
晶体苏氨酸和微囊苏氨酸对幼建鲤生长性能和消化吸收能力影响的比较研究
2011-03-28 10:35姜维丹周小秋
动物营养学报 2011年5期
冯 琳 彭 艳 刘 扬 姜 俊 姜维丹 胡 凯 周小秋*
目前国际市场鱼粉资源的短缺日趋严重,寻求新的蛋白质源替代鱼粉对水产养殖业具有重要的意义。研究表明,植物蛋白是比较有效的替代品[1],但与鱼粉相比,绝大部分植物蛋白不能完全满足鱼类的氨基酸需要[2]。苏氨酸是保证鱼类正常生长的一种必需氨基酸[3]。在鱼类常用的饲料原料中,苏氨酸被认为是继赖氨酸和蛋氨酸之后最重要的限制性氨基酸。研究表明,苏氨酸是高粱[4]、大麦[5]的第二限制性氨基酸,是谷物的第三限制性氨基酸[6]。
在陆生动物上已有较好的研究结果用于指导配合饲料中晶体氨基酸的添加量从而优化饲料的氨基酸平衡[7],而在水生动物上有关晶体氨基酸的利用效果存在较大争议[8-9]。研究表明,普通鲤[10]、日本对虾[11]对晶体氨基酸的利用效果不如完整蛋白质。晶体氨基酸的表观利用效率的下降与其吸收速率较快有关,吸收过快会导致过量的氨基酸分解代谢从而降低其利用效率[12]。因此,通过一种可降低消化速率的材料对晶体氨基酸进行包被处理,降低晶体氨基酸在肠黏膜转运位点的释放率,可能会大大提高氨基酸的利用效率[13]。本课题组前期的研究表明,在建鲤上使用包被处理的赖氨酸的效果优于其晶体形式[14]。然而,目前未见饲料中添加不同形式的苏氨酸对水生动物利用效果比较的研究报道。因此,本论文拟在幼建鲤的饲料中分别添加晶体苏氨酸和微囊苏氨酸(包被处理),比较 2种不同形式的苏氨酸对幼建鲤生长性能、肠道生长发育、肠道酶活力及肠道刷状缘酶活力的影响,以考察微囊苏氨酸的利用效果是否优于晶体氨基酸。
1 材料与方法
1.1 试验饲料
试验用晶体苏氨酸(L-苏氨酸)、微囊苏氨酸(包被处理)均由四川省畜科饲料有限公司提供。参考 Teshima等[15]的方法,采用高效液相色谱(HPLC)法测定晶体苏氨酸和微囊苏氨酸中苏氨酸的含量,实测值分别为99.21%和49.82%。配制2种试验饲料,即在基础饲料中分别添加晶体苏氨酸和微囊苏氨酸,并使苏氨酸含量(计算值)均达到 1.25%[参考 NRC(1993)[16]普通鲤苏氨酸需要量]。试验饲料中除苏氨酸以外的其他必需氨基酸模仿 32%全卵鸡蛋蛋白中的氨基酸模式添加[17],而吡哆醇、泛酸、肌醇和核黄素的营养需要量参考本实验室关于幼建鲤营养需要量的研究结果[18-21]。试验饲料经逐步添加6.0 mol/L的氢氧化钠(NaOH)调整 pH至 7.0,然后按照 Tibaldi等[22]的方法制成颗粒料,储藏于 -20℃的冰柜中备用。试验饲料组成及营养水平见表 1。

表1 试验饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of the experimental diets(DM basis)
1.2 试验鱼及饲养管理
试验在四川农业大学动物营养研究所水生动物营养试验场进行。鱼苗购自四川省雅安市鱼苗孵化场,鱼苗购回后进行 4周的适应性饲养后开始正式饲养试验。将 300尾平均体重为(13.61±0.02)g的健康幼建鲤随机分为2组(每组 3个重复,每个重复 50尾),分别饲喂添加晶体苏氨酸和微囊苏氨酸的试验饲料。采用连续充氧的循环水系统进行养殖,以重复为单位放养于水族箱(长 ×宽 ×高 =90 cm×30 cm×40 cm)中。试验鱼每天饲喂 6次,试验期 60 d。试验期间定期检查水质,每天日光灯光照 12 h,水温和 pH分别保持在(25±10)℃和7.0±0.3。
1.3 样品收集
每缸鱼在试验开始前和试验结束后分别称重,并记录试验期间每缸鱼的摄食量。试验开始前在试验鱼中随机选择 30尾鱼冷藏保存用于初始鱼体成分的分析,试验结束后每缸选择 5尾鱼冷藏保存用于终末鱼体成分分析,鱼体成分分析方法参考文献[23]。试验结束后,饥饿 12 h后每缸选择 15尾鱼解剖,分离肠道、肝胰脏和肌肉,称重后在液氮中快速冷冻,保存在 -70℃的冰箱中,用于肠蛋白质(IPC)和肝胰脏蛋白质含量(HPC)及肠道、肝胰脏和肌肉相关酶活力等指标的测定。每缸另选择 5尾鱼,分离其肠道,参考 Lin等[24]的方法测定其肠长和各肠段皱襞高度。最后 1次饲喂 6 h后,按照饲喂先后顺序每缸选择 5尾鱼尾静脉采血,一部分放置在用肝素钠预先处理的离心管中,4℃离心(3 000×g)15 min后立即测定血浆中的氨浓度,另一部分血样装在离心管中置于 4℃冰箱中静止过夜,4℃离心(3 000×g)15 min后分离血清,分装后-20℃保存待测血清中谷草转氨酶(GOT)和谷丙转氨酶(GPT)的活力。以冰冻的生理盐水为匀浆介质,用超声波分别粉碎肠道和肝胰脏、肌肉样品,然后 4℃离心(6 000×g)20 min,收集上清液。肠道和肝胰脏胰蛋白酶活力的测定参考 Hummel[25]的方法,脂肪酶和淀粉酶活力的测定参考 Furne等[26]的方法。各肠段碱性磷酸酶(AKP)、γ-谷氨酰转肽酶 (γ-GT)和 Na+,K+-ATP酶 (Na+,K+-ATPase)活力的测定分别参考 Bessey等[27]、Rosalki等[28]和 McCormic[29]的方法。肠道和肝胰脏的蛋白质含量采用 Bradford[30]的方法。肝胰脏、肌肉和血清中谷草转氨酶和谷丙转氨酶的活力均采用试剂盒进行测定(试剂盒由 BITKT临床试剂有限公司提供),血浆中氨浓度的测定参考 Tantikitti等[31]的方法。
1.4 计算公式
饲料转化率(FCR)=摄食量(g)/净增重(g);
特定生长率(SGR,%/d)=100×(ln终末体重 -ln初始体重)/试验天数;
蛋白质效率比(PER)=增重(g)/蛋白质摄入量(g);
蛋白质沉积率(PPV,%)=100×体蛋白增量(g)/蛋白质摄入量(g);
脂肪沉积率(LPV,%)=100×体脂肪增量(g)/脂肪摄入量(g);
肝体指数(HIS,%)=100×肝胰脏重量(g)/体重(g);
肠体指数(ISI,%)=100×肠道重量(g)/体重(g);
肠长指数(RGL,%)=100×肠长(cm)/体长(cm)。
1.5 苏氨酸体外释放速率的测定
参考 Lopez-Alvarado等[32]的方法分别测定晶体苏氨酸和微囊苏氨酸在 15、30、45、60、75、90、105、120、180和 240 min时的体外释放速率。具体操作方法为:取 50 mg的晶体或微囊苏氨酸于试管中,加入 10 mL硼酸缓冲液(25 mol/L,pH 8.5),用正亮氨酸作为内源标准物,在 25℃下不断振荡,分别在上述时间点过滤(滤纸孔径为0.45μm)后转入另一个试管,并用 HPLC法测定其中的苏氨酸含量。
1.6 数据处理与统计
试验结果以平均值 ±标准差表示,采用SPSS 11.5对数据进行 t检验,比较组间的差异显著性,以 P<0.05作为差异显著性判断标准。
2 结 果
2.1 晶体或微囊苏氨酸对幼建鲤生长性能的影响
由表 2可知,饲喂微囊苏氨酸的幼建鲤特定生长率和摄食量均显著高于饲喂微囊苏氨酸的幼建鲤(P<0.05),蛋白质沉积率和脂肪沉积率也呈现相同的结果。微囊苏氨酸组的蛋白质效率和饲料转化率与晶体苏氨酸组间差异不显著(P>0.05)。
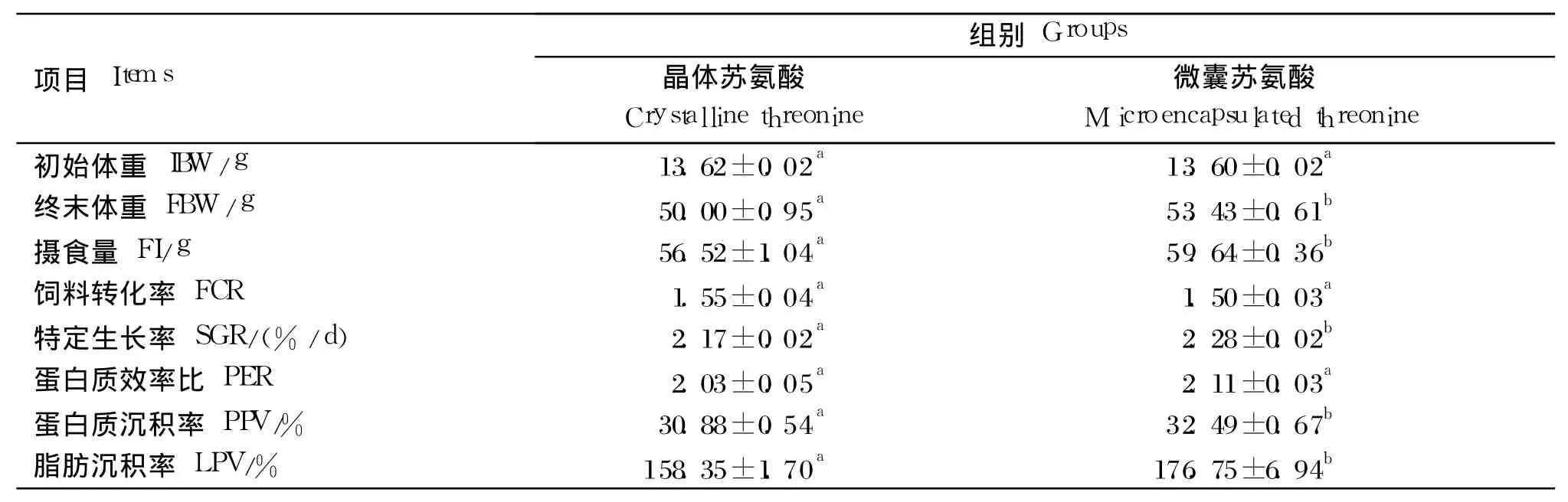
表2 晶体或微囊苏氨酸对幼建鲤生长性能的影响Table 2 Effects of crystalline or microencapsulated threonine on growth performance of juvenile Jian carp(Cyprinus carpio var.Jian)
2.2 晶体或微囊苏氨酸对幼建鲤肝胰脏、肌肉和血清中谷草转氨酶和谷丙转氨酶活力及血浆氨浓度的影响
肝胰脏、肌肉和血清中谷草转氨酶和谷丙转氨酶活力及血浆氨浓度见表 3。晶体苏氨酸组幼建鲤肝胰脏和肌肉中谷草转氨酶和谷丙转氨酶活力均显著高于微囊苏氨酸组(P<0.05)。相反的,晶体苏氨酸组幼建鲤血清中的谷草转氨酶和谷丙转氨酶活力显著低于微囊苏氨酸组(P<0.05)。此外,晶体氨基酸组幼建鲤血浆中氨浓度比微囊苏氨酸组增加了 9.6%,差异显著(P<0.05)。
2.3 晶体或微囊苏氨酸对幼建鲤消化吸收能力的影响
2.3.1 晶体或微囊苏氨酸对幼建鲤肝胰脏和肠重及蛋白质含量、肠长、肠长指数、肝体指数和肠体指数的影响
由表 4可知,微囊苏氨酸组幼建鲤的肝胰脏和肠道的重及蛋白质含量、肠长、肠长指数、肝体指数和肠体指数均显著高于晶体苏氨酸组(P<0.05)。

表3 晶体或微囊苏氨酸对幼建鲤肝胰脏、肌肉和血清中谷草转氨酶和谷丙转氨酶活力及血浆中氨浓度的影响Table 3 Effects of crystalline or microencapsulated threonine on activies of glutamate-oxaloacetate transaminase and glutamate-pyruvate transaminase in hepatopancreas,muscle and serum as well as plasma ammonia concentration of juvenile Jian carp(Cyprinus carpio var.Jian)

表4 晶体或微囊苏氨酸对幼建鲤肝胰脏和肠重及蛋白质含量、肠长、肠长指数、肝体指数和肠体指数的影响Table 4 Effects of crystalline or microencapsulated threonine on weight and protein content of hepatopancreas and intestine,intestine length,relative gut length,hepatosomatic index and intestosomatic index of juvenile Jian carp(Cyprinus carpio var.Jian)
2.3.2 晶体或微囊苏氨酸对幼建鲤各肠段皱襞高度的影响
由表 5可知,各组均以前肠的皱襞高度最高,中肠次之,后肠最低。组间比较发现,微囊苏氨酸组幼建鲤前肠和中肠皱襞高度分别比晶体氨基酸组高13.0%和 15.5%(P<0.05),而后肠皱襞高度组间差异不显著(P>0.05)。

表5 晶体或微囊苏氨酸对幼建鲤各肠段皱襞高度的影响Table 5 Effects of crystalline or microencapsulated threonine on fold height in different intestinal segments of juvenile Jian carp(Cyprinus carpio var.Jian) μm
2.3.3 晶体或微囊苏氨酸对幼建鲤肝胰脏和肠道消化酶的活力的影响
由表 6可知,微囊苏氨酸组肝胰脏和肠道胰蛋白酶以及肠道脂肪酶的活力均显著高于晶体苏氨酸组(P<0.05),而肝胰脏和肠道淀粉酶以及肝胰脏脂肪酶活力组间差异不显著(P>0.05)。

表6 晶体或微囊苏氨酸对幼建鲤肝胰脏和肠道消化酶活力的影响Table 6 Effects of crystalline or microencapsulated threonine on digestive enzyme activities in hepatopancreas and intestine of juvenile Jian carp(Cyprinus carpio var.Jian) U/g
2.3.4 晶体或微囊苏氨酸对幼建鲤各肠段Na+,K+-ATP酶、碱性磷酸酶和 γ-谷氨酰转肽酶活力的影响
由表 7可知,微囊苏氨酸组幼建鲤前肠、中肠、后肠碱性磷酸酶以及中肠、后肠 Na+,K+-ATP酶和γ-谷氨酰转肽酶的活力均高于晶体苏氨酸组(P<0.05)。晶体苏氨酸组前肠 γ-谷氨酰转肽酶和Na+,K+-ATP酶活力有低于微囊苏氨酸组的趋势,但差异不显著(P>0.05)。
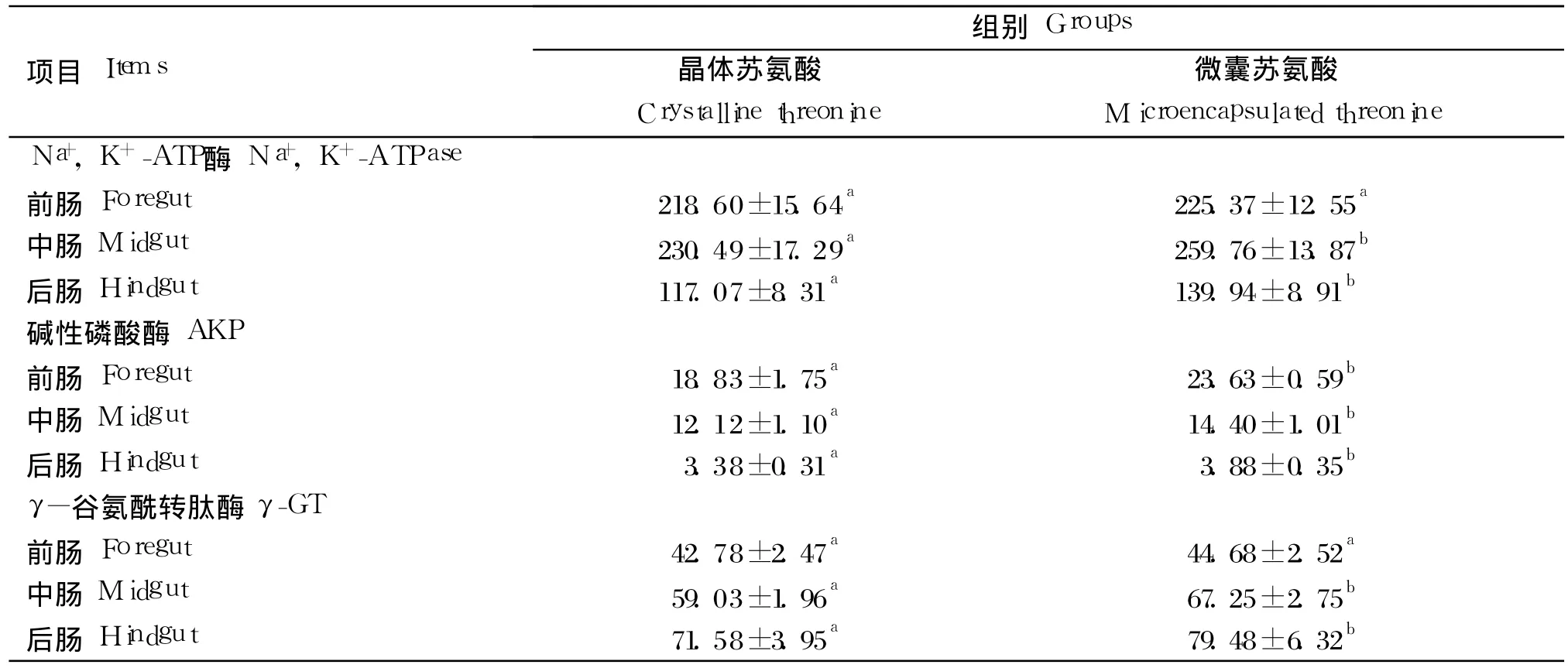
表7 晶体或微囊苏氨酸对幼建鲤各肠段Na+,K+-ATP酶、碱性磷酸酶和 γ-谷氨酰转肽酶活力的影响Table 7 Effects of crystalline or microencapsulated threonine on activities of Na+,K+-ATPase,alkaline phosphatase and γ-glutamyltransferase in different intestinal segments of juvenile Jian carp(Cyprinus carpio var.Jian) U/g
2.3.5 晶体或微囊苏氨酸的体外释放速率
由表 8可知,与微囊苏氨酸相比,晶体苏氨酸的释放速率较快,晶体苏氨酸在 15 min内就释放完全,而微囊苏氨酸相对要慢得多,在 15 min的时候释放了不到 20%,但 120 min后 2种形式的苏氨酸释放速率差异不显著(P>0.05)。
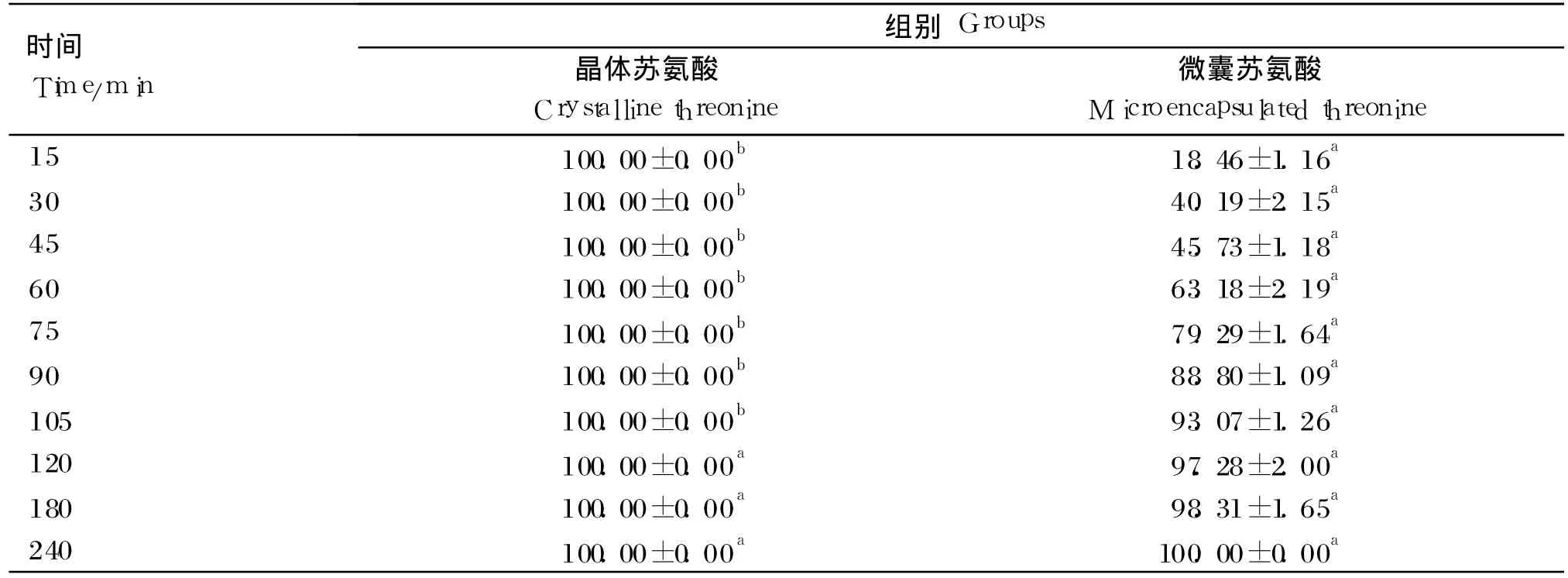
表8 晶体或微囊苏氨酸的体外释放速率Table 8 Release rate of crystalline or microencapsulated threonine in vitro %
3 讨 论
3.1 晶体苏氨酸和微囊苏氨酸对幼建鲤生长性能影响的比较
关于包被处理的氨基酸对水生动物生长的影响有少量报道。Zhou等[14]报道,包被处理的赖氨酸促进了幼建鲤的生长,与晶体赖氨酸相比增重提高了 14%,Alam等报道[11],与晶体氨基酸相比,包被处理的晶体氨基酸能显著提高日本对虾的生长。本研究结果表明,与晶体苏氨酸相比,微囊苏氨酸对幼建鲤的生长有更好的促进作用。包被处理的氨基酸促进鱼类的生长可能与提高其摄食量有关。上述文献中,包被处理的赖氨酸[14]和包被处理的晶体氨基酸[11]分别显著提高了幼建鲤和日本对虾的摄食量。本研究结果也发现,与晶体苏氨酸相比,包被处理的苏氨酸使幼建鲤摄食量提高了 5%。此外,本试验结果发现,2种形式的苏氨酸对幼建鲤的饲料转化率没有显著影响,但是包被处理的苏氨酸显著提高了幼建鲤的蛋白质沉积率和脂肪沉积率。这说明鱼类能更有效地利用包被处理的苏氨酸。本课题组在赖氨酸的研究上也得出了同样的结果[14]。因此,包被处理的苏氨酸比晶体苏氨酸能更有效促进幼建鲤的生长也可能与提高了营养物质在体内的沉积有关。
3.2 晶体苏氨酸和微囊苏氨酸对幼建鲤肝胰脏、肌肉和血清中谷草转氨酶和谷丙转氨酶活力及血浆氨浓度影响的比较
包被处理的苏氨酸比其晶体形式能更好地提高幼建鲤的蛋白质沉积率,可能与调节机体氨氮代谢,提高体蛋白合成能力有关。鱼类主要通过联合脱氨基作用满足机体脱氨基的需要,谷草转氨酶和谷丙转氨酶是蛋白质代谢中最重要的 2种酶,其活力的大小能反映氨基酸代谢强度的大小[33]。肝脏是蛋白质合成的重要部位,其转氨酶的活力可反映蛋白质的合成能力,而肌肉中转氨酶的活力主要反映蛋白质的分解代谢。从本试验结果可知,无论是肝胰脏还是肌肉中的谷草转氨酶和谷丙转氨酶活力,晶体苏氨酸组均显著高于微囊苏氨酸组,说明晶体苏氨酸组蛋白质代谢强度更大。推测出现这种差异的原因很可能是由于晶体苏氨酸组有更多的游离氨基酸被用于分解供能造成的。氨是硬骨鱼类蛋白质代谢的最主要的终产物。在本研究中,晶体苏氨酸组的幼建鲤血浆氨浓度显著高于微囊苏氨酸组,说明在微囊苏氨酸组的氨基酸平衡程度优于晶体苏氨酸组。目前未见不同形式的苏氨酸对血浆氨浓度影响的研究报道,但是在赖氨酸的研究上发现:与晶体赖氨酸相比,饲料中添加包被处理的赖氨酸使幼建鲤血浆氨浓度显著降低[14]。
3.3 晶体苏氨酸和微囊苏氨酸对幼建鲤消化吸收能力影响的比较
肠道消化酶的活力大小能反映鱼类对营养物质的消化能力[14]。本研究结果表明,与晶体苏氨酸相比,微囊苏氨酸能显著提高幼建鲤肠道蛋白酶和脂肪酶的活力,但是对淀粉酶活力影响差异不显著。这说明饲料中添加包被处理的苏氨酸能更有效地提高幼建鲤对蛋白质和脂肪的消化能力,但对淀粉的消化没有影响。胰腺是鱼类分泌蛋白酶和脂肪酶最主要的器官。本研究发现,微囊苏氨酸组幼建鲤肝胰脏中脂肪酶和淀粉酶的活力显著高于晶体苏氨酸组。这说明饲料中包被处理的苏氨酸能更有效地提高幼建鲤分泌消化酶的能力。有关不同形式的氨基酸对鱼类消化能力的影响目前仅见本课题组的 1篇研究,即:与晶体赖氨酸相比,包被处理的赖氨酸能更有效地提高幼建鲤蛋白酶、脂肪酶和淀粉酶的活力[14]。包被处理的苏氨酸能更有效地提高幼建鲤的消化能力与其更有效促进消化器官的生长发育有关。本研究结果表明:与晶体苏氨酸组相比,微囊苏氨酸组幼建鲤的肠重及蛋白质含量、肠长、肝胰脏重及蛋白质含量均显著增加。本研究结果与不同形式赖氨酸对幼建鲤消化器官生长发育的影响上的研究结果一致[14]。
肠道皱襞高度被认为是衡量水生动物吸收能力的重要标志。本研究结果显示,微囊苏氨酸组幼建鲤的前肠和中肠皱襞高度均高于晶体苏氨酸组,说明包被处理苏氨酸能更有效地增加幼建鲤肠道的吸收面积从而提高吸收能力。有研究认为绝大部分的氨基酸的吸收转运都要依靠 Na+,K+-ATP酶分解ATP产生能量[34],而碱性磷酸酶[35]、γ-谷氨酰转肽酶[27]与营养物质的吸收有关,其活力能间接地反映小肠的吸收能力。在本研究中,微囊苏氨酸组的幼建鲤前肠、中肠、后肠的碱性磷酸酶、Na+,K+-ATP酶和中肠、后肠的 γ-谷氨酰转肽酶活力均显著高于晶体苏氨酸组。这说明微囊苏氨酸对提高幼建鲤肠道的吸收能力的效果优于晶体苏氨酸。在幼建鲤饲料中添加包被处理的赖氨酸[14]时也得到了同样的结果。
3.4 晶体苏氨酸和微囊苏氨酸体外释放速率的比较
研究表明,晶体氨基酸的吸收速率过快可能导致氨基酸的过度分解代谢和利用率降低[12],这样就会使氨基酸在蛋白质的合成位点不平衡,因此被吸收的氨基酸更多地被分解代谢[36-37]。这可能正是本试验中 2种形式的苏氨酸利用效果不同的主要原因。体外释放速率的试验结果表明:晶体苏氨酸在缓冲液中 15 min后即全部溶解,而此时不到 20%的微囊苏氨酸被溶解,直到 120 min后才有 97.28%的苏氨酸被溶解。这说明包被处理能降低苏氨酸的释放速率。有关包被处理氨基酸释放速率的研究有少量报道。Schuhmacher等[38]的研究发现,晶体氨基酸被吸收的速率显著高于完整蛋白质结合形式分解的氨基酸。Zhou等[14]比较晶体赖氨酸和包被处理赖氨酸的研究发现,晶体赖氨酸在体外释放的速率显著大于包被处理的赖氨酸,包被处理的赖氨酸在体外完全释放需要的时间长达 3 h。氨基酸经过稳定化处理,能降低其溶解和吸收速率,增加在肠道的停留时间,利于氨基酸吸收平衡,便于更多的氨基酸用于蛋白质合成,从而促进鱼类生长[39]。上述结果说明,包被处理的苏氨酸能更好地提高幼建鲤的生长性能的部分原因在于通过特殊材料对苏氨酸进行包被处理后降低了苏氨酸的溶解速率,从而增加在肠道的滞留时间,更有利于氨基酸吸收平衡。
4 结 论
①与晶体苏氨酸相比,微囊苏氨酸能更有效地提高幼建鲤的生长性能和消化吸收能力。
②微囊苏氨酸在幼建鲤肠道中的吸收速率低于晶体苏氨酸。
[1] BODIN N,MAMBRINI M,WAUTERS J B,et al.Threonine requirements for rainbow trout(Oncorhynchus mykiss)and Atlantic salmon(Salmo salar)at the fry stage are similar[J].Aquaculture,2008,274:353-365.
[2] WILSON R P.Amino acids and proteins[M]//HALVERJE.Fish nutrition.2nd ed.New York:Academic Press Inc.,1989:112-153.
[3] GATLIN D M,BARROWS F T,BROWN P,et al.Expanding the utilization of sustainable plant products in aquafeeds:a review[J].Aquaculture Research,2007,38:551-579.
[4] COHEN R S,TANKSLEY T D.Limiting amino acids in sorghum for growing and finishing swine[J].Journal of Animal Science,1976,43:1028-1034.
[5] FULLER M F,MENNIE I,CROFTSR M J.The optimal amino acid supplementation of barley for the growing pig 2.Optimal additions of lysine and threonine for growth[J].British Journal of Nutrition,1979,41:333-340.
[6] GROSBACH D A,LEWISA J,PEO ER.An evaluation of threonine and isoleucine as the third and fourth limiting amino acids in corn for growing pigs[J].Journal of Animal Science,1985,60:487-494.
[7] LEWIS A J,BAYLEY H S.Amino acid bioavailability[M]//AMMERMAN C A,BAKER D,LEWIS A J.Bioavailability of nutrients for animals:amino acids,minerals and vitamins.San Diego,CA:Academic Press Inc.,1995:35-65.
[8] COWEY C B.Amino acid requirements of fish:acritical appraisal of present values[J].Aquaculture,1994,124:1-11.
[9] ZARATE D D,LOVELL R T,PAYNE M.Effects of feeding frequency and rate of stomach evacuation on utilization of dietary free and protein-bound lysine for growth by channel catfish Ictalurus punctatus[J].Aquaculture Nutrition,1999,5:17-22.
[10] MURAI T,AKIYAMA T,OGATA H.Effect of coating amino acids with casein supplemented to gelatin diet on plasma free amino acids of carp[J].Bulletin of the Japanese Society of Scientific Fisheries,1982,48:703-710.
[11] ALAM M S,TESHIMA S,KOSHIO S,et al.Effects of supplementation of coated crystalline amino acids on growth performance and body composition of juvenile kuruma shrimp Marsupenaeus japonicus[J].Aquaculture Nutrition,2004,10:309-316.
[12] LOVELL T.Nutrition of aquaculture species[J].Journal of Animal Science,1991,69:4193-4200.
[13] SEGOVIA-QUINTERO M A,REIGH R C.Coating crystalline methionine with tripalmitin-polyvinyl alcohol slows its absorption in the intestine of Nile tilapia,Oreochromis niloticus[J].Aquaculture,2004,238:355-367.
[14] ZHOU X Q,ZHAO C R,LIN Y.Compare the effect of diet supplementation with uncoated or coated lysine on juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Nutrition,2007,13:457-461.
[15] TESHIMA S,KANAZAWA A,YAMASHITA M.Dietary value of several proteins and supplemental amino acids for larvae of the prawn,Penaeus japonicus[J].Aquaculture,1986,51:225-235.
[16] NRC.Nutrient requirements of fish[S].Washington,D.C.:National Academies Press,1993.
[17] WILSON R P,ALLEN O W J R,ROBINSON E H,et al.Tryptophan and threonine requirements of fingerling channel catfish[J].Journal of Nutrition,1978,108:1595-1599.
[18] HE W,ZHOU X Q,FENG L,et al.Dietary pyridoxine requirement of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Nutrition,2009,15:402-408.
[19] WEN Z P,ZHOU X Q,FENG L,et al.Effect of dietary pantothenic acid supplement on growth,body composition and intestinal enzyme activities of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Nutrition,2009,15:470-476.
[20] JIANG W D,ZHOU X Q,FENG L,et al.Growth,digestive capacity and intestinal microflora of juvenile Jian carp(Cyprinus carpio var.Jian)fed graded levels of dietary inositol[J].Aquaculture Research,2009,40:955-962.
[21] LI W,ZHOU X Q,FENG L,et al.Effect of dietary riboflavin on growth,feed utilization,body composition and intestinal enzyme activity of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Nutrition,2010,16:137-143.
[22] TIBALDI E,LANARI D.Optimal dietary lysinelevels for growth and protein utilisation of fingerling sea bass(Dicentrarchus labrax L.)fed semipurified diets[J].Aquaculture,1991,95:297-304.
[23] AOAC.Official methods of analysis of the AOAC international[S].16th ed.Gaithersburg,MD:AOAC International,1998.
[24] LIN Y,ZHOU X Q.Dietary glutamine supplementation improves structure and function of intestine of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture,2006,256:389-394.
[25] HUMMEL B C W.A modified spectrophotometric determinations of chymotrypsin,trypsin,and thrombin[J].Canadian Journal of Physiology and Pharmacology,1959,37:1393-1399.
[26] FURNE M,HIDALGO M C,LOPEZA,et al.Digestive enzyme activities in Adriatic sturgeon Acipenser naccarii and rainbow trout Oncorhynchus mykiss.A comparative study[J].Aquaculture,2005,250:391-398.
[27] BESSEY O A,LOWRY O H,BROCK M J.Rapid coloric method for determination of alkaline phosphatase in five cubic millimeters of serum[J].Journal of Biological Chemistry,1946,164:321-329.
[28] ROSALKI S B,RAU D,LEHMANN D,et al.Determination of serum gamma-glutamyl transpeptidase activity and its clinical applications[J].Annals of Clinical Biochemistry,1970,7:143-147.
[29] MCCORMIC S D.Methods for nonlethal gill biopsy and measurements of Na+,K+-ATPase[J].Canadian Journal of Fisheries and Aquatic Sciences,1993,50:656-658.
[30] BRADFORD M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye-binding[J].Analytical Biochemistry,1976,72:248-254.
[31] TANTIKITTI C,CHIMSUNG N.Dietary lysine requirement of freshwater catfish(Mystus nemurus Cuv.&Val)[J].Aquaculture Research,2001,32:135-141.
[32] LOPEZ-ALVARADO J,LANGDON C J,TESHIMA S I,et al.Effects of coating and encapsulation of crystalline amino acids on leaching in larval feeds[J].Aquaculture,1994,122:335-346.
[33] BERGEG E,SVEIER H,LIED E.Nutrition of Atlantic salmon(Salmo salar):the requirement and metabolic effect of lysine[J].Comparative Biochemistry and Physiology Part A:Molecular&Integrative Physiology,1998,120:477-485.
[34] KLEIN S,COHN SM,ALPERS D H.The alimentary tract in nutrition[M]//SHILS M E.Modern nutrition in health and disease.Baltimore,MD:Williams and Wilkins,1998:605-630.
[35] VILLANUEVA J,VANACORE R,GOICOECHEA O,et al.Intestinal alkaline phosphatase of the fish Cyprinus carpio:regional distribution and membrane association[J].Journal of Experimental Zoology,1997,279:347-355.
[36] TRIPATHI P K,SINGH A.Carbaryl induced alterations in the reproduction and metabolism of freshwater snail Lymnaea acuminate[J].Pesticide Biochemistry and Physiology,2004,79:1-9.
[37] FARHANGI M,CARTER C G,HARDY R W.Growth,physiological and immunological responses of rainbow trout(Oncorhynchus mykiss)to different dietary inclusion levels of dehulled lupin(Lupinus angustifolius)[J].Aquaculture Research,2001,32:329-340.
[38] SCHUHMACHER A,WAX C,GROPP J M.Plasma amino acids in rainbow trout(Oncorhynchus mykiss)fed intact protein or a crystalline amino acid diet[J].Aquaculture,1997,151:15-28.
[39] MILLAMENA O M,BAUTISTA-TERUEL M N,KANAZAWA A.Methionine requirement of juvenile tiger shrimp Penaeus monodon Fabricius[J].Aquaculture,1996,143:403-410.
猜你喜欢
意林·全彩Color(2019年9期)2019-10-17
中国乳品工业(2018年10期)2018-11-16
科学与财富(2018年8期)2018-05-09
广东饲料(2016年2期)2016-12-01
广东饲料(2016年1期)2016-12-01
上海故事(2015年13期)2016-01-22
伴侣(2015年5期)2015-09-10
湖南饲料(2015年1期)2015-04-07
中国饲料(2015年12期)2015-01-25
湖南饲料(2014年4期)2014-08-26
