型病毒的核苷酸序列的相似性在76.4%~77.0%,氨基酸序列的相似性在87.4%~88.6%,进化树分析结果表明ZJ-Ⅴ株与这几株病毒处于不同的分支,亲缘关系较远。因此,本文在对DHV-I ZJ-V株RdRp基因序列克隆的基础上,分析了它与国内外不同基因型之间的亲缘关系,旨在从分子角度探讨RdRp基因的变异程度与不同基因型DHV-I遗传距离之间的相关性,进一步对DHV-I ZJ-V株的基因型进行鉴定,为将来研究不同基因型DHV-I复制机理的特异性做准备。
[1]韦平, 秦爱建. 重要动物病毒分子生物学[M]. 北京: 科学出版社, 2008, 219.
[2]Levine P P, Hofstad M S. Duck disease investigation[M].Annual Repport New York State Vet College, Ithaca, 1945,55-56.
[3]Ding C Y, Zhang D B. Molecular analysis of duck hepatitis virus 1 [J]. Virology, 2007, 361(1): 9-17.
[4]张大丙. 鸭肝炎病毒的分子生物学研究现状[A]//中国畜牧兽医学会禽病学分会第十四次学术研讨会论文集[C].2008.
[5]殷震, 刘景华. 动物病毒学[M]. 2版. 北京: 科学出版社,1997.
[6]Yamashita T, Sakae K, Tsuzuki H, et al. Complete nucleotide sequence and genetic organization of Aichi virus, a distinct member of the Picornaviridae associated with acute gastroenteritis in humans [J]. J Virol, 1998, 72(10): 8408-8412.
[7]Martinez-Salas E, Regalado M P, Domingo E. Identification of anessential region for internal initiation of translation in the aphthovirus internal ribosome entry site and implications for viral evolution[J]. J Virol, 1996,70(2):992-998.
[8]Plotch S J, Palant O. Poliovirus protein 3AB form a complex with and stimulates the activity of the viral RNA polymerase,RdRppol[J]. J Virol, 1995, 69(11): 7169-7179.
[9]Silvestri L S, Parilla J M, Morasco B J, et al. Relationship between poliovirus negative-strand RNA synthesis and the length of the 3'poly(A)tail[J]. Virology, 2006, 345(2):509-519.
[10]Kim M C, Kwon Y K, Joh S J, et al. Molecular analisis of duck hepatitis virus type Ⅰreveals a novel lineage close to the genus Parechovirus in the family Picornaviridae[J].J Gen Virol, 2006, 87(Pt11): 3307-3316.
猜你喜欢
数学物理学报(2022年5期)2022-10-09
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
河北画报(2020年8期)2020-10-27
中国饲料(2019年19期)2019-03-25
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
