
北京山区栓皮栎水源涵养林空间结构特征研究
2011-05-07 10:59赵阳余新晓宋思铭杨芝歌王贺年邓文平
水土保持研究 2011年3期
赵阳,余新晓,宋思铭,张 艺,杨芝歌,王贺年,邓文平
(水土保持与荒漠化防治教育部重点实验室,北京林业大学水土保持学院,北京100083)
林分空间结构与树木在林内的空间位置密切相关[1]。一方面,它不仅体现了树木在林地上的分布格局及其属性在空间上的排列方式,另一方面,它还决定了树木的空间生态位以及树木之间的竞争优势,在很大程度上影响着林分生长、发育和稳定性[2]。为此,林分空间结构特征研究对森林经营技术研究、森林结构与功能研究乃至森林健康评价都有着重要意义。森林结构研究方法主要有三种,即:传统森林经理学方法、经典植被生态学方法以及现代森林生态和森林经理学方法[3]。与前两种研究方法不同,现代森林生态和森林经理学方法是以相邻木关系为基础,在充分考虑点的空间位置前提之下,通过获取种群数量分布的空间信息,来分析林木在空间结构单元中的空间关系,较前两种方法而言,现代森林生态学和森林经理法分析所得的林分空间结构可解析性更高。
栓皮栎(Quercus variabilis),山毛榉科,栎属植物,原产北非地中海沿岸,是我国重要的用材树种,在我国分布极为广泛。其作为北京低山地区森林组成的主要树种之一,在水源涵养、水土保持以及净化环境等方面发挥着不可替代的作用。几十年来,受立地条件等因素的制约,栓皮栎林生态系统呈现出系统稳定性差,生态服务功能低下等问题。在以往对北京山区栓皮栎水源涵养林结构的研究主要集中在群落结构、物种组成、物种多样性等方面[4-6],鲜有对栓皮栎林空间结构特征的研究。本研究将采用现代森林生态学和森林经理法,利用描述林分空间结构的混交度、大小比数等参数并结合空间点格局分析法对西山林区栓皮栎水源涵养林的空间结构特征进行分析研究,从优化森林空间结构的角度,分析和准确表达栓皮栎水源涵养林生态系统目前所存在的问题及其原因,旨在为北京地区栓皮栎水源涵养林的近自然经营提供理论模式和技术支持,进而全面提高水源涵养林的生态服务功能。
1 研究区概况
西山林区位于北京市西部山区,东经116°28′,北纬 39°34′,属于太行山余脉,平均海拔 300.4 m,最高峰克勒峪峰海拔800 m,山脚线海拔100 m,坡度15°~45°。该区属暖温带大陆性气候,冬寒夏热,春季多风,平均风速4.1 m/s,年平均气温为11.6℃,无霜期193 d,年平均日照2 662 h。年降水量630 mm,主要集中在夏季,其中6-8月的降雨量占全年降雨量的70%以上。本研究区地带性土壤为山地褐色土,大部分为淋溶褐色土,土壤发育层次不明显,含石砾较多,一般土层厚度40 cm左右。全林区内植物种类丰富,自然生长的乔木种类较少,多为20世纪50-60年代营造的人工林。主要乔木树种有:油松(Pinus tabulaef ormis)、侧柏(Platycladus orientalis)、白皮松(Pines bungeana)、华北落叶松(Larix principisrupprechti)、栓皮栎(Quercus variabilis)、刺槐(Robinia pseudoacacia)、元宝枫(Acer truncatum)、槲栎(Quercus dentata)、辽东栎(Quercus liaotungensis)、栾树(Koelreuteria paniculata)、桑(Mores alba)等;主要灌木有:黄栌(Cotinus coggygria)、荆条(Vitex negundo var.heterophylla)、胡枝子(Lespedeza bicolor)、孩儿拳头(Grewia biloba)、蚂蚱腿子(Myripnois dioica)、紫穗槐((Amorp f ruticosa)等。
2 研究方法
2.1 标准地调查
2010年8月在对西山水源涵养林区森林植被进行全面踏查的基础之上,根据林分特点,按照选取具有代表性、原始性、典型性样地的原则,采用典型选样法,在西山林区栓皮栎林内设置了栓皮栎林固定调查样地1块,样地设在山地深处,地理位置较偏僻,样地周围没有发现人为采伐痕迹,人类的轻度干扰并未对栓皮栎林的空间结构产生较大影响。样地以栓皮栎种群为优势种,样地面积为120 m×120 m。用GPS进行定位,测定样地坡度、坡向、经度、纬度、海拔等立地因子,并进行植被群落学调查。经测定:样地海拔为401 m,坡向东偏南20°,坡度18°,立地条件较好,郁闭度为0.7,林龄为40 a。采用相邻网格法,将样地分成36个20 m×20 m的样方网格,网格从左到右、从上到下依次放置。再以每个网格为调查单元,运用地面三维激光扫描技术采用“多站式”扫描方法对样地内每个样方网格进行全景扫描,通过对扫描所得的各个网格的点云图像进行坐标构建、去噪、选取、拼接处理进而得到整个样地的三维立体点云模型,在该模型基础之上,利用地面三维激光扫描仪所配备的数据处理软件采用聚点法(clustering method)可对林分内的乔木进行树高、胸径、枝下高、胸高断面积、冠幅、冠径等参数信息的量测以及树木的精确定位[7]。此外,参照整个标准地的全景立体式点云图像以及扫描同步所拍摄的样地照片可以辨别与参照树最近的4株胸径大于4 cm树木的树种、大小比数和混交度等空间结构参数信息。与采用传统方法获取测树因子人为工作量大、测算不准确、给林业生产造成极大不便相比,地面三维激光扫描技术的应用不论是在测树效率还是在测量精度上都有了较大提高[8-13]。此外,鉴于林分空间结构参数与样地大小有关,惠刚盈等研究认为,当样地面积为0.25 hm2及以上时能够较为真实地反映林木空间分布格局[14],本研究中的栓皮栎林样地面积为1.414 hm2,完全能够满足林分空间结构分析的需要。
扫描样地过程中,为了消除林分边缘树木对林分结构的影响,样地采用8邻域样地法进行边缘校正。所谓8邻域样地就是分别在原样地的上、下、左、右、左上、左下、右上、右下 8个方向复制原样地,形成由9个相同样地组成的大样地,再计算原样地的林分混交度,以消除边缘影响[15]。
2.2 林分空间结构分析
以样地扫描数据为基础,利用空间结构分析软件Winkelmass 1.0对固定样地的林分结构参数进行综合分析,分别计算了栓皮栎林林分内的树种混交度、大小比数等林分结构参数。林分空间结构参数的计算如下[16]:
(1)树种混交度(Mi)是指参照树i的n株最近相邻木中与参照树不属于同种的个体所占的比例,用公式表示为

式中:n——参照树的总株数,当参照树i与第j株相邻木非同种时,vij=1,否则vij=0。当考虑参照树周围的4株相邻木时,Mi的取值有5种:0.00,0.25,0.50,0.75,1.00。当 Mi=0.00时,参照树 i周围 4株最近相邻木与参照树均属于同种;Mi=0.25表示参照树i周围4株最近相邻木有1株与参照树不属于同种;Mi=0.50表示参照树i周围4株最近相邻木有2株与参照树不属于同种;Mi=0.75表示参照树i周围4株最近相邻木有3株与参照树不属于同种;Mi=1.00表示参照树i周围4株最近相邻木与参照树均不属于同种。这5种取值对应于混交度的描述为零度、弱度、中度、强度和极强度混交。
(2)林分大小比数定义为大于参照树的相邻木株数占所考查的全部最近相邻木的比例。所谓的“大小”用胸径、树高和冠幅均可表示。用公式表示为

其中,如果相邻木j比参照树i小,kij=0;否则,kij=1。当所观察树种的参照树数量为 4时,大小比数(Ui)有5种取值可能,即:0.00,0.25,0.50,0.75,1.00。Ui=0.00(相邻木均比参照树小);Ui=0.25(1株相邻木比参照树大);Ui=0.50(2株相邻木比参照树大);Ui=0.75(3株相邻木比参照树大);Ui=1.00(4株相邻木比参照树大)。对应于参照树在4个相邻木中不同的优势程度,即优势、亚优势、中庸、劣态和绝对劣态。
在本研究的相关计算中,为能明显地表达出混交度等结构参数的林学意义,选用n=4,即在参照树周围选用4株相邻木组成一个结构框架。惠刚盈等研究表明由1株参照树与其4株相邻木所组成的结构单元中,参照树与4株最近相邻木构成的结构关系有5 种 ,即零度 、弱度 、中度 、强度 、极强度,相比 n=3 或5在其类型划分上仅有4种或6种(偶个数),缺乏中间过渡类型,不符合自然现象而言,n=4时,过渡阶段更加完整,生物学意义则更加明显,空间结构信息比较完整,且这种结构单元的可释性和可操作性都比较强,较适宜于描述林分的空间结构特征[17-19]。
2.3 点格局分析
点格局分析统计学理论由B.D.Ripley首先提出[20],该方法可用于分析不同尺度的种群空间分布格局及其种间关系,因此,广泛应用至今。
该法所需数据来源于树种个体在空间中的二维坐标,树种的个体在空间中都可以看成是二维空间中的1个点,因此,1个样方中所研究的所有树种都可用一系列点来表示,即样方中所研究的植物个体构成了空间分布的点图,空间点格局分析便是在点图基础之上进行的。同样,群落内不同种群和同一种群的分布格局分析均可应用这一原理。
密度(λ)和协方差(k)是二维数集的一次和二次特征结构。对于点格局,λ是单位面积内的期望点数,k是点间距离分布的测定指标,k随着尺度的变化而变化[21]。证明该二次特征结构可以简化为1个函数方程K(r),公式为

式中:r——大于0的任何值;λ——单位面积上的点数,可以用n/A来估计;A——样地面积;n——总点数(植物个体数)。
在实际应用中,用下式估计

式中:uij——2个点i和 j之间的距离,当 uij≤r时,It(uij)=1,当 uij>r时,It(uij)=0;Wij— —以点 i为圆心;uij——半径的圆周长在面积A中的比例,其为1个点可被观察到的概率,可校正边界效应引起的误差。用K(r)和随机分布 k(r)相比构造统计量,用L(r)表示为
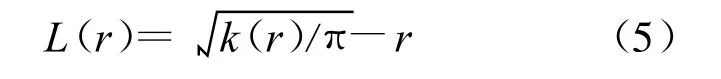
当 L(r)=0时,表示随机分布;当 L(r)>0时,为聚集分布;当L(r)<0时,为均匀分布。
用Monte-Carlo拟合检验计算上下包迹线,即置信区间。假定种群是随机分布,则用随机模型拟合一组点的坐标值,对每一个r值,计算 L(r);同样,用随机模型再拟合新一组点坐标值,分别计算不同尺度r的L(r),这一过程重复进行,直到达到事先确定的次数,L(r)的最大值和最小值分别为上下包迹线的坐标值。用r作为横坐标,上下包迹线作为纵坐标绘图,置信区间一目了然。用种群实际分布数据计算得到的不同尺度下的L(r)值。若在包迹线以内,则符合随机分布;若在包迹线以外,则显著偏离随机分布,呈集群分布。当种群表现为聚集分布时,把偏离随机置信区间最大值作为最大聚集强度指标,而聚集规模是以聚集强度为半径的圆[22]。
种间关系分析的定义和计算原理与单种格局相近,k12(r)可以用下式估计。

式中:n1,n2——种1和种2的个体数;i,j——种1和种2的个体,同样计算L12(r):
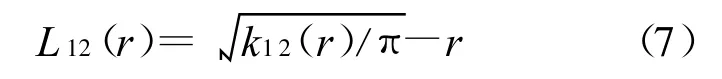
当L12(r)=0时,表示两个种在r尺度下无关联性;当L12(r)>0时,表示正关联;L12(r)<0时,表示负关联。仍然用Monte-Carlo方法检验拟合置信区间,以检验2个种之间是否有显著的关联。本文中点格局分析过程是通过生态学R软件和Excel软件完成。
3 结果与分析
3.1 栓皮栎林树种组成
栓皮栎水源涵养林林分树种组成概况见表1。从表1可以看出,在所调查的样地内,乔木层共有16个树种,林分密度约为1 415株/hm2,林分郁闭度为0.7。其中,栓皮栎种群在该林分内株数比例达67%,占有绝对优势,既是该群落的优势种,也是建群种;伴生树种主要有元宝枫、油松、槲树、构树等15个树种,伴生树种株数所占比例较小,除元宝枫外,其他伴生树种株数比例均不足10%。其中,林分内针叶树株数比例为0.5%,阔叶树为99.5%,栓皮栎种群断面积占林分总断面积的77%,元宝枫的断面积占林分总断面积的8%,其它树种的断面积占林分总断面积的比例均小于8%,说明该林分以阔叶树种为主,针叶树种为辅的栓皮栎元宝枫混交林,林分树种组成式为:8栓皮栎1元宝枫1槲树+油松+刺槐-构树-五角枫-桑树-侧柏-栾树-枸树-小叶朴-山楂-小叶椴-酸枣-山桃。从林分垂直结构看,该林分可划分为3个林层:小于5 m,5~8 m,大于8 m。其中,栓皮栎种群的平均树高处于最高层,在空间中占据优势。综合林分树种各项指标分析表明,栓皮栎水源涵养林物种多样性较丰富,整个林分内以栓皮栎种群个体分布范围最广,蓄积量最大,占据着林分的上层空间,林木生长状况与其它树种相比较好。

表1 栓皮栎林林分概况
3.2 栓皮栎林空间结构
3.2.1 树种混交度 栓皮栎林林分混交度及其分布见表2和图1。由图1可知,栓皮栎林树种混交度分布规律明显,混交度从Mi=0到Mi=1不同取值的比例大致呈减少趋势,林分平均混交度为0.32,林分以零度混交为主,零度混交所占总体比例为43%,说明该栓皮栎林林分内树种混交程度较低,林分中同种树大多聚集在一起,处于混交不良的范畴。其中,林分内主要树种栓皮栎种群以零度、弱度混交为主,两种混交方式的林木株数比例为75%,说明栓皮栎种群的混交状况不良,空间结构单元中常与3株或3株以上的同种相伴;元宝枫种群零度和弱度混交的比例为40%,混交度较低,仅次于栓皮栎种群;油松、槲树 、构树 、桑树 、栾树 、枸树 、小叶朴 、山楂 、小叶椴 、山桃的混交度以强度和极强度为主,说明这些伴生种群混交程度普遍较高,种群同种个体聚集程度较低,林木混交状况较好。总体而言,由于该林分优势种群栓皮栎株数占整个林分总株数的67%,其混交度较低进而影响到整个林分混交状况不良,整个林分林木聚集度高,伴生树种除元宝枫种群外,其他种群同种个体聚集程度较低,树种混交度较大,林木混交状况较好。

表2 栓皮栎林各树种混交度及大小比分布

图1 林分混交度分布图
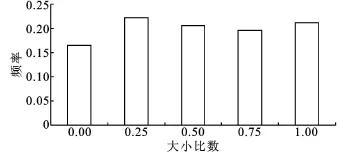
图2 林分大小比数分布图
3.2.2 林木大小分化程度 栓皮栎水源涵养林林分大小比数分布见图2。由图2可知,栓皮栎林林分大小比数分布比较均匀,优势木相比而言较少。也说明栓皮栎混交林内林木大小差异不明显,在空间结构单元内,不同等级的林木分布频率相差不大,分布较均匀,林木分化程度高。由表2可得知,栓皮栎、油松、槲树、刺槐种群在空间结构单元中以优势木、亚优势木和中庸木为主,分别占种群总株数的67%、51%、71%和57%。其中,栓皮栎种群有45%的个体在胸径上处于优势地位,超过22%的个体处于中庸状态,处于极劣势地位的株数则相对较少,加之栓皮栎种群株数在林分内所占比例大,在林分垂直结构中占据着主林层有利生态位,故栓皮栎种群在林分内优势度十分明显。相比而言,元宝枫、构树、桑树、五角枫、侧柏、栾树、构树、小叶朴等树种主要以劣态和绝对劣态分布,分化程度差异较大,在林分中处于被压状态。其它树种在空间结构单元中由于株数较少,树种优势度并不明显。从树种优势度和种群密度综合分析:栓皮栎和槲树的树种优势度都较大,但由于栓皮栎株数多,个体分布广,较槲树种群而言优势度更加明显。
3.2.3 点格局分析
(1)栓皮栎水源涵养林种群单种的分布格局从调查数据可以看出,样地内栓皮栎林的伴生树种中以元宝枫所占的比例最大,因此只对元宝枫的分布格局进行分析。由图3、图4可知,栓皮栎种群在所研究的尺度上呈显著性聚集分布,但并没有显示出最大聚集程度;元宝枫种群的分布格局在所研究的尺度上呈显著性聚集分布,当尺度 r=14 m时,最大聚集程度为L(r)=13.24。栓皮栎种群的聚集规模远大于取样面积,在所研究的尺度上,栓皮栎种群没有达到最大的聚集程度,说明今后研究栓皮栎种群的分布格局时,取样面积应该大于本研究的取样面积。

图3 栓皮栎种群格局分析
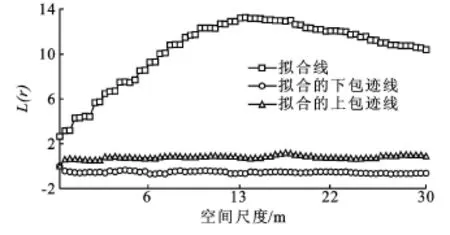
图4 元宝枫种群格局分析
(2)种群空间关联性分析。由图5可知,栓皮栎种群和元宝枫种群空间关联性在所研究的尺度上,当2 m<r<21 m时,二者之间关联性为显著性负相关,当r>21 m时,负相关性减弱。说明在小尺度上栓皮栎种群和元宝枫种群的生长势相当,分布均较广,二者之间竞争激烈,当尺度扩大到一定的范围,竞争减弱。

图5 种间关联性分析
4 讨论
林分结构决定林分功能,有什么样的林分结构,就有相应的林分功能[23]。采用地面三维激光扫描技术获取林分结构参数并配合混交度等空间结构指标来分析林分空间结构特征的方法,在国内外森林结构研究方面尚未多见。Tansey、Moorthy、Danson等研究表明,应用地面三维激光扫描技术对林分结构进行研究,在一次扫描过程中可以获取多棵树木数据,通过数据处理可精确测算出每一棵树二维坐标及树高、胸高断面积、冠径等测树因子,大大提高了数据的准确性和采集效率[12-13,24]。为此,将地面三维激光扫描技术引入林分空间结构特征研究,是我国数字化森林结构研究的未来发展方向。
本研究在地面三维激光扫描数据基础之上,利用树种混交度、大小比数等参数并结合空间点格局分析法从单木角度对西山林区栓皮栎水源涵养林林分结构规律进行了描述。首先,栓皮栎林的空间结构从树种组成上看,林分乔木层中共分布有16个树种,物种丰富度较高;林分垂直结构特征明显,可分为3个林层,呈现出典型的复层林结构特征。其次,从空间结构各指标来看:(1)林分平均混交度为0.32,主要以零度混交和弱度混交为主,林分内同种林木多聚集在一起,隔离度低,林分整体混交状况不良。以栓皮栎种群为例,其作为该林分的建群种与优势种,较其他伴生树种而言,混交度最差,元宝枫次之。除此之外,其他种群同种个体聚集程度较低,在某一空间结构单元中常与2株或2株以上不同树种相邻,树种混交度较大,混交状况良好。(2)栓皮栎水源涵养林林分胸径大小比数分布比较均匀,优势木相比而言较少。栓皮栎种群在空间结构单元中的优势度最大:一方面,栓皮栎种群在株数上占有较大比例,在胸径上处于优势地位;另一方面,在垂直结构中占据主林层有利的生态位,林木生长受相邻木的干扰较小,故栓皮栎种群将在种间竞争中继续保持优势地位。槲树、油松、元宝枫的优势度次之。其中,以槲树种群的胸径优势尤为明显,并占据着群落的最高层,在一定时期内槲树仍将保持其优势地位。但槲树株数所占比例较小,径级分布不连续,缺少中小径级的个体,当现存的槲树老化枯死后,树种优势将不复存在。其他伴生种群中除刺槐、油松等在竞争中具有一定优势外,绝大部分树种均处于劣势地位,且各树种个体数量少,径级分布不连续,优势度并不明显且更新较为困难。(3)通过单树种点格局分析得知:优势树种栓皮栎种群及主要伴生树种元宝枫种群在研究尺度(30 m)上均呈现出显著性聚集分布。在种群空间关联性分析上,栓皮栎种群和元宝枫种群在所研究尺度上,二者关联性为显著性负相关,说明二者中间竞争十分激烈。综合分析表明,在对西山林区栓皮栎水源涵养林进行经营时,可根据生态学原理,从优化空间结构的角度出发,采取相应的保护措施,保证非优势种群的顺利生长,以保持物种多样性和群落的稳定性,使林分结构更接近于自然状态的林分结构特征,从而提升林分的整体生态服务功能。
[1] Pretzsch H.Structural diversity as a result of silvicultural operations[M]//Olsthoorn A F M,Bartelink H H,Gardiner J J,et al.Management of mixed species forest:silviculture and economics.Dlo Institute for Forestry and Nature Research(IBN-DLO),Wageningen,1999:157-174.
[2] Hui G-Y,von Gasow K.Modern Forest Management of Germany[M].Beijing:China Science and Technology Press,2001.
[3] 岳永杰,余新晓.北京松山自然保护区蒙古栎林空间结构特征[J].应用生态学报,2009,20(8):1811-1816.
[4] 沈年华,万志洲.紫金山栓皮栋群落结构及物种多样性[J].浙江林学院学报,2009,26(5):696-700.
[5] 何加宜.北京妙峰山地区栓皮栋林物种多样性的研究[J].现代园林,2006(8):36-38.
[6] 吴晓莆,王志恒.北京山区栋林的群落结构与物种组成[J].生物多样性,2004,12(1):155-163.
[7] Király G,Brolly G.Tree height estimation methods for terrestrial laser scanning in a forest reserve[J].IAPRS Volume,2007,36:211-215.
[8] 赵阳,余新晓,信忠保,等.地面三维激光扫描技术在林业中的应用与展望[J].世界林业研究,2010,23(4):41-45.
[9] Jakob Weiβ.Application and statistical analysis of terrestrial laser scanning and forest growth simulations to determine selected characteristics ofDouglas-Firstands[J].Folia Forestalia Polonica:series A,2009,51(2):123-137.
[10] Gá bor B.Algorithms for stem mapping by means of terrestrial laser scanning[J].Acta Silv Lign Hung,2009,5(1):119-130.
[11] Koukoulas S,Blackburn G.Mapping individual tree location,height and species in broadleaved deciduous forest using airborne lidar and multi-spectral remotely sensed data[J].International Journal of Remote Sensing,2005,26(3):431-455.
[12] Tansey K,Selmes N.Estimating tree and stand variables in a Corsican Pine woodland from terrestrial laser scanner data[J].International Journal of Remost Sensing,2009,30(19):5195-5209.
[13] M oorthy I,Miller J R.Retrieving crown leaf area index from an individual tree using ground-based lidar data[J].Canadian Journal of Remote Sensing,2008,34(3):320-332.
[14] Li L,Hui S R,Hui G Y,et al.A Study on the minimum area of forest spatial investigation[J].Forest Resources Management,2007(2):47-51.
[15] 周红敏,惠刚盈,赵中华,等.林分空间结构分析中样地边界木的处理方法[J].林业科学,2009,45(2):1-5.
[16] Hui G Y,von Gadow K.Das winkelmass-theoretische ǜberlegungen zum optimalen standardwinke[J].Allgemeine Forst und Jagdzeitung,2002,173(9):173-177.
[17] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):177-181.
[18] 胡艳波,惠刚盈,戚继忠,等.吉林蛟河天然红松阔叶林的空间结构分析[J].林业科学研究,2003,16(5):523-530.
[19] 邓送求,闫家锋.南京紫金山枫香风景林空间结构分析[J].南京林业大学学报:自然科学版,2010,34(4):117-122.
[20] Ripleyb D.Modelling spatial pattern[J].Journal of the Royal Statistical Society:Series B,1977,39:178-212.
[21] Ripleyb D.Spatial statistics[M].New York:Wiley,1981.
[22] 常静,潘存德.梭梭-白梭梭群落优势种种群分布格局及其种间关系分析[J].新疆农业大学学报,2006,29(2):26-29.
[23] 袁正科,田育新.不同功能防护林类型的判别技术研究[J].林业科学,1998,11(3):1-5.
[24] Danson F M,Hetherington D.Forest canopy gap fraction from terrestrial laser scanning[J].Ieee Geoscience and Remote Sensing Letters,2007,4(1):157-160.
猜你喜欢
中国林业产业(2021年5期)2021-12-14
南方农业·下旬(2020年1期)2020-06-01
临床检验杂志(电子版)(2020年1期)2020-04-03
山东林业科技(2019年2期)2019-06-03
新农民(2019年9期)2019-02-19
现代园艺(2019年24期)2019-01-07
吉林农业(2019年9期)2019-01-06
现代农业科技(2009年21期)2018-09-03
农村百事通(2017年18期)2017-02-08
学苑创造·B版(2015年12期)2016-06-23
