
饲粮中色氨酸对0~3周龄肉仔鸡生长性能及血液生化指标的影响
2010-08-09 02:38李丽娟
饲料工业 2010年20期
王 鹏 王 安 李丽娟
色氨酸(tryptophan,Trp)于1902年由Hokinst首先从酪蛋白中分离获得,化学名称为α-氨基-β-吲哚丙酸,分子式C11H12N2O2,色氨酸是动物维持和生长的必需氨基酸,是构成机体蛋白质的基本组成成分之一,具有多种生理功能,但动物体内不能合成,需从饲料中供给。在玉米-豆粕型配合日粮中,色氨酸较易缺乏,需要额外添加(Goihl,1996)。色氨酸(Trp)的作用是1910年以玉米蛋白质饲喂白鼠,生长发育受阻,添加色氨酸后提高了生长速度才明确的。色氨酸在玉米饲料中为第二限制性氨基酸,在大豆饼饲料中为第三限制性氨基酸。我国传统的玉米-豆粕(饼)型为主的肉鸡配合饲料中,色氨酸较易缺乏,故色氨酸被认定为是继蛋氨酸、赖氨酸之后的第三限制性氨基酸。
近年来,国内外在肉鸡对蛋氨酸、赖氨酸的需要量方面的研究较多,对色氨酸的研究甚少,但在国外的某些发达国家(如日本),色氨酸已广泛地用于畜禽饲料。本文通过在0~3周龄AA肉仔鸡日粮中添加不同水平的色氨酸来研究探讨对其生长性能及血液生化指标的影响,确定玉米-豆粕型饲粮饲喂的AA肉仔鸡色氨酸最适添加量,为实际生产提供理论依据,从而加快国内色氨酸添加剂的应用步伐,推动饲料工业和畜牧业的发展。
1 材料与方法
1.1 试验材料
色氨酸(纯度99.8%,购于华达饲料公司),1日龄AA肉仔鸡购于哈尔滨远大孵化场。
1.2 试验动物与试验设计
选用1日龄体重相近、健康AA肉仔鸡180只,体重经方差分析差异不显著,采用单因子试验设计,随机分为5个处理,每个处理6个重复,每个重复6只鸡。试验肉鸡饲养于3层重叠式金属鸡笼中,饲养前对鸡舍进行严格消毒。试验期间保证鸡舍较好的通风和光照,全期自由采食和饮水,按照常规方法和程序进行饲养管理和免疫,具体实施办法参照《现代肉鸡生产手册》进行。每周空腹称重并记录饲料消耗量。
1.3 试验日粮
I组为对照组,饲喂玉米-豆粕型基础日粮(色氨酸含量为0.06%),II组~V组为试验组,分别饲喂玉米-豆粕型日粮,并额外添加0.03%、0.06%、0.09%、0.12%色氨酸,日粮配制参照NRC(1994)和中华人民共和国农业行业肉鸡饲养标准(2004),基础日粮组成及营养水平见表1。
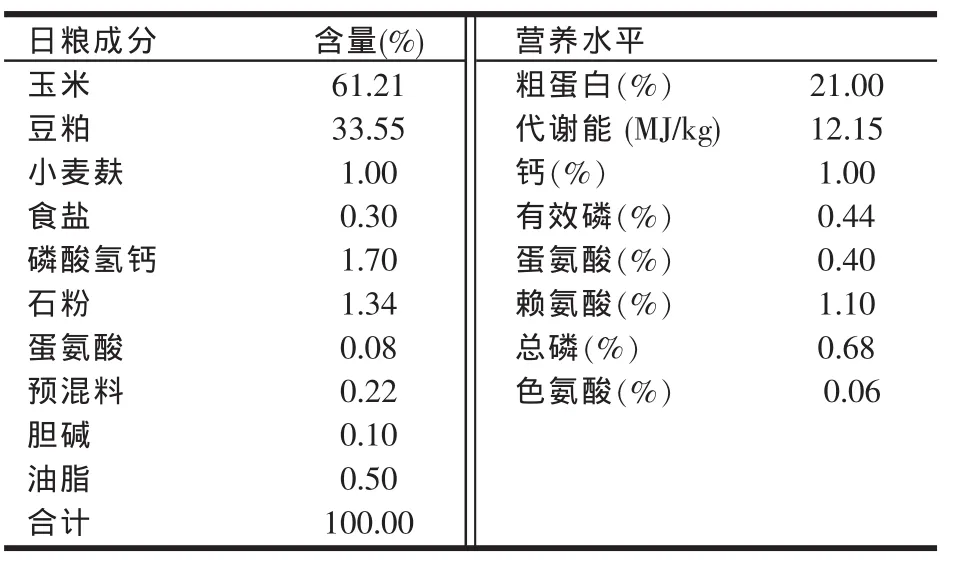
表1 基础日粮组成及营养水平(干物质基础)
1.4 样品采集和指标测定
每周称量肉仔鸡体重,统计饲料消耗量,到试验结束时计算平均日增重、日采食量和料重比。在试验的第21 d分别从每个重复中选取1只体重接近本组平均值的肉鸡进行颈静脉采血5 ml,放置于离心机中,3 500 r/min离心15 min,取上层血清分装于EP管中,放置于-20℃保存待用。血液生化指标分别测定总蛋白(TP)、总胆固醇(CHOL)、甘油三酯(TG)、葡萄糖(Glu)和尿素氮(BUN),血液生化指标的测定均在意大利产FULLY全自动生化分析仪上进行,所用试剂均采用中生北控公司所提供的试剂盒。
1.5 数据处理
全部数据采用SAS6.12版软件的Anova法进行方差分析,差异显著则进行Duncan's法多重比较分析,数据均以平均值±标准差形式表示。
2 结果
2.1 日粮中添加色氨酸对肉仔鸡生长性能的影响

表2 日粮中添加色氨酸对肉仔鸡生长性能的影响
由表2可以看出,日粮中添加色氨酸对0~3周龄的肉仔鸡无论是采食量,还是日增重都有显著的影响。和对照组相比,大部分试验组的平均日采食量和平均日增重差异明显(P<0.05)。但是第V组的平均日采食量、平均日增重与对照组相比差异不显著(P>0.05)。各试验组中日增重以第III组效果最好,好于其他各试验组。料重比各组之间差异不明显(P>0.05),第III组的料重比最低,饲喂效果最好,而添加水平较高的第V组和未添加色氨酸的第I组数据基本一致。由此可以看出,在饲粮中添加适当浓度的色氨酸可以不同程度降低料重比而提高饲粮的利用率。
2.2 日粮中添加色氨酸对0~3周龄AA肉仔鸡血液生化指标的影响(见表3)
从表3可以看出,第II、III、IV和V组血清中总蛋白含量均高于第I组,分别比第I组提高了9.4%、17%、8.3%和3.8%,其中各试验组中,以第III组含量最高,并且和第Ⅰ组相比差异显著(P<0.05),但与其他各试验组之间存在差异,但不显著(P>0.05),从总体来看,本试验中肉仔鸡血清总蛋白(TP)含量随着色氨酸添加量逐渐增大呈现先升高后降低的变化趋势。
AA肉仔鸡血清中总胆固醇的含量第II、III、IV和第V组相对于第I组都有不同程度降低。其中第III组和第IV组与第I组相比差异显著(P<0.05),分别比第I组降低了20.3%和19.4%,这两组之间并无显著差异(P>0.05)。第II组和第Ⅴ组总胆固醇含量虽低于第I组,但是差异不显著(P>0.05)。从表中数据可以看出,本试验中各组肉仔鸡血清中总胆固醇含量随着色氨酸添加水平的增加有不同程度的降低,但是当添加量升高到一定水平后又表现为上升的趋势。
肉仔鸡血清中甘油三酯(TG)含量趋势为各试验组均低于对照组,其中含量最低为第Ⅳ组,相比于第Ⅰ组降低了6%。但各试验组数据和第I组数据无显著差异,且各试验组间差异也不显著(P>0.05)。
各试验组肉仔鸡血清中葡萄糖(Glu)含量均低于第I组,第III组和第I组之间差异显著(P<0.05),比第I组含量降低了4.2%。其余各组间无显著差异(P>0.05)。从整体上来看,本试验中肉仔鸡血清中葡萄糖含量随着色氨酸添加量的增加表现出下降的趋势,但当添加量达到一定水平后开始出现上升的现象。
日粮中添加色氨酸以后可不同程度的降低肉仔鸡血清中尿素氮(BUN)的含量。其中含量最低的是第III组,比第I组含量降低了8.5%。各试验组中含量最高的是第V组,第II、IV组和III组含量比较接近。各试验组肉仔鸡血清尿素氮的含量均低于第I组,但差异均不显著(P>0.05)。
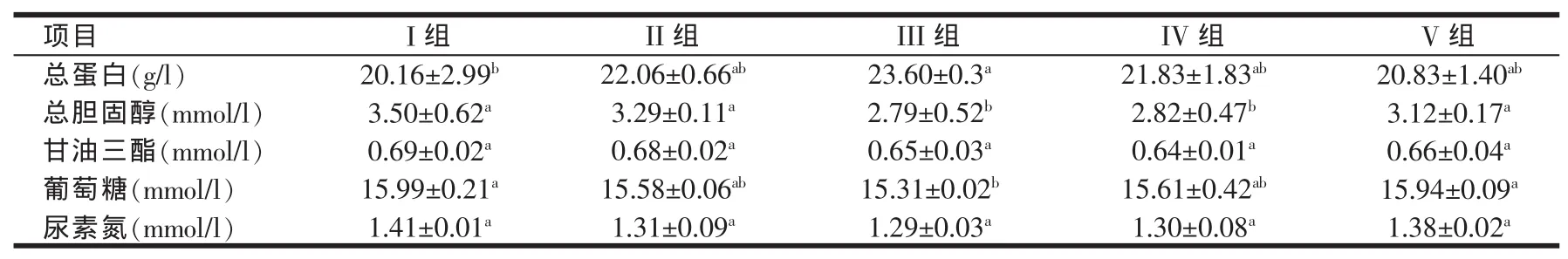
表3 日粮中添加色氨酸对肉仔鸡血液生化指标的影响
3 讨论
3.1 日粮中添加色氨酸对0~3周龄肉仔鸡生长性能的影响
据 Walstrom(1985)、Bory等(1985)、Sato等(1987)、Burgoon等(1990)的报道表明,饲粮中色氨酸水平微弱变化(0.03%)可显著影响断奶仔猪的采食量、日增重和饲料转化率。本试验研究表明,在0~3周龄AA肉仔鸡生长期间日粮中添加色氨酸可影响其生长性能,其中平均日采食量和平均日增重变化与对照组相比较为明显。在色氨酸添加量为0.06%水平时,差异显著(P<0.05)。料重比虽然没有显著变化,但饲粮中添加色氨酸的试验组除第V组外与对照组相比仍有不同程度的降低。日采食量随着色氨酸添加水平不同表现为“低-高-低”的变化趋势,其可能原因在于色氨酸在动物体内代谢产物5-羟色胺的作用。5-羟色胺能够调节人和动物的进食量(Rossi等,1991;Ponter等,1993),另外一些研究认为,色氨酸可能是直接作用于氨基酸受体或通过增加外周5-羟色胺含量而调节采食作用(Lacy等,1986)。Crozo(2005)研究认为,在适当水平下添加色氨酸对肉仔鸡有促进采食和生长作用,缺乏色氨酸则会引起采食量下降,体增重降低和饲料利用率下降。NRC标准(1994)推荐的0~3周龄肉仔鸡色氨酸需要量为0.20%,而本次试验研究表明,0~3周龄肉仔鸡饲喂玉米-豆粕型日粮时,适宜色氨酸添加水平为0.06%。造成结果差异较大的原因可能是由于基础日粮组成、日粮可消化率、试验方法研究手段不同所致。另外,由于色氨酸在测定过程中容易受到水解和所使用的药品试剂等因素的影响,因而饲料中色氨酸的准确测定非常困难,从而造成不同研究者得出的色氨酸需要量结果差异变化很大(张庆生等,2009)。
3.2 日粮中添加色氨酸对0~3周龄肉仔鸡血液生化指标的影响
动物体内物质代谢所需的原料以及代谢的产物的运输都要依靠血液循环来完成,因此,血液生化指标反映了动物体内物质合成与分解的详细情况。色氨酸一方面可以作为蛋白合成的原料影响蛋白的合成;另一方面色氨酸也可以通过一些其他作用来促进蛋白合成。Challakondan等(1980)在饲料中添加色氨酸,发现鼠肝细胞内蛋白合成增加;Herschel等(1984)发现,色氨酸可以改变核膜的渗透性,并使核膜上有关酶的活性增强,从而促进了肝细胞的蛋白合成。Borg等(1987)和 Malmlof等(1988)研究指出,血浆尿素氮(BUN)可以比较准确地反映动物体内蛋白质代谢的情况和日粮中氨基酸的平衡情况。苏有键等(2005)研究结果表明,在低蛋白的饲料中添加色氨酸可以不同程度的降低仔猪血浆尿素氮的水平。色氨酸对于脂类的代谢影响比较复杂。Akiba等(1992)研究报道,给鸡补充色氨酸会使鸡肝总脂下降,同时会影响血浆胆固醇和甘油三酯的含量,其可能机理是色氨酸可以减少肝脏脂肪的合成。色氨酸对糖类(碳水化合物)代谢作用相关的研究并不多,多数集中在色氨酸与其他大分子中性氨基酸的比值(TRP/LNAA)上,Venziale等(1981)研究发现,色氨酸及其代谢产物对动物体内糖原异生有抑制作用,这可能就是色氨酸可以降低动物血糖含量的原因之一。
本试验结果表明,在0~3周龄肉仔鸡玉米-豆粕型日粮中添加不同水平色氨酸,肉仔鸡血清中总蛋白含量有不同程度的提高,但是当添加到一定剂量的时候又出现了下降的情况。而血糖和血脂的含量则随着色氨酸的添加出现了不同程度的下降,血清尿素氮的含量也随着色氨酸的添加出现了下降的情况。
4 结论
①在本试验条件下,在玉米-豆粕型日粮中添加色氨酸提高了0~3周龄AA肉仔鸡的日增重,并降低了料重比。
②随着色氨酸的添加,AA肉仔鸡血清中蛋白含量均有不同程度的提高,而血脂和血糖则有不同程度的下降。而色氨酸添加水平过高会导致血液中蛋白含量降低,血脂和血糖升高。
③综合分析,建议在饲喂AA肉仔鸡的玉米-豆粕型日粮中添加0.06%水平的色氨酸为宜。
[1]林映才,蒋宗勇,余德谦,等.断奶仔猪色氨酸需求参数的研究[J].动物营养学报,1999,11(3):44-50.
[2]李剑欣,张绪梅,徐琪寿.色氨酸的生理生化作用及其应用[J].氨基酸和生物资源,2005,27(3):58-62.
[3]吴妙宗,蔡辉益.0~3周龄肉用仔鸡色氨酸营养需要及其与烟酸互作关系的研究[J].动物营养学报,2002(02):5-8.
[4]尹靖东.色氨酸对猪摄食和生长的调控及机理研究进展[M].猪营养与饲料研究进展,2007:79-85.
[5]崔芹,崔山.色氨酸营养研究进展[J].中国饲料,2003(15):20-23
[6]吴妙宗,蔡辉益.色氨酸营养代谢研究进展.[J].饲料与添加剂,2001(11):46-48.
[7]苏有键,李德发,邢建军,等.在低蛋白日粮中添加色氨酸对仔猪生产性能及血清游离氨基酸和尿素氮的影响[J].中国畜牧杂志,2005,41(1):26-28.
[8]张庆生,黎观红.日粮中色氨酸水平对0~4周龄泰和乌骨鸡生产性能的影响[J].江西农业学报,2009,21(7):158-161.
[9]Sidransky H,kurl H N,Holmes S C,et al.Tryptophan binding to nuclei of rat liver andhepatoma[J].J.Nutr.Biochem.,1995,6(2):73-79.
[10]Henry Y,Seve B,Colleaux Y,et al.Interactive effects of dietary levels of tryptophan and protein on voluntary feed intake and growth performance in pigs,in relation to plasma free amino acids and hypothalamic serotonin[J].J.Anim.Sci.,1992,70(6):1873-1887.
[11]Walstorm RC,et al.Efficiency of suppenntal trypiophan,threo nien,isole ucine and methionine of wauning pigs fed a low-protein lysine-supplemeated,corn-sunflower meal diet,1985.
[12]Borg B S,et al.Effect of tryptophan supplementation of a low protein corn-sunflower meal diet for young growing pigs.Journal of animal science,1985,61(1):300
[13]Akiba Y,Takahashi K,Horiguchi M,et al.L~tryptophan alleviates fatty liver and modifies hepatic microsomal mixed function oxidase in laying hens[J].Comp.Biochem.Physiol.Comp.Physiol.,1992,102(4):769-774.
[14]VenezialeCM,DonofrioJC,HansenJB,etal.Specificradioimmunoassays forrabbitliverfructose bisphosphatase,pyruvate kinase,and glycerol-3-phosphate dehydrogenase [J].MethodsEnzymol.,1981,74:210-232.
[15]Rao P R,Rao A B,Ramakrishnan S.Biochemical mechanism of induction of fatty liver by tryptophan[J].Indian J.Exp.Biol.,1980,18(11):1335-1336.
猜你喜欢
动物营养学报(2022年11期)2022-12-08
北方牧业(2022年9期)2022-11-22
特产研究(2021年6期)2021-12-08
养殖与饲料(2021年5期)2021-11-30
现代畜牧科技(2021年10期)2021-11-19
家畜生态学报(2020年7期)2020-07-14
河南农业科学(2019年9期)2019-09-24
兽医导刊(2019年20期)2019-08-19
今日畜牧兽医(2018年9期)2018-02-13
农村百事通(2016年6期)2016-05-14
