
优雅蝈螽细胞减数分裂期染色体行为的观察
2010-09-04 08:22:40关晓利
山西大同大学学报(自然科学版) 2010年1期
关晓利
(山西大同大学继续教育学院,山西大同 037009)
优雅蝈螽细胞减数分裂期染色体行为的观察
关晓利
(山西大同大学继续教育学院,山西大同 037009)
本文以螽斯精巢为实验材料,经过低渗、固定、压片、空气干燥、改良品红染色制备染色体标本,对优雅蝈螽减数分裂染色体行为进行了研究.结果:优雅蝈螽精巢细胞染色体2n♂=31,染色体几乎全部为近端部着丝粒染色体,其性别决定机制为:XO为雄性,XX为雌性.减数分裂双线期同源染色体形成环状或8字状,端部交叉的结构.
优雅蝈螽 减数分裂 染色体
昆虫染色体的研究,包括分析染色体核型、有丝分裂、减数分裂、特殊染色体、染色体的结构与数量变异,不仅对昆虫分类系统的建立、近缘种的区分、昆虫种类演替规律及种间种内进化,而且对遗传学、害虫的遗传防治及害虫抗药性机理具有重要意义.我国在昆虫染色体研究方面,对许多种类的染色体核型进行了研究,取得了一些成果[1].研究优雅蝈螽减数分裂染色体行为,将为螽斯生殖生理、系统发育等提供资料.
1 材料与方法
1.1 材料
花鸟市场购买优雅蝈螽末龄若虫,室内饲养.
1.2 方法
1.2.1 取材、预处理、固定及保存
将螽斯雄虫活体注射0.05%浓度的秋水仙素溶液,每只个体注射2~8μL,以虫体大小而定,6~8 h后,进行活体解剖,在虫体腹部侧膜处剪一小口,用尖头镊子夹出精巢,除去精巢表面的结缔组织,将精巢浸入低渗液中5~10min,转入卡诺固定液 (甲醇:冰醋酸=3:1)中约8~12 h,其间换固定液一次,然后移至70%酒精中,于4℃冰箱保存备用[2].
1.2.2 染色体制片
本文采用压片法制作染色体玻片标本.用镊子夹取精巢小管,至于培养皿中,切取其末端膨大部分,放到洗净的载玻片上,滴加一滴40%~ 60%的冰醋酸溶液,软化10~20 min,加盖片,覆滤纸,施加压力,使细胞分散,然后液氮冰冻揭片,无尘处自然干燥.用品红染色,蒸馏水冲洗,干燥,镜检[2].
1.2.3 染色体的观察拍照
将干燥后的染色体标本置于光学显微镜下观察,选择好的分裂相进行拍照分析.
2 优雅蝈螽精原细胞减数分裂的染色体行为
减数分裂是一种特殊的有丝分裂形式,仅发生于有性生殖细胞形成过程中的某个阶段.减数分裂的主要特征是细胞仅进行一次染色体的复制,随后进行两次分裂.两次分裂分别称为减数分裂Ⅰ期和减数分裂Ⅱ期.两性生殖细胞经过减数分裂,各自的染色体数减少一半.
减数分裂与有丝分裂的一个主要区别在于其复杂的分裂前期,在粗线期同源染色体形成联会复合体,在双线期同源染色体分离,表现出特殊的染色体形态行为.
2.1 细线期(Leptonema)
细线期为前期Ⅰ的开始阶段,染色体细长缠绕在一起,难以区分各条染色体 (图1-1).每条染色体复制为两条姐妹染色单体,但在显微镜下还分辨不出.
2.2 偶线期(zygotene)
染色体开始浓缩,但仍然是细丝状.在这个时期,同源染色体开始配对,但还没有完全配对,在同源染色体之间存在着间隙(图1-2).
2.3 粗线期(Pachytene)
同源染色体完全配对,形成二价染色体,在此过程中,染色体进一步浓缩,变粗变短,此时已经可以区分各个二价染色体,同时我们在这一时期观察到了分叉现象的存在(图1-3).
2.4 双线期(Diplotene)
同源染色体间的联会逐渐解体,两侧轴从中部开始分离.在早双线期,同源染色体刚刚开始分离,联会复合体之间大部分还相互连接,只有一小部分已经分开 (图1-4).晚双线期同源染色体之间几乎完全分离,只是在中间部分还相互交联,有的已经完全分开,并排排列在一起.有的形成8字状(图1-5).
2.5 终变期(Diakinesis)
同源染色体已经完全分离,形成了两条非常明显的染色体,染色体形成短棒状结构(图1-6).
2.6 中期Ⅰ(MetaphaseⅠ)
染色体进一步缩短变粗,几乎成为点状.此时的同源染色体在纺锤丝的牵引下,排列在赤道板周围(图1-7).
2.7 后期Ⅰ(NanphaseⅠ)
同源染色体相互分离并向两极移动,但染色体只是刚刚开始分离,还没有到达细胞的两极 (图1-8).
2.8 早末期Ⅰ(Early telophase)
同源染色体到达细胞两极,并且开始去凝集(图1-9).
2.9 中期Ⅱ(MetaphaseⅡ),末期Ⅱ(TelophaseⅡ)
减数分裂中期Ⅱ与有丝分裂的过程相似,但仔细观察会发现:第一细胞内没有成对的同源染色体,第二染色体有时会以奇数的形式出现.(图1-10),在末期Ⅱ,染色体去凝集,有成为染色质状态.此时,原来的亲本细胞经过减数分裂得到四个子细胞(图1-11).
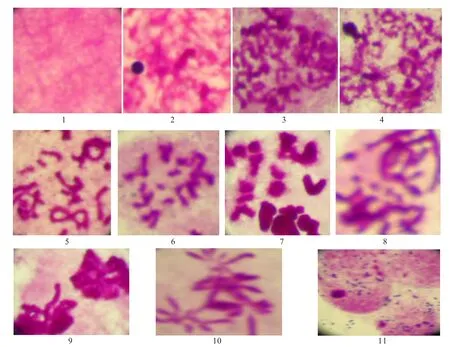
图1 优雅蝈螽精原细胞减数分裂染色体
3 讨论
优雅蝈螽属于螽斯科Tettigoniidae,染色体数目为2n=31,对于其他做过染色体研究的螽斯总科昆虫的染色体数目多为2n♂=29,但也有几种与优雅蝈螽相同,如亚螽亚科Saginae、以及盾螽亚科Decticinae和露螽亚科Phaneropterinae中的个别种类.从图中可以看到大部分的染色体没有很明显的中部着丝粒,因此本人认为优雅蝈螽精原细胞染色体几乎全部为近端部着丝粒染色体,性别决定机制为XO♂/XX♀[3].
优雅蝈螽的X染色体呈现异固缩现象,在减数分裂的前期就已经浓缩.在减数分裂的中期Ⅰ分裂相中可以很清楚的看到,X染色体与其它的染色体有所不同,它可能是中部着丝粒[4].
我国昆虫染色体的研究经历几十年发展,取得了一批重大的成果,填补了较多的研究空白.如今我国直翅目蝗总科的细胞分裂学初步形成了体系[5],随着对半翅目、同翅目、双翅目等昆虫染色体核型研究的展开[6],染色体分析已成为分析系统关系的一个新手段.然而,我们也必须看到,我国昆虫染色体的研究整体水平同国际差距仍然很大,研究规模也不符合我国昆虫种类繁多,分布广泛的现状,同害虫治理的实际需求好存在一定的差距.因此,探寻有效的核型分析方法应是昆虫染色体深入研究的首要任务.
[1]张礼生,张青文,蔡青年,等.中国昆虫染色体研究现状与展望[J].昆虫学报,2003,46(6):773-782.
[2]常岩林,廉振民.露螽科四种昆虫的核型研究[J].遗传,2000,22(6):385-388.
[3]Warchalowska SliwaE,BugrovAG.Maryanska Nadachowska AKaryotyoes and the banding pattern so for some species of the Phaneropterinae(Orthoptera,Tettigonioidea)[J].FoliiaBiol(Krakow),1996,44:5-10.
[4]WarchalowskaSliwaE,BugrovAG,Maryanska NadachowskaA.Karyotypes of the three species of the genera Poecilimon Fisch.and IsophyaBr.W.(Orthoptera,Tettigonioidea,Phaneropterinae)from the NorthCancasus[J].Caryologia,1995,48:27-34.
[5]马恩波,欧晓红,乔格侠.蝗总科染色体研究及科级综合比较(直翅目)[J].昆虫分类学报,2000,22(1):6-9.
[6]付建业,张青文,蔡青年,等.甜菜夜蛾细胞分裂期染色体的观察[J].昆虫学报,2005,48(1):135-138.
Observation on M eiotic C hromosomes of the Gampsodeis G ratiosa
GUAN Xiao-li
(School of Continual Education,ShanxiDatong University,Datong Shanxi,037009)
Chromosome preparations weremade from testis tissue of the Gampsodeis gratiosa with the airdrymethod.Themorphological characteristics and behaviors of the chromosomes at various phases ofmitosis and meiosis were studied.The chromosome number of the Gampsodeis gratiosawas determined as 2n=31,all chromosomes are acrocentric.During the diplotene the synaptonemal complexes showed rings crossed.
Gampsodeis gratiosa;meiosis;chromosome
Q969.26+3.2
A
〔编辑 杨德兵〕
1674-0874(2010)01-0059-03
2009-07-18
关晓利(1963-),女,山西大同人,副教授,研究方向:微生物学.
猜你喜欢
东坡赤壁诗词(2022年4期)2022-10-30 12:18:17
生物技术进展(2022年1期)2022-02-11 03:45:46
植物保护(2019年4期)2019-09-04 09:34:39
科学之谜(2018年9期)2018-12-17 09:05:08
安徽农业科学(2018年22期)2018-05-14 08:59:50
课外生活(小学1-3年级)(2017年11期)2017-12-11 12:33:05
小学生作文·小学中高年级适用(2016年4期)2017-01-16 11:07:17
小学时代(2016年11期)2016-10-11 06:14:51
生物学教学(2016年12期)2016-04-10 23:04:28
青少年科技博览(中学版)(2015年3期)2015-06-01 19:08:33
