
虫生真菌的生物防治研究进展
2011-02-05 09:46杨运华杜开书石明旺
河南科技学院学报(自然科学版) 2011年1期
杨运华,杜开书,石明旺
(河南科技学院,河南新乡453003)
近年来,随着化学防治造成的环境污染等问题日趋严重(3R问题),生物治虫越来越来受到人们的重视.人们发现,在自然界中由真菌引起的流行病可自然制约着害虫的种群数量,维护生态平衡.此外,虫生真菌还有诸多优点:如种类多、应用期长、不伤害天敌、不易产生抗性及能快速大量生产等,故在生物防治中有着无可比拟的反复侵染性和生产便利性,因而越来越受到人们的重视.
1 虫生真菌资源及开发
虫生真菌资源极为丰富,据不完全统计,全世界迄今已记载的虫生真菌约100属800多种.而我国已发现的昆虫病原真菌也涉及40多个属400多种,其中虫草属真菌80种,捕食和寄生线虫的真菌10种,寄生昆虫的真菌215种,报道的新种达24种[1].主要分布鞭毛菌亚门(Mastigomycotina)壶菌纲(Chytridiomycetes)、接合菌亚门(Zygomycot)接合菌纲(Zygomycetes)、子囊菌亚门(Ascomycota)核菌纲(Pyrenomycetes)和不整囊菌纲(Plectomycetes)、担子菌亚门(Basidiomycota)层菌纲(Hymenomycetes)、半知菌亚门(Deuteromycota)丝孢菌纲(Hyphomycetes)和腔孢纲(Coelomycetes).其中比较常见的属、种及代表种见表1.
现今生物防治中应用较广主要为绿僵菌、白僵菌和蜡蚧轮枝菌等.我国利用白僵菌在早春和梅雨季节防治松毛虫防效可达90%以上[2].其与化学杀虫剂混用,可以提高防治效果.邱国森等用白僵菌与其他药剂(灭幼脲)混合防治马尾松毛虫,防治效果可达98%以上,并具有持续作用[3].绿僵菌喷洒蝗虫或蚱蜢,死亡率可以达到70%-90%,并对非目标害虫无害[4].蜡蚧轮枝菌可自然寄生蚜虫和介壳虫,李国霞等用其防治大棚蔬菜蚜虫,效果可以达到93.1%[5].国外已经把虫瘟霉应用带小菜蛾的防治中[6].
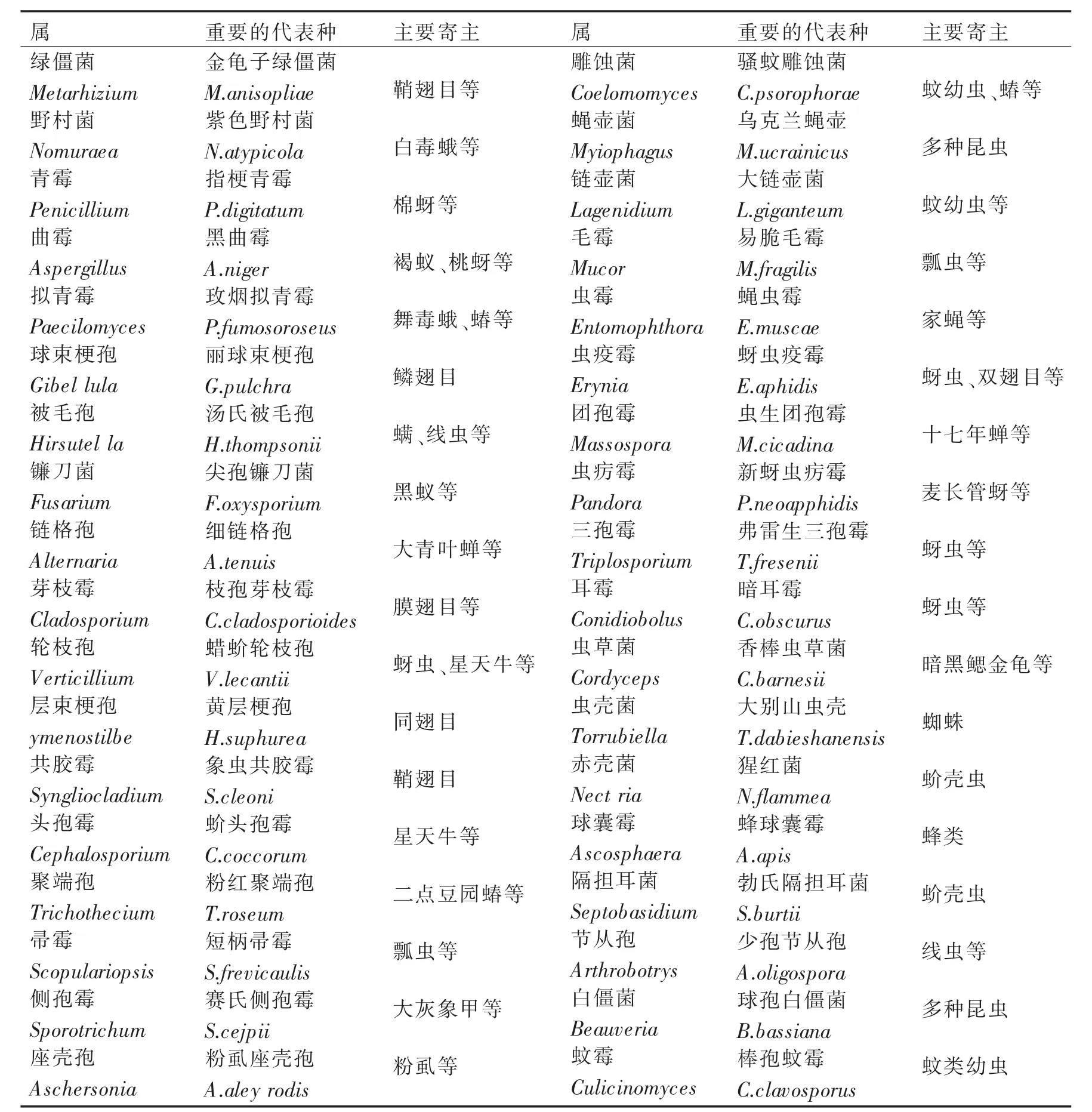
表1 虫生真菌常见的36个属
2 虫生真菌的入侵及致病机制
虫生真菌的入侵主要分4个阶段:首先在昆虫体表形成芽管(germtube),然后芽管分化形成附着胞(appressorium)等侵染结构,接着附着胞凭借机械和酶的压力形成侵染钉(penet ration peg)进入寄主体内,最后菌丝在寄主体内大量繁殖,导致寄主昆虫死亡[7].虫生真菌的入侵是机械压力和酶共同作用的结果[8].虫生真菌侵入过程中发现的主要酶类有蛋白质酶、几丁质酶、酯酶、脂酶和淀粉酶[9]等.并且酶的出现有一定的顺序,首先出现的是蛋白水解酶,接着才是几丁质酶、脂酶等.蛋白水解酶在入侵过程中起着重要作用,它使几丁质暴露,诱导几丁质酶对几丁质作用.研究发现,真菌侵入后产生的毒素是引起昆虫致死的重要因子.如白僵菌和绿僵菌在侵入后产生的破坏素,可破坏寄主的免疫系统或使其逐步死亡,也可以激活鳞翅目昆虫肌肉细胞中的Ca2+通道,引起昆虫痉挛麻痹,最终导致死亡[10].当然,昆虫致死过程与酶也有着重要的联系.
2.1 毒素
1952 年Aoki和Shimodara首次发现虫生真菌能产生有杀虫活性的代谢产物.接着Kodaira通过绿僵菌致死家蚕的实验证明了毒素存在,首次纯化出了破坏素(destruxin)[11].此后,又有大量的毒素被陆续分离纯化.Mollier将这些毒素分为两大类:低分子量毒素和高分子蛋白毒素[12].
低分子量毒素主要有环缩肽类、色素类及有机酸类等,其中环缩肽类(如白僵菌素)可使寄主组织崩解,肌肉弛缓,及对鳞翅目昆虫表现为强直性麻痹,色素类(如卵孢素)可抑制虫尸中的细菌繁殖,有机酸类(如类草酸晶体)可破坏昆虫的体壁,有明显的协同作用;高分子蛋白毒素主要有酶类毒素和非酶类蛋白毒素两大类,其中酶类毒素(如胞外蛋白酶)可降解昆虫体壁,有辅助致死作用,非酶类蛋白毒素(如糖蛋白)可直接将寄主昆虫致死,对幼虫表现高毒性并伴随有体壁黑化现象[13].
2.2 酶
虫生真菌侵入昆虫过程中产生的酶除辅助侵入外还与菌株的毒力有密切关系.Gupta等的实验结果表明酶与毒力之间存在较强的相关性[14],酶活愈高,寄主死亡率愈大,LT50愈低.樊美珍等研究结果表明蛋白酶产量与毒力之同存在相关关系[15].El-sayed等通过试验证明真菌对某些昆虫的幼虫毒力与几丁质酶的活性有关[16].
3 虫生真菌的毒力稳定性研究
虫生真菌的田间测试经常产生不一致的结果[17].这说明其毒力的不稳定性,由于真菌毒力最终要表现为杀虫,所以阐明杀虫机理颇为重要.王中康[18]等已分别就这方面作了详细综述.研究证明,无论是虫生真菌自身内部因素,还是环境因子都会影响杀虫效果[19].菌株来源对同种昆虫毒力往往差异较大,同一菌株对不同寄主或同一寄主不同虫龄的毒力也有所不同[20].这就表现出真菌毒力的复杂性.为了保证致死效果,我们应注意以下两方面:
1)提高毒力从自然界中大量选育野生优良菌株;选育抗逆性优质突变株;优化产酶条件;遗传改良菌株,主要是基因工程技术及分子克隆技术的应用.
2)稳定毒力单孢分离株具相对稳定的毒力,Samsinakova等用此法获得毒力相对稳定且高效的天然突变株[21].也可在不影响菌种毒力的前提下进行遗传改良,利用准性生殖或原生质体融合形成杂合二倍体.也能采取生物工程技术,引入毒力相关基因,培育出稳定的高毒力菌株.方卫国等成功借助根癌农杆菌把毒力相关基因CDEP-1、Bbchitl和抗除草剂基因bar转入了球孢白僵菌[22],St.Leger等把易于检测的报告基因GUS转入了虫生真菌[23].
4 虫生真菌研究展望
迄今国内外已成功地通过基因工程技术将相关毒力基因导入昆虫病原真菌,从而显著提高其毒力和杀虫效率,未来还需要继续通过基因工程技术提高虫生真菌的环境稳定性,扩大其应用领域;应用细胞工程技术(原生质体及菌丝融合)培育高产、高毒和易流行扩散的新菌株,这将是今后虫生真菌遗传育种的主要研究方向;应用分子生物学对虫生真菌相关毒性蛋白酶克隆和测序结果表明,其重要致病性基因是以基因组的形式存在的,通过基因重组可显著提高虫生真菌的致病速率;大量生产时,最适剂型的应用、技术合成等问题,及微生物杀虫剂的市场定位问题,还要通过政府部门或者资助中介机构等推动真菌治虫的大规模应用;同时必须研制延长菌剂孢子寿命保护剂,期限至少1 a.
[1] 王利军,谭万忠,罗华东,等.虫生真菌及其在害虫生物控制中的应用现状与展望[J].河南农业科学,2010,(4):119-125.
[2] 王清海,万平平,黄玉杰,等.虫生真菌在害虫生物防治中的应用研究[J].山东科学,2005,18(4):37-41.
[3] 邱国森.灭幼脲3号与白僵菌混合剂对马尾松毛虫的防治效果[J].广东林业科技,2000,16(3):21-27.
[4] Lomer CJ,Bateman R P,Johnson DL,et al.Biological control of locusts and grasshopp-ers[J].Annu Rev Entomo1,2001,46:667-702.
[5] 李国霞,郭友中,茅洪新,等.应用蜡蚧轮枝菌对大棚蔬菜蚜虫防治效果的研究[J].中国病毒学,2000,(15):262.
[6] Furlong MJ,Pell J K.Horizontal transmission of entomopathogenic fungi by the diamondback moth[J].Biol Control,2001,(22):288-299.
[7] Wang C S,Hu G,St Leger R J.Differential gene expression by Metarhizium anisopliae growing in rootexudate and host(Manduc ASexta)cuticle or hemolymph reveals mechanisms of physiological adaptation[J].Fungal Genetics and Biology,2005,42:704-718.
[8] 王记祥,马良进.虫生真菌在农林害虫生物防治中的应用[J].浙江林业学报,2009,26(2):286-291.
[9] Robert A,Messing-AI-aidfroosk.Acid production by Metarhiziumm anisoplia:Efects on virulence against mosquitoes and on detection of invitroamylase,protease and lipase Activity[J].Invertebr Pathol,1985,45:9-15.
[10] 吕丁丁,李增智,王成树.虫生真菌分子致病机理及基因工程改造研究进展[J].微生物学通报,2008,35(3):443-449.
[11] Kodaira T.Studies on the new toxic substance to insects,destruxiNAAnd B,produced by Oospora destructor.I.Isolation and purification of destruxin Aand B[J].Agr Biol Chem,l961,26:36-42.
[12] Mollier P,Lagnel J,Quiot J M,et a1.Cytotoxic activity in culture filtrates from the entomopathogenic fungus BeauveriA Sulfurescens[J].Journal of Invertebrate Pathology,1994,64:208-213
[13] 李建庆.虫生真菌毒素的性质与分离研究进展[J].滨州学院学报,2005,21(3):53-58.
[14] Gupt AS C,Leathers T D,EL-Sayed G N,et al.Production of degradatiou enzyme by,Metarhizium anisoplia during growth on defined media and insect cuticle[J].Experi Mycol,1991,15:310-3l5.
[15] 樊美珍,胡锦江,李农昌,等.球孢白僵苗胞外蛋白酶及其与毒力关系的研究[J].微生物学通报,1994,21(4):202-206.
[16] EL-Sayed G N,Goudron T A,lgnoffcm.Chtinolytic activity and virulence associated with native and mutant isolates of an entomopathogenic fungus Nomuraca[J].J Invertebr Path of,1989,54:394-403.
[17] E.Quesada-Morage,Vey A.Intra-specific variation in virulence and in vitro production of macromolecular toxins active against locust among beauveria bassian A Strains and effects of in vivo and in Vitro passage on these factors[J].Biocontrol Science and Technology,2003,13:323-340.
[18] 王中康,殷幼平,彭国雄,等.杀虫真菌功能基因克隆和菌株改良研究进展[J].中国虫生真菌研究与应用,2003(5):18-28.
[19] 赵杰宏,刘爱英,梁宗琦,等.虫生真菌的毒力稳定性研究[J].西南农业学报,2004,17(6):790-793.
[20] 何恒果,李正跃,陈斌,等.虫生真菌对害虫防治的研究与应用[J].云南农业大学学报,2004,19(2):167-173.
[21] Samsinakova A,Kalalov AS.Journal of Inbertebrate Pathology[J].1983,42:156-161.
[22] 方卫国,裴炎.昆虫病原真菌降解寄主体壁酶基因的克隆及球孢白僵菌高毒力重组菌株的获得[D].重庆:西南农业大学,2003.
[23] St.Leger R J,Shimizu S,Joshi L,et al.Co-transformation of metarhizium anisopliae by electropo ration or using the gene gun to produce stable GU Strans formats[J].FEMS Microbiology Letter,1995,131:289-294.
猜你喜欢
食品工业科技(2021年22期)2021-11-14
生物技术通报(2021年4期)2021-05-14
农药科学与管理(2019年6期)2019-11-23
湖北林业科技(2019年2期)2019-05-05
中国酿造(2017年8期)2017-09-03
中成药(2017年5期)2017-06-13
上海农业学报(2016年2期)2016-10-27
广西林业科学(2016年4期)2016-03-16
华南农业大学学报(2015年5期)2015-12-04
华南农业大学学报(2015年5期)2015-12-04
