
高效产脂肪酶菌株C1的分离鉴定与酶学性质研究
2010-09-04 03:00于玉凤陆兆新卢亚萍刘茜茜陈舟舟
湖南农业科学 2010年23期
于玉凤 ,陆兆新 ,卢亚萍 , ,刘茜茜 ,陈舟舟
(1.南京农业大学生命科学学院,江苏 南京 210095;2.南京农业大学食品科技学院,江苏 南京 210095)
脂肪酶(Lipase,EC3.1.1.3)是一类重要的甘油酯键水解酶,广泛存在于动物、植物各种组织及微生物中,可以在油水界面上催化脂类物质分解、合成和酯交换反应,是最早研究的酶类之一[1]。微生物脂肪酶具有种类多,比动物脂肪酶更广的作用pH值和作用温度范围,便于进行工业生产和获取高纯度制剂等优点,在食品、制革、饲料等多个行业得到广泛应用[2-3]。不同种属的微生物所产生的脂肪酶具有不同的催化特性,发现具有不同催化特性(如耐高温、耐低温、耐有机溶剂等)的脂肪酶产生菌以满足不同的工业生产需求具有重要意义。从自然界中寻找产生优良特性的脂肪酶的菌株,是促进脂肪酶研究与应用的有效途径之一。笔者从油菜地土壤中分离到一株高效产脂肪酶菌株C1,对其进行了菌种鉴定,并对其酶学性质进行了初步研究,旨在为脂肪酶工程菌株的研究提供科学依据。
1 材料与方法
1.1 材料
1.1.1 土样本实验所用土壤样品采自江苏农科院多年种植油菜的试验田。
1.1.2 培养基 富集培养基:酵母抽提物2 g/L,橄榄油 5 g/L,K2HPO41 g/L,MgSO40.1 g/L,(NH4)2SO41 g/L,NaCl 0.5 g/L,自然 pH;初筛培养基:乳化橄榄油 10 g/L,K2HPO41 g/L,MgSO40.1 g/L,(NH4)2SO41 g/L,KCl 0.5 g/L,Agar 15 g/L,自然 pH;罗丹明 B复筛培养基:LB平板中添加0.001%罗丹明B和1%乳化橄榄油;产酶基础培养基:葡萄糖 10 g/L,酵母抽提物5 g/L,K2HPO42 g/L,橄榄油10 g/L,MgSO40.5 g/L,自然 pH。
1.2 方法
1.2.1 菌种的筛选 称取5 g土壤置于45 mL生理盐水中,加玻璃珠打散。移取5 mL至45 mL富集培养基,37℃培养2~4 d。梯度稀释富集培养液,涂布到以橄榄油为唯一碳源的初筛平板上,37℃培养2~4 d。从初筛平板上挑取有水解圈的菌落,用无菌牙签接种到罗丹明B复筛平板上,7℃培养2~4 d。观察并测量菌落周围荧光圈的大小。选取荧光圈直径/菌落直径较大的菌落,划线分离后接种到LB中过夜培养,种子液加入产酶基础培养基中,30℃振荡培养,24 h后每隔6 h取样测酶活。
1.2.2 菌种的鉴定 根据《伯杰细菌鉴定手册》对筛选的菌体进行表型和生理生化的鉴定。并以C1基因组DNA为模板,16S-F(5'-AGAGTTTGATCCTGGCTCAG-3') 和 16S-R(5'-GGTTACCTTGTTACGACTT-3')为引物,通过PCR扩增其16Sr RNA的编码基因,测序后进行序列比对。
1.2.3 酶活性的测定 参照Lee et al.的脂肪酶活测定方法[4],略作修改。取10μL发酵上清液,100 μL 10 mM pNPB(对硝基苯丁酯)加入到2.89 mL PBS(pH 7.4)中,37℃进行酶促反应,测定产物对硝基苯酚在410nm处的吸光值。以已知浓度(0.01、0.05、0.10、0.15、0.20 μmol)的对硝基苯酚作标准曲线,计算酶活。每分钟产生1μmol对硝基苯酚所需的脂肪酶定义为1个酶活单位。
(1)酶的最适温度和温度稳定性测定:分别于35~80℃温度下测定去除菌体的上清发酵液中脂肪酶活力,以测定脂肪酶最适反应温度。将酶液分别于40~80℃温度冰浴中保温1 h和5 h,立即取出冷却后测定其酶活。
(2)酶的最适pH值和pH稳定性测定:配制不同pH值的0.5 mmol/L缓冲液,包括PBS缓冲液(pH 5.0~8.0),Gly-NaOH 缓冲液(pH 9.0~12.0)。在37℃下对不同pH值的缓冲液测定酶活力,测定酶的最适pH。将酶在37℃,pH值分别为5.0、6.0、7.0、8.0、9.0、10.0、11.0 的缓冲液中(1∶1/v/v)保温 1 h 和5 h,然后在 37℃,pH 7.4,0.5 mmol/L PBS 缓冲液中测定残余酶活力,检测酶的pH稳定性。
(3)酶的有机耐性测定:选取极性不同的有机溶剂,分别取各种有机溶剂25μL与75μL的酶液充分混合后,放在37℃下保温,处理0.5、4.5、11.5 h后测定其残余酶活。以加入相同体积的蒸馏水为对照,并定义为100%酶活。
1.2.4 酶的金属离子耐性测定 选取常见的金属离子,将0.1 mol/L相对应的金属离子10μL与90 μL酶液充分混合后,放入37℃条件下保温1 h,测定其残余酶活。同样以加入相同体积的蒸馏水为对照,并定义为100%酶活。
2 结果与分析
2.1 产脂肪酶菌株的筛选与鉴定
2.1.1 菌株的筛选与生理生化特性鉴定 通过多轮初筛和复筛,得到40株产脂肪酶的细菌。从罗丹明B复筛平板上挑取有明显水解圈的菌落(见图1),接种到基础产酶培养基中培养。在培养至72 h时,其中的一株产酶量最高,酶活可达到28.5 U/mL,命名为C1。C1菌落呈白色,表面光滑不透明,突起,易挑起,有淡淡的异味,结合其生理生化特性(见表1),初步鉴定C1为Burkholderia cepacia。

表1 菌株C1的生理生化特性
2.1.2 16Sr DNA序列分析及系统进化树的建立经上海生工生物工程技术服务有限公司测序,菌株C1的16Sr DNA序列为2 084 bp,将所得序列在NCBI网页上进行BLAST比对,结果发现C1的16Sr DNA序列与数据库中Burkholderia cepacia strain S31 LipA(lipA)和 LipB(lipB)(FJ638612.1)比较接近,相似性达99%。从BLAST比对的结果中选取来源不同的序列,利用软件MEGA4.1绘制进化树,从图2可以看出,进化树中菌株C1与菌株Burkholderia cepacia(FJ638612.1)位于同一分支,亲缘关系最近。根据菌株C1的形态特征、生理生化特征和16Sr DNA序列分析及系统发育构建,鉴定菌株 C1为 Burkholderia cepacia。

2.2 酶学性质研究
2.2.1 酶的最适温度和温度稳定性 如图3所示,酶在65℃时达到最高酶活力,即酶的最适温度为65℃。

如图4所示,处理1 h后,其酶活下降较少,其中70℃下仍残留酶活91.2%;处理5 h后,60℃下残留酶活76.2%,随后酶活大幅下降,可见酶的耐温性较好。

2.2.2 酶的最适pH值和pH稳定性 由于在pH值9.0~11.0条件下,底物分解非常迅速,使得测定数据严重失真。因此,图5中只反映pH值5.0~8.0条件对脂肪酶活力的影响,其中在pH值8.0时,酶活最高,比pH值7.4时酶活多出41.8%,酶的最适pH值为8.0。

由图6可知,脂肪酶在pH值8时稳定性最高,在pH值5~10范围内温育1 h,酶活残留量均保持在80.0%以上,只在pH值>10以后,酶活大幅下降,只残留32.9%;温育5 h后,在pH值9处仍然残留59.8%的酶活,说明酶具有较稳定的pH耐性。

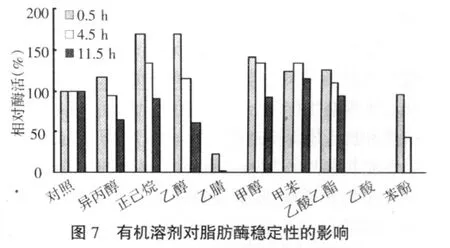
2.2.3 酶的有机耐性 如图7所示,酶对正己烷、乙醇、甲醇耐性最好,对乙酸乙酯、甲苯和异丙醇耐性较好,而对苯酚、乙腈和乙酸耐性较差。
2.2.4 酶的金属离子耐性 如图8所示,K+、Mg2+对脂肪酶活性有明显促进作用;Na+、Mn2+、Zn2+和 Fe3+对脂肪酶活性有微弱的抑制作用,残留酶活保持在80.0%以上;而Ca2+和Cu2+对脂肪酶活性有严重抑制作用,残留酶活在35.0%以下。
3 结论

图8 金属离子对脂肪酶稳定性的影响
本研究从土壤中筛选得到了一株高效产脂肪酶的菌株C1,经形态特征、生理生化特征和16Sr DNA序列分析鉴定为Burkholderia cepacia。对其所产脂肪酶的酶学性质进行研究,结果表明,酶的最适温度为65℃,最适pH值为8.0。酶的温度耐性较好,70℃下温育1 h残留酶活91.2%,60℃ 下温育5 h残留酶活76.2%;酶的pH耐性优良,在pH值5.0~10.0范围内处理1 h,酶活残留量均保持在80.0%以上,在pH值5.0~9.0范围内处理5 h,酶活残留量均在59.8%以上;酶对正己烷、乙醇、甲醇具优良的耐性,对乙酸乙酯、甲苯和异丙醇耐性较好,而对苯酚、乙腈和乙酸耐性较差;K+、Mg2+对脂肪酶活性有明显促进作用,Na+、Mn2+、Zn2+和 Fe3+对脂肪酶活性有微弱的抑制作用,而Ca2+和Cu2+对脂肪酶活性有严重抑制作用。与吴继光[5]、赵春雷[6]等的研究相比,该脂肪酶的温度耐性明显较强,且经过对其发酵条件的优化,其产酶活性可大幅提高,此脂肪酶具有较大的工业应用价值。
[1]Jaeger K E,Eggert T.Lipases for biotechnology[J].Current Opinion in Biotechnology,2002,13(4):390-397.
[2]刘海洲,刘均洪,张媛媛,等.微生物脂肪酶的最新应用研究进展[J].食品研究与开发,2009,30(1):141-143.
[3]GUPTA R,GUPTA N,RATHI P.Bacterial lipases:an overview of production,purification and biochemical properties [J].Applied Microbiology and Biotechnology,2004,64:763-781.
[4]Lee D W,Koh Y S,Kim K J,et al.Isolation and characterization of a thermophilic lipase from Bacillus thermoleovorans ID-1[J].FEMSMicrobiology Letters,1999,179:393-400.
[5]吴继光,舒正玉,程蓝骍,等.耐受有机溶剂洋葱伯克霍尔德菌ZYB002全细胞脂肪酶酶学性质 [J].微生物学通报,2010,37(1):2-6.
[6]赵春雷,闫丽娟,谢振荣,等.一株耐热脂肪酶产生菌的筛选及酶学性质研究[J].生物技术通报,2010,(2):185-188.
猜你喜欢
昆明医科大学学报(2021年5期)2021-07-22
中华养生保健(2020年3期)2020-11-16
快乐语文(2020年26期)2020-10-16
World Journal of Clinical Cases(2019年6期)2019-04-17
中国洗涤用品工业(2017年2期)2017-04-16
国外医药(抗生素分册)(2016年2期)2016-07-12
中国塑料(2016年7期)2016-04-16
中国卫生标准管理(2015年3期)2016-01-14
中国粮油学报(2014年7期)2014-02-06
植物营养与肥料学报(2010年4期)2010-11-06
