
靶向VP1和VP2双拷贝shRNA重组表达质粒抑制传染性法氏囊病病毒复制的研究
2011-05-21 09:41欧阳伟王永山马金荣张海彬
中国预防兽医学报 2011年10期
欧阳伟,王永山*,马金荣,,张海彬
(1.江苏省农业科学院兽医研究所/国家兽用生物制品工程技术研究中心,江苏 南京 210014;2.南京农业大学动物医学院,江苏 南京 210095)
传染性法氏囊病(Infectious bursal disease,IBD)是由传染性法氏囊病病毒(IBDV)引起的一种危害雏鸡的急性高度接触性传染病。IBDV感染不仅可导致患病鸡死亡,而且引起鸡的免疫抑制,造成疫苗免疫失败[1-7]。
IBDV属于双RNA病毒科双RNA病毒属,基因组包括大(A)小(B)两个片段。VP1是病毒的非结构蛋白,由B片段编码的一种依赖RNA的RNA聚合酶(RdRp),与病毒RNA的复制和毒力有关[8-10]。VP2是病毒的主要结构蛋白之一,由A片段编码的多聚蛋白加工而成,位于病毒粒子的外表面,占总结构蛋白量的51%,属于病毒的主要保护性抗原,与病毒中和抗体的诱导、抗原和毒力的变异以及细胞凋亡的诱导等有关[11]。
RNA干扰(RNA interference,RNAi)能够使mRNA发生降解而导致基因表达沉默[12-13]。RNAi被认为是机体阻止外来病毒感染、保护细胞免受病毒入侵的一种细胞自我保护机制[14]。
本实验室前期的研究采用体外转录的方法分别制备了3个靶向VP1和3个靶向VP2的小干扰RNA(Short interfering RNA,siRNA),在鸡胚成纤维细胞(CEF)中的功能分析表明,3个VP1-siRNA和3个VP2-siRNA均能明显抑制IBDV复制,其中VP1-siRNA2550和VP2-siRNA392的抑制效果最为显著[15]。在此基础上,本实验选择这两个高效siRNA,构建了由chU6启动的靶向IBDV VP1和VP2基因的双拷贝shRNAs重组表达质粒,鉴定其在CEF及鸡胚中对IBDV复制的抑制效果,为新型抗IBDV生物制剂与疫苗设计探索新的技术途径。
1 材料和方法
1.1 病毒株及主要试剂 IBDV B87 CEF适应病毒株和鸡胚适应病毒株为本实验室保存;9日龄~10日龄SPF鸡胚购自南京天邦生物科技有限公司;pSilencer 2.1-U6 neo购自Ambion公司;SYBR PrimeScript RT-PCR试剂盒、MiniBEST质粒纯化试剂盒、DNA凝胶回收试剂盒购自TaKaRa公司;Lipofectamine 2000转染试剂购自Invitrogen公司。
1.2 鸡U6启动子(chU6)的克隆 根据已发表的chU6基因序列(DQ531569)设计引物,chU6(F):5'-CGGAATTCACGCGTCGACGCGGCCGCGACAAC ACAAGCATCGAG-3'; chU6(R): 5'-CCCAAGCTT CCGCTCGAGCGGGATCCTAGTATATGTGCTGCCG AAGCGAGCACGGAC-3'。上游引物引入 EcoRⅠ、SalⅠ位点,下游引物引入HindⅢ、XhoⅠ、BamHⅠ位点,引物由Invitrogen公司合成。
取60日龄的来航鸡肝脏组织,按TIANDZ公司的柱式动物DNAout说明书提取基因组DNA,通过PCR扩增chU6启动子序列,克隆于pMD18-T载体中,EcoRⅠ和HindⅢ双酶切分析,获得含chU6启动子的重组质粒pMD-chU6并进行测序分析。
1.3 靶向VP1和VP2基因的shRNA分子设计根据本实验室前期研究结果[15]和shRNA设计原则[16],选择IBDV VP1基因(DQ403249)的2550序列和VP2基因(AF508177)的392序列为靶标,设计编码shRNA的正义和反义链DNA序列,退火形成双链DNA,插入RNAi表达载体pSilencer 2.1-U6 neo,同步设立无shRNA的阴性对照。序列设计见表1和图1。

表1 编码shRNA的DNA序列Table 1 DNA sequence encoding shRNA
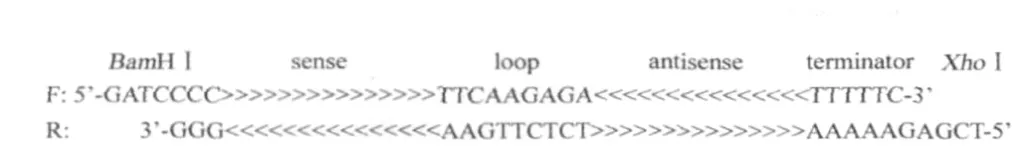
图1 shRNA的设计Fig.1 Design of the shRNA
1.4 双向chU6启动子双拷贝shRNA重组表达质粒的构建 将合成的编码VP1-shRNA2550和VP2-shRNA392的正向和反向互补单链DNA序列分别退火(图1),得到VP1-shRNA2550和VP2-shRNA392双链DNA。将其分别插入pMD-chU6的BamHⅠ与XhoⅠ位点,分别获得pMD-chU6-VP1shRNA2550和pMD-chU6-VP2shRNA392重组质粒。用XhoⅠ单酶切pMD-chU6-VP2shRNA392,SalⅠ与XhoⅠ双酶切pMD-chU6-VP1shRNA2550,回收pMD-chU6-VP2shRNA392片段和chU6-VP1shRNA255片段,连接,转化,BamHⅠ单酶切分析,获得pMD-2chU6-VP1VP2shRNA。将pMD-2chU6-VP1VP2shRNA和表达载体pSilencer 2.1-U6 neo分别用EcoRⅠ和HindⅢ双酶切,回收2chU6-VP1VP2shRNA片段与pSilencer 2.1-U6 neo片段,连接,转化,EcoRⅠ和HindⅢ双酶切分析,获得双chU6启动子双shRNA重组表达质粒pSilencer2.1-2chU6-VP1VP2shRNA(中国发明专利申请号:201110127319.7),简称为:pchU6-shRNA12(图2)。同步构建不含shRNA的对照质粒pchU6-CON。

图2 双chU6启动子双shRNA表达质粒的分子图谱Fig.2 Schematic diagram of the recombinant plasmid pchU6-shRNA12
1.5 pchU6-shRNA12在CEF中体外抑制IBDV试验 试验在6孔板上进行,按Lipofectamine2000说明书操作方法将 pchU6-shRNA12(4μg/孔)转染CEF,24 h后接种103TCID50的IBDV(B87株),继续培养72 h,观察细胞病变(CPE),收集CEF培养物,冻融3次,测定病毒滴度,用Reed-Muench法计算病毒的TCID50。实验同步设立pchU6-CON转染CEF、病毒感染和正常CEF对照组。
1.6 pchU6-shRNA12在鸡胚中抑制IBDV试验将 pchU6-shRNA12(8 μg/ 胚 )与 IBDV(103TCID50/胚)混合,一起接种于10日龄SPF鸡胚尿囊腔中,同步设立pchU6-CON以及单独接种IBDV对照组。观察鸡胚生长和发育状况,96 h后,收集鸡胚组织和尿囊液,接种10日龄SPF鸡胚,测定病毒滴度,用Reed-Muench法计算病毒的ELD50。
根据IBDV VP1基因、VP2基因和鸡β-actin基因(作为内参对照)序列设计引物(表2),引物由上海Invitrogen公司合成。

表2 荧光定量RT-PCR引物Table 2 Primers for real time quantitative RT-PCR
用TRIzol试剂方法提取鸡胚组织和尿囊液混合物的总RNA,按照SYBR PrimeScript RT-PCR Kit使用说明进行反转录合成cDNA,将cDNA用EASY Dilution按10倍梯度稀释,再将各稀释度的cDNA作为模板进行SYBR GreenⅠreal-time PCR扩增,以β-actin基因作为内参照。实时荧光定量PCR仪为LightCycler 480Ⅱ(Roche公司),扩增结果采用Roche LightCycler 480 SW1.5.0软件做相对定量分析,计算抑制效率。
2 结 果
2.1 chU6启动子的克隆 以鸡的基因组DNA为模板,PCR扩增获得约为600 bp的chU6启动子,将其克隆于pMD18-T中,经EcoRⅠ和HindⅢ双酶切分析,可见2条长度分别约600 bp和2700 bp的DNA片段,与chU6基因片段(593 bp)和pMD18-T载体大小理论值相符合(图3)。

图3 pMD-chU6的酶切图谱Fig.3 Identification of pMD-chU6
2.2 chU6的生物信息学分析 将测定的chU6启动子序列在GenBank中进行BLAST分析,chU6启动子173-566位核苷酸序列(GenBank中登录号:JN127375)与已发表的鸡U6(DQ531569)启动子序列同源性为96%,而与人源的U6启动子(人染色体15∶NCBI35∶15∶65919386∶65919660∶-1)同源性为 6.2%,与鼠源的 U6启动子(鼠染色体 9∶NCBIM33∶9∶63370029∶63370291∶1)同源性为 9.9%。用 TFSEARCH分子生物学软件对克隆的chU6启动子转录因子结合位点分析,有4个潜在的Oct-1结合位点和其它转录因子结合位点,含有RNA聚合酶Ⅲ的3个上游元件:OCT(ATTTGCAT)、PSE(GTCACTGTGTTCT AAAAGAACTTG)、TATA(TTAAATA)。
2.3 双向chU6启动子双拷贝shRNA重组表达质粒的构建 将2chU6-VP1VP2shRNA基因片段和pSilencer 2.1-U6 neo载体片段连接,获得重组表达质粒pchU6-shRNA12,经EcoRⅠ和HindⅢ双酶切分析,可见2条长度分别约1300 bp和4200 bp的DNA片段,与pSilencer 2.1-U6 neo和2chU6-VP1VP2shRNA基因片段(1324 bp)的理论值相符合(图4)。
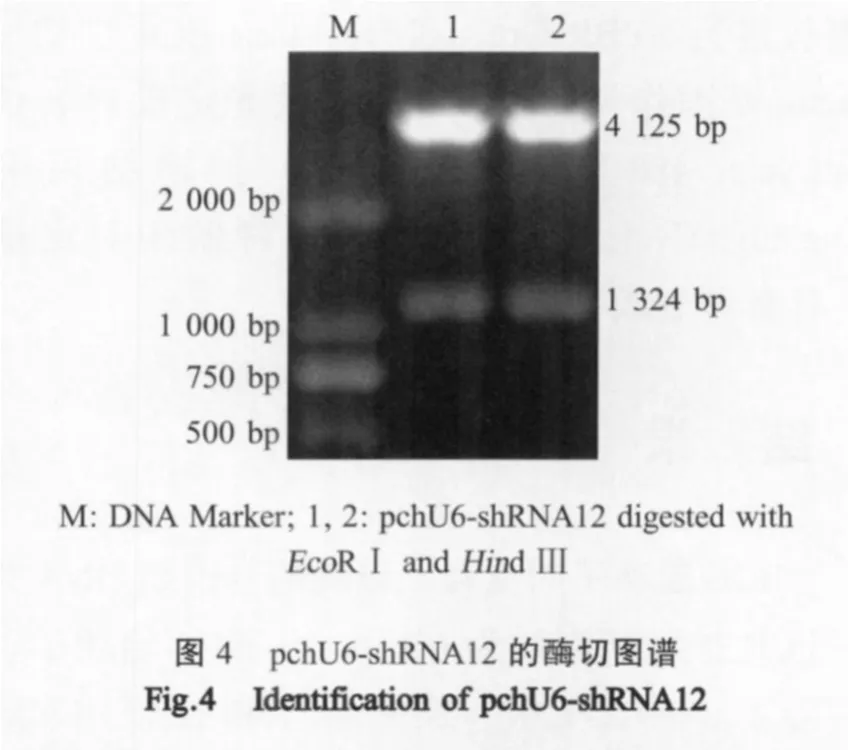
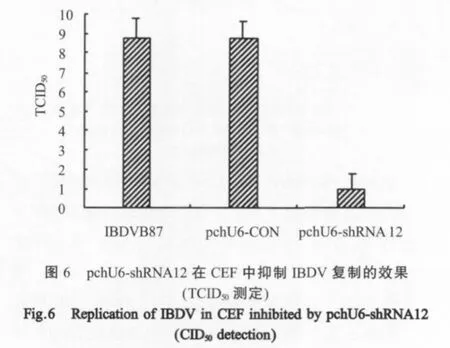
2.4 pchU6-shRNA12在CEF中的抗病毒活性检测将 pchU6-shRNA12转染的CEF,接种 IBDV,72 h后,未见CPE,细胞形态呈正常的梭状。而pchU6-CON转染组和IBDV感染CEF两个对照组则呈现典型的CPE(图5)。病毒滴度测定结果表明,pchU6-shRNA12转染组的病毒滴度小于101TCID50/0.1 mL,而pchU6-CON转染组和IBDV感染组两个对照组的病毒滴度均达到108.75TCID50/0.1 mL(图6)。
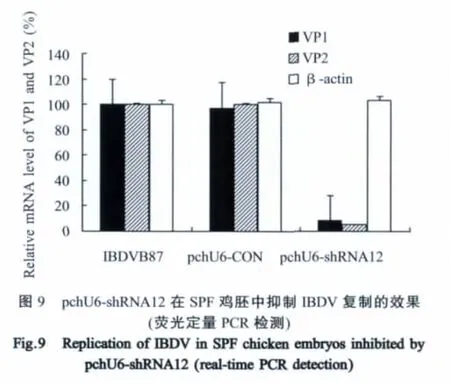
2.5 pchU6-shRNA12在鸡胚中的抗病毒活性检测将pchU6-shRNA12与IBDV混合接种的SPF鸡胚,96 h后,鸡胚继续存活,检查胚体发育正常;而同步接种pchU6-CON以及单独接种IBDV的两个对照组的鸡胚则全部死亡,胚体发育阻滞、出血(图7)。采用10日龄SPF鸡胚测定鸡胚组织和尿囊液混合物的ELD50,pchU6-shRNA12组病毒滴度小于101ELD50/0.1 mL,而pchU6-CON组和IBDV感染组两个对照组的病毒滴度均达到107.00ELD50/0.1 mL(图8)。用实时荧光定量RT-PCR对VP1基因和VP2基因的拷贝数进行定量检测,pchU6-shRNA12转染组VP1基因比IBDV对照组降低了92%,VP2基因比IBDV对照组降低了95%(图9)。

3 讨 论


常用的shRNA表达载体中RNA聚合酶Ⅲ启动子(polⅢ)主要为人源或鼠源的U6启动子或人H1启动子,它们在禽源细胞中的转录活性说法不一。Katahira等的研究结果显示,小鼠U6启动子能够驱动shRNA在鸡胚中表达[17]。Dai等也证明,人H1和U6启动子能驱动GFP基因特异性shRNA在鸡胚中表达[18]。而Das等则获得与上述相反的结果[19]。鉴于此,本研究为了构建靶向IBDV的shRNA高效表达质粒,将shRNA表达载体pSicencer2.1-U6 neo中的人源U6启动子替换成chU6启动子。采用PCR方法从鸡的基因组DNA中扩增克隆的chU6启动子,与鸡的同源性较高,而与人、鼠的同源性较低,该chU6启动子转录因子结合部位含有RNA polⅢ的3个上游元件,通常认为,在RNA polⅢ的上游启动子中,只要靠近起点存在TATA元件,就能起始转录,PSE和OCT元件的存在可提高转录效率[20]。
本研究在前期研究基础上,根据RNAi效率高的VP1-siRNA2550和VP2-siRNA392两个序列,设计VP1-shRNA2550与VP2-shRNA392,选用chU6启动子,构建一个同时靶向IBDV VP1和VP2基因的双向chU6启动子双拷贝shRNA表达质粒pchU6-shRNA12,其中 VP1-shRNA2550和 VP2-shRNA392分别由独立的chU6驱动表达。该pchU6-shRNA12有3个特点:用chU6启动子替代人源U6启动子,以提高外源shRNA在鸡细胞中的转录效率;VP1-shRNA2550和VP2-shRNA392双shRNA可以同时抑制IBDV的两个功能基因VP1和VP2转录表达,双倍提高RNAi效果;双启动子独立驱动两个shRNA高效转录,不受转录位置先后的影响。
为验证pchU6-shRNA12在活体外抑制IBDV复制的功能,用pchU6-shRNA12在CEF进行了IBDV复制的抑制实验,干扰组未出现CPE,细胞培养物中的病毒滴度小于101TCID50/0.1 mL;而对照组均出现明显的CPE,病毒滴度均达到108.75TCID50/0.1 mL。Gao等根据IBDV VP1基因的2571序列构建了shRNA表达载体,在Vero细胞上进行了对IBDV复制的抑制实验,抑制率为87.4%[21]。与之相比,pchU6-shRNA12具有更高的抑制效果,分析原因可能是由于pchU6-shRNA12转录产物能同时靶向降解VP1和VP2基因,因此,对IBDV的干扰效率更高。本实验用转染试剂Lipofectamine2000递呈表达质粒,其干扰效果与不用转染试剂相当(实验数据未提供)。
为进一步验证pchU6-shRNA12在活体内抑制IBDV的复制能力,用鸡胚进行了鸡胚感染抑制试验。pchU6-shRNA12转染组鸡胚发育正常,鸡胚组织和尿囊液混合物病毒滴度显著低于两个对照组,表明pchU6-shRNA12在鸡胚内也能抑制IBDV的复制;用荧光定量RT-PCR分别检测VP1和VP2的表达量,pchU6-shRNA12转染组比单纯接种IBDV对照组分别降低了92%和95%,进一步表明IBDV复制的抑制是由于VP1和VP2基因被特异性降解所致。
本研究结果表明:chU6启动子在pchU6-shRNA12中能高效驱动双拷贝shRNA的转录,转录的siRNA在体外(CEF)以及体内(鸡胚)均能抑制IBDV的复制,为IBDV新型治疗预防生物制品的研发探索出了一条新的技术途径。目前正在进行动物实验。
[1]Wang Yong-shan,Wang Zong-can,Tang Yu-de,et al.Comparison of four infectious bursal disease virus isolated from different bird species[J].Arch Virol,2007,152(10):1787-1797.
[2]王永山,欧阳伟,潘群兴,等.近期引起免疫失败的传染性法氏囊病病毒VP2基因的分子特征[J].中国兽医科学,2008,38(12):919-925.
[3]曹冰玉,王永山,范红结,等.引起免疫失败的传染性腔上囊病病毒VP2基因在大肠杆菌中的表达[J].中国兽医科学,2010,40(02):130-134.
[4]欧阳伟,王永山,周宇,等.传染性法氏囊病病毒AH1株vp2基因在昆虫细胞中的表达与应用[J].生物工程学报,2010,26(5):595-603.
[5]Wang Yongs-han,Fan Hong-jie,Li Yin,et al.Development of a multi-mimotope peptide as a vaccine immunogen for infectious bursal disease virus[J].Vaccine,2007,25(22):4447-4455.
[6]刘小娟,王永山,欧阳伟,等.传染性法氏囊病病毒抗原表位与乙型肝炎病毒核心抗原嵌合基因的构建及其表达产物分析[J].中国预防兽医学报,2010,32(11):879-883.
[7]周宇,王永山,范红结,等.IBDV抗原表位与VP2组合基因的分子构建及其在昆虫细胞中的表达[J].中国兽医学报,2010,30(10):1301-1306.
[8]Mirriam G J,Tacken L,Ben P H,et al.Infectious bursal disease virus capsid protein VP3 interacts both with VP1,the RNA-dependent RNA polymerase,and with viral double-stranded RNA[J].J Virol,2002,76(22):11301-11311.
[9]Liu Mei-hong,Vakharia V N.VP1 protein of infectious bursal disease virus modulates the virulencein vivo[J].Virology,2004,300(1):62-73.
[10]Boot H J,Hoekman A J,Gielkens A L.The enhanced virulence of very virulent infectious bursal disease virus is partly determined by its B-segment[J].Arch Virol,2005,150(1):137-144.
[11]Armando F A,Siomara M,Jose F R.The major antigenic protein of infectious bursal disease virus,VP2,is an apoptotic inducer[J].J Virol,1997,71(10):8014-8018.
[12]Fire A,Xu Sai-long,Montgomery M K,et al.Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans[J].Nature,1998,391(6669):806-811.
[13]Zamore P D,Tuschl T,Sharp P A,et a1.RNAi:double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals[J].Cell,2000,101(1):25-33.
[14]Meister G,Tuschl T.Mechanisms of gene silencing by double-stranded RNA[J].Nature,2004,431(7006):343-349.
[15]欧阳伟,王永山,刘小娟,等.靶向传染性法氏囊病病毒VP1基因siRNA抑制病毒复制实验[J].中国兽医学报,2011,30(10):印刷中.
[16]Sui Guang-chao,Soohoo C,Affar el B,et a1.A DNA vector-based RNAi technology to suppress gene expression in mammalian cells[J].PNAS,2002,99(8):5515-5520.
[17]Katahira T,Nakamura H.Gene silencing in chick embryos with a vector-based small interfering RNA system[J].Dev Growth Differ,2003,45(4):361-367.
[18]Dai Fang-ping,Yusuf F,Farjah G H,et al.RNAi-induced targeted silencing of developmental control genes during chicken embryogenesis[J].Dev Biol,2005,285(1):80-90.
[19]Das R M,van Hateren N J,Howell G R,et al.A robust system for RNA interference in the chicken using a modified microRNA operon[J].Dev Biol,2006,294(2):554-563.
[20]王镜岩,朱圣庚,徐长法,主编.生物化学[M],3版,下册,北京:高等教育出版社,2005,464.
[21]Gao Yu-long,Liu Wei.Effective inhibition of infectious bursal disease virus replicationin vitroby DNA vector-based RNA interference[J].Antiviral Res,2008,79:87-94.
猜你喜欢
吉林畜牧兽医(2022年2期)2022-11-16
昆明医科大学学报(2022年2期)2022-03-29
中国农业大学学报(2022年2期)2022-01-05
华侨大学学报(自然科学版)(2021年4期)2021-07-30
江西农业学报(2021年4期)2021-04-20
水生生物学报(2021年1期)2021-02-04
三农资讯半月报(2020年11期)2020-06-21
中国畜牧兽医文摘(2018年6期)2018-07-28
中国全科医学(2016年14期)2016-06-16
中国当代医药(2015年9期)2015-03-01
