
受烟草花叶病毒侵染程度不同的烤烟叶片光合特性的变化
2010-07-31 07:41:46倪国仕章新军毕庆文
中国烟草科学 2010年5期
倪国仕,章新军,毕庆文,王 瑞*
(1.湖北省烟草公司恩施州公司,湖北 恩施 445000;2.湖北中烟工业有限责任公司,武汉 430051)
烟草花叶病毒(Tobacco mosaic virus, TMV)是烟草病毒病主要的毒源之一,烟草感染TMV后,常出现花叶或坏死及矮化畸形等症状,正常的生理功能受到不同程度的影响。光合作用是绿色植物最重要的生命活动,植物感染病毒后光合作用一般都会受到影响,已有研究表明,受病毒侵染后植物的光合速率明显下降,叶绿体受到破坏,叶绿素含量降低,光合电子传递活力下降,光合磷酸化作用降低[1-4]。Wilhelmova等[5]曾研究了受TMV侵染后烟草叶中光合色素含量和叶绿素荧光参数的变化,刘国顺等[6]曾报道受 TMV侵染后烟草叶中保护酶活性和超微机构的变化。本试验研究了受TMV侵染程度不同的烟草叶片光合气体交换参数、光合色素含量以及叶绿素荧光参数的变化,旨在进一步探讨病毒影响植物光合作用的生理生化机理。
1 材料与方法
1.1 供试材料
试验于 2009年在湖北省恩施州宣恩县烟草示范田进行,材料为烤烟品种云烟87。黄棕壤土,土层深厚,质地偏粘,pH为5.6,有机质为24.2 g/kg,碱解氮为 135.15 mg/kg,速效磷为 21.42 mg/kg,速效钾为195.00 mg/kg。在烟株旺长期,通过感病症状确定主要受TMV侵染的烟株,选取正常株、轻度感病株(有轻微花叶症状,感病植株只在叶片上形成黄绿相间的花叶病驳,叶片基本不变,但叶肉会出现明显变薄或厚薄不均的症状)和重度感病株(花叶明显且叶片严重畸形叶缘逐渐形成缺刻并向下卷曲,皱缩扭曲)。
1.2 测定项目与方法
1.2.1 光合气体交换参数的测定 在晴天上午9:30—10:30,采用美国Licor公司生产的LI-6400便携式光合测定系统测定净光合速率(Pn),气孔导度(Gs),胞间 CO2浓度(Ci),气孔限制值(Ls)=1-Ci/Co(Co为叶外空气 CO2浓度)。人工控制条件为:CO2浓度 380 μmo1/mol,25℃,光照强度 l 000 μmol/(m2·s)。每个处理3株,选取中部叶(自下而上第 10位叶)进行光合参数的测定,每株测定 2次,取其平均值。
1.2.2 叶绿素荧光参数的测定 同步用英国Hansatech公司生产的 FMS2脉冲调制式荧光仪测定光适应条件下的稳态荧光(Fs)和稳态最大荧光(Fm′),叶片经暗适应30 min后初始荧光(F0)和最大荧光(Fm),并计算PSⅡ活性(Fv/F0)、PSⅡ最大光能转换效率(Fv/Fm)、光化学猝灭系数(qp)、非光化学猝灭系数(NPQ)和电子传递的量子产率(ΦPSⅡ),分别按下式计算[7]:Fv/F0=(Fm-F0)/F0,Fv/Fm=(Fm-F0)/Fm,qp=(Fm′-Fs)/(Fm′-F0),NPQ=(Fm-Fm′)/Fm′,ΦPSⅡ=(Fm-Fs)/Fm′。
1.2.3 光合色素的测定 同步用直径1 cm打孔器取鲜叶,用80%丙酮研磨提取,以UV-754型分光光度计(上海亚研电子科技有限公司)测定 663、646、和470 nm处的OD值,然后计算单位面积叶绿素和类胡萝卜素含量[8]。
2 结 果
2.1 TMV侵染对烟草叶片气体交换参数的影响
从表1可见,与正常株相比,轻度感病株叶片净光合速率(Pn)、气孔导度(Gs)和胞间CO2浓度(Ci)分别下降了 26.05%、73.08%和 55.76%,气孔限制值(Ls)上升了 151.85%,差异均达到显著水平;重度感病株叶片Pn、Gs和Ls分别下降了44.61%、50.03%和18.51%,差异均达到显著水平,Ci上升了3.76%,但差异未达到显著水平。
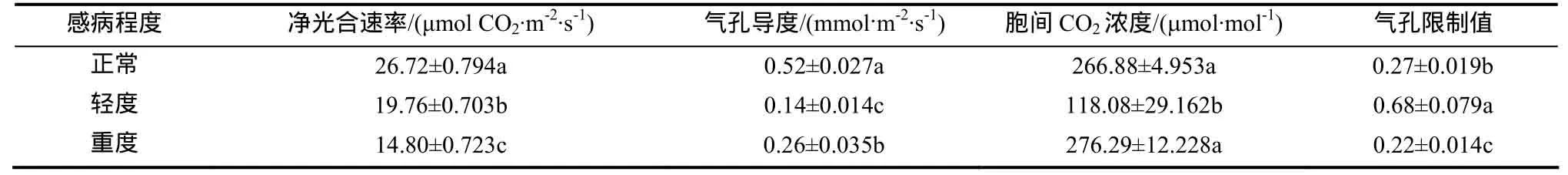
表1 TMV侵染对烟草叶片净光合速率,气孔导度,胞间CO2浓度和气孔限制值的影响Table 1 Effects of TMV infection on net photosynthetic rate, stomatal conductance, intercellular CO2 concentration and stomatal limitation value of tobacco leaves
2.2 TMV侵染对烟草叶片光合色素含量的影响
从表2可见,受TMV侵染的叶中叶绿素和类胡萝卜素含量均有下降。与正常株相比,轻度感病株叶片叶绿素a、叶绿素b、叶绿素a+b和类胡萝卜素分别下降了 16.28%、17.18%、16.55%和16.89%,除类胡萝卜素外,其他均未达到显著水平;重度感病株分别下降了 56.13%、58.69%、56.83%和57.03%,均达到显著水平。不同感病程度烟株叶片叶绿素a/叶绿素b和叶绿素/类胡萝卜素的比值与正常株接近,变化不大。

表2 TMV侵染对烟草叶片光合色素含量的影响Table 2 Effects of TMV infection on photosynthetic pigment content of tobacco leaves
2.3 TMV侵染对烟草叶片叶绿素荧光参数的影响
从表3可见,受TMV侵染的叶片各荧光参数均有下降。与正常烟株相比,轻度感病株叶片Fv/Fm、Fv/F0、qp、NPQ和ΦPSⅡ分别下降了1.22%、6.40%、4.60%、31.03%和4.55%,除NPQ外,其他均未达到显著水平;重度感病株叶片分别下降了17.07%、50.90%、32.18%、75.86%和 40.91%,均达到显著水平。
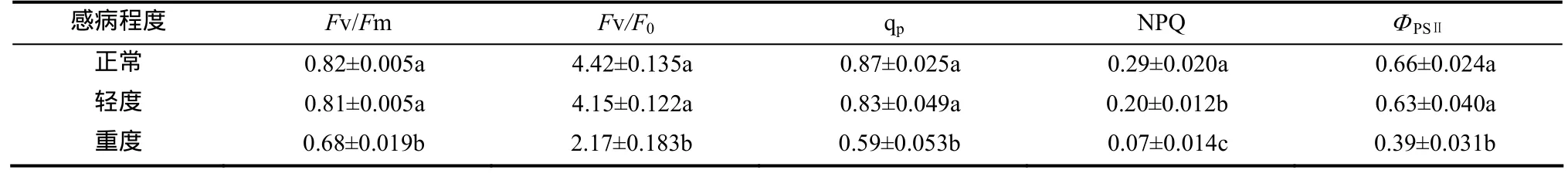
表3 TMV侵染对烟草叶片叶绿素荧光参数的影响Table 3 Effects of TMV infection on chlorophyll fluorescence parameters of tobacco leaves
3 讨 论
通过对受TMV侵染的烟草叶片光合气体交换参数、光合色素以及叶绿素荧光参数变化分析,可以看出TMV对寄主植物光合作用的影响与其他病毒相似。本研究中受TMV侵染叶片叶绿素和类胡萝卜素含量均有所下降,其主要原因可能是病毒侵染寄主植物后使其叶绿素分解酶的活性增加,或使叶绿素酯的含量降低,从而导致叶绿素含量下降而产生浅绿和暗绿相间的花叶症状,致使光合作用面积减少[9];也有可能是 TMV侵染后破坏了叶绿体结构,从而导致叶绿素含量的下降[6];类胡萝卜素既是光合色素,又是内源抗氧化剂,除在光合作用中具有一定的功能外,在细胞内还可吸收剩余能,猝灭活性氧,从而防止膜脂过氧化[10]。由于类胡萝卜素含量的降低,减少其对活性氧的猝灭,导致细胞内积累较多的氧自由基,破坏了叶绿体膜结构,加速了叶绿素的分解[11]。
叶绿素荧光经常被用于评价光合机构的功能和环境胁迫对其的影响[12],本实验测定结果表明,TMV重度侵染可以明显降低Fv/F0、Fv/Fm和ΦPSⅡ,同时qp和NPQ也明显表现出下降的趋势。Fv/F0代表PSⅡ活性[13],其值的降低表明受TMV侵染烟草叶片PSⅡ活性失活,而在TMV侵染下烟草叶片叶绿素含量下降较快和ΦPSⅡ降幅较高的结果表明,光系统失活的可能原因是 PSⅡ天线色素或色素蛋白复合体受到破坏[14];Fv/Fm的下降表明,受TMV侵染烟草叶片的光合机构受到一定程度的破坏,因而,对光能的吸收和转化效率明显下降[15];qp反映的是 PSⅡ天线色素吸收的光能用于光化学传递的份额,要保持高的光学猝灭就要使PSⅡ反应中心处于开放状态,所以qp也反映了 PSⅡ反应中心开放程度,qp值的降低表明PSⅡ反应中心受体QA的还原程度更高,开放部分比例下降,关闭部分比例提高,这些关闭状态的PSⅡ反应中心不能进行稳定的电荷分离,因而不能参与光合电子的传递[9];NPQ代表PSⅡ天线色素吸收的光能不能用于光合电子传递而以热能的形式耗散部分,Wilhelmova等[5]认为受TMV侵染的烟草叶片的NPQ值是上升的,而本实验结果表明,随着TMV侵染程度的加重,NPQ值逐渐降低,下降的可能原因是在热耗散和清除活性氧方面起重要作用的类胡萝卜素含量大幅下降,导致光合机构易受到破坏,耗散过剩光能的能力减弱。
引起叶片光合速率降低的植物自身因素不外乎气孔的部分关闭和叶肉细胞光合活性下降两类,前者使胞间CO2浓度(Ci)降低,而后者使Ci增高。当两种因素同时存在时,变化的方向依赖于占优势的那个因素。也就是说,当气孔的部分关闭是叶片光合速率降低的主要原因时,Ci降低;而当叶肉细胞光合活性降低是叶片光合速率降低的主要原因时,Ci增高[16]。根据Farquhar和Sharkey[17]提出的气孔限制值分析观点,判定引起叶片光合速率降低主要因素的主要根据是Ci和气孔限制值(Ls)的变化方向,Ci降低和Ls升高表明气孔导度降低是主要原因,而Ci增高和Ls降低则表明引起光合速率降低的主要原因是非气孔因素。从本研究结果来看,受 TMV侵染的烟草叶片的净光合速率(Pn)明显下降,且随着感病程度的增加,Pn下降的程度增大,但不同感病程度引起Pn下降的主要原因不同。在轻度感病条件下,伴随着气孔导度(Gs)的下降,Ci降低和Ls升高,表明引起Pn下降的主要原因是气孔限制。在重度感病条件下,伴随着气孔导度(Gs)的下降,Ci增高和Ls降低,表明引起Pn下降的主要原因是非气孔限制及叶肉细胞光合活性的下降。
由此可以推测,TMV轻度侵染主要是引起烟草叶片气孔关闭或堵塞,阻碍了气体的交换,从而影响到叶片的光合作用,随着侵染程度的加重,光合色素含量大幅减少,光合机构损伤严重,叶片光合作用进一步受到影响。
[1]Culver J N, Lindbeck A G C, Dawson W O.Virus-host interactions:induction of chlorotic and necrotic responses in plants by tobacco mosaic virus[J].Annu Rev Plant Pathol., 1991, 29: 193-217.
[2]郭兴启,温孚江,朱汉城.烟草感染马铃薯 Y病毒(PVY)后光合作用的变化规律[J].浙江大学学报:农业与生命科学版,2000,26(1):75-78.
[3]王春梅,施定基,朱水芳, 等.黄瓜花叶病毒对烟草叶片和叶绿体光合活性的影响[J].植物学报,2000,42(4):388-392.
[4]何金祥,黄宁珍,付传明,等.烟草花叶病毒对烟草叶片光合特征和 POD表达的影响[J].西北植物学报,2009,29(5):0961-0967.
[5]Wilhelmová N, Procházková D, Šindelárová M, et al.Photosynthesis in leaves of Nicotiana tabacum L.infected with tobacco mosaic virus[J].Photosynthetica,2005, 43(4): 597-602.
[6]刘国顺,张彩霞,王婵娟,等.受烤烟花叶病病毒侵染后叶中保护酶活性和超微结构的变化[J].植物生理学通讯,2009,45(5):459-463.
[7]马新明,李春明,田志强,等.镉污染对烤烟光合特性、产量及其品质的影响[J].生态学报,2006,26(12):4039-4044.
[8]李合生.植物生理生化实验原理与技术[M].北京:高等教育出版社,2000:134-137.
[9]李燕宏,洪健,谢礼,等.蚕豆萎蔫病毒2号分离物侵染对蚕豆叶片光合活性和叶绿体超微结构的影响[J].植物生理与分子生物学学报,2006,32(4):490-496.
[10]王大平,曾明,朱均,等.绿斑病藻寄生对夏橙叶片光合作用特性的影响[J].应用生态学报,2006,17(6):1141-1144.
[11]Sakaki T, Kondo N, Sugahra K.Breakdown of photosynthetic pigment and lipids in spinach leaves ozone fumigation:Role of active oxygen[J].Physiol Plant,1983, 59: 28-34.
[12]Lichtenthaler H K, Buschmann C, Rinderlel V, et al.Application of chlorophyll fluorescence in ecophysiology[J].Radiation and Environmental Biophysics, 1986, 25: 297-308.
[13]张其德,刘合芹,张建华,等.限水灌溉对冬小麦旗叶某些光合特性的影响[J].作物学报,2000,26(6):869-873.
[14]Saitanis C J, Riga-Karandinos A N, Karandinos M G.Effects of ozone on chlorophyll and quantum yield of tobacco(Nicotiana tabacum L.) varieties[J].Chemosphere, 2001, 42: 945-953.
[15]张乃华,高辉远,邹琦.Ca2+缓解 NaCl胁迫引起的玉米光合作用下降的作用[J].植物生态学报,2005,29(2):324-330.
[16]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[17]Farqhar G D, Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant physiology,1982, 33: 317-345.
猜你喜欢
科学(2022年4期)2022-10-25 02:43:00
当代水产(2022年1期)2022-04-26 14:35:30
新农民(2020年22期)2020-08-31 01:32:08
科学咨询(2020年10期)2020-04-01 03:43:16
乡村科技(2019年25期)2019-02-21 07:40:48
Coco薇(2016年5期)2016-06-03 09:17:41
西南农业学报(2016年5期)2016-05-17 05:42:33
西南农业学报(2016年6期)2016-04-16 05:12:51
考试周刊(2016年6期)2016-03-11 08:14:32
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:45:31
