
5个热带禾本科牧草品种的光合-光响应特性研究
2010-07-25 08:52黄玉清桂堂辉黎彦余
草业科学 2010年9期
莫 凌,黄玉清,桂堂辉,2,黎彦余,2
(1.广西壮族自治区中国科学院广西植物研究所,广西桂林 541006;2.广西师范大学生命科学学院,广西桂林 541004)
广西属于营养体农业发展的适宜地区,植物生长旺盛,草类产量较高。全区可利用草山草坡面积646.67万hm2[1],但其牧草与温带草原牧草相比,适口性差,草场载畜力低,牧业发展潜力有限,且很多天然草地出现了不同程度的退化[2]。随着广西畜牧业的飞速发展,发展人工草地是解决牧草资源的有效途径,对优质饲草饲料的需求也与日俱增。通过引种优良牧草,充分发挥区域资源优势,寻求能适应广西环境、生长快且生物产量高的牧草品种,扩大人工牧草种植面积将成为广西急需解决的问题,也是广西畜牧业稳定发展的重要保障。而推广种植优良禾本科牧草是发展草地畜牧业的关键,选择与利用不同种类的优良牧草对改良天然草地,建立栽培草地及各类饲料生产基地均十分重要[3]。
桂牧一号杂交象草(Pennisetum purpureumcv.Mott)×[(P.americanum×P.purureum)cv.Guimu No.1]是广西畜牧研究所采用从美国引进的杂交狼尾草[(P.americanum)×象草(P.purpureum)]为母本,矮象草(P.purpureum)为父本进行杂交选育而成的一种新型牧草,已在桂西北喀斯特山区广泛引种栽培[4],是我国南方地区的主要多年生当家草种之一。杂交狼尾草(P.americanum×P.purpureum)是20世纪90年代初引进和筛选出的具有发展潜力的热带牧草品种,韦兰英等[5]研究表明,在岩溶石漠化地区,杂交狼尾草无论是单播还是混播都能保持较高的生长速度和极强的适应能力。黑籽雀稗“热研11号”(Paspalum atratumcv.Reyan No.11)是中国热带农业科学院从印度尼西亚引进的热带牧草,适宜在湿热地区中等肥力或贫瘠土壤种植,并有一定的耐寒和耐旱性[6],是中国热带、亚热带地区优良的禾本科牧草[7],在低纬度热带和亚热带一些光照充足地带生长良好[8]。MulatoⅡ杂交臂形草(Brachiaria ruziziensis×B.decumbens×B.brizantha)和乌邦雀稗(P.atratumUbon)是著名的热带牧草育种科学家泰国乌汶大学新西兰籍教授Hare博士与CIAT合作,最近3年推出的新品种,MulatoⅡ杂交臂形草具有抗病虫害强、产量高、品质好的特点。乌邦雀稗原产于巴西,引进泰国后由 Ubon Ratchathani大学筛选并进行种子繁殖研究,然后在泰国得名,该种产量高,适口性好。这2种禾本科热带牧草抗湿和抗旱性能均好,具有很高的叶茎比[9-11]。
光合作用是植物将太阳能转换为化学能的过程,是植物体内最重要的化学反应。光是影响植物生长发育和生存的重要环境因子之一。植物适应光环境变化的能力很大程度上决定了它的分布模式和物种丰度[12]。本研究对以上优良牧草品种的光合、蒸腾及水分利用效率等生理特征对光照强度的响应进行测定,探讨光照强度对植物生长发育的影响,为广西开发利用优良牧草品种提供理论依据。
1 材料与方法
1.1 研究地自然概况试验地设在广西桂林中国科学院广西植物研究所苗圃地,地理位置为110°12′E,25°11′N,海拔 170 m 。年平均温度19.2℃,最热月(7月)平均温度28.3℃,最冷月(1月)平均温度8.4℃,极端最高温38℃,极端最低温-6℃,冬有霜冻,偶见雪。年降水量1 655.6 mm,降水集中在4-6月,秋冬季水量较少,干湿交替明显,年平均相对湿度78%,地势平坦、开阔,光照、通风条件良好[13]。试验地土壤全氮含量1.41 g/kg,全磷含量0.97 g/kg,全钾含量8.48 g/kg。
1.2 试验材料参试牧草分别为:杂交臂形草MulatoⅡ、桂牧一号杂交象草、杂交狼尾草、乌邦雀稗、黑籽雀稗。其中杂交臂形草和乌邦雀稗引自泰国,其余3个种引自广西壮族自治区草业中心。2008年5月中旬播种,种植密度适宜,小区面积为2 m×3 m,随机区组设计,重复3次,田间肥水管理一致。
1.3 叶片光响应曲线测定试验于2009年8月下旬进行,5种牧草均处于营养生长期,且长势良好。采用Li-6400光合测定仪,选取自然生长条件下健康植株的正常叶片,每种植物测量4株,每株选取1个叶片,分别挂牌标记,测定植株相同叶位(旗叶最宽部位),测定时保持叶片自然生长角度不变。将叶片放入叶室内,启动人工光源,将光强设定为 2 200、1 800、1 500、1 200、1 000、800、600 、400、200 、150、 100 、50 、20 和 0 μ mol/(m2·s),温度控制为33℃、CO2浓度和空气湿度均为自然状态,测量指标有叶片净光合速率[Pn,μ mol/(m2·s)]、蒸腾速率[Tr,mmol/(m2·s)]、气孔导度[Gs,mol/(m2·s)]、胞间 CO2浓度(Ci,μ mol/mol)、以Pn/Tr计算瞬时水分利用效率(WUE),结果取平均值。
1.4 数据分析本研究用Excel进行数据处理,用SPSS13.0软件拟合光响应曲线及对数据进行相关分析与One-Way ANOVA方差分析,并采用LSD法进行多重比较。根据Walker[14]的方法进行计算和拟合。
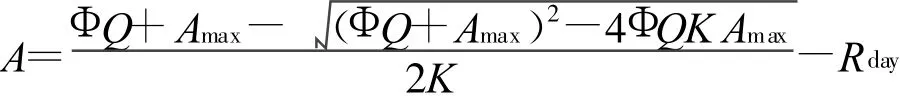
式中,A为净光合速率,Amax为最大净光合速率,Φ为表观量子效率,K为曲角,Rday为光下呼吸速率,Q为光合有效辐射。
将 200 μ mol/(m2· s)以下的 Pn-PAR 响应曲线进行直线回归,其斜率即为表观量子效率。利用该直线方程与计算得出来的最大净光合速率与x轴平行的这两条直线相连,得出交点,其x轴的数值即为光饱和点(LSP)。利用植物叶片在200 μ mol/(m2·s)以下的光强时的光曲线符合直线方程来计算光补偿点(LCP)。
2 结果与分析
2.15个禾本科牧草品种的光合-光响应参数在一定环境条件下,叶片的最大光合速率表示了叶片的最大光合能力[15],表观量子利用效率反映了叶片对光能的利用情况,尤其是对弱光的利用能力[16]。光饱和点与光补偿点反映了植物对光照条件的要求,光补偿点较低、光饱和点较高的植物对光环境的适应性较强;而光补偿点较高、光饱和点较低的植物对光照的适应性较弱[17]。通过非直角双曲线拟合计算出5个牧草品种的叶片最大光合能力、光下呼吸速率、表观量子效率、光饱合点与光补偿点(表1)。5个牧草品种中,3个杂交品种的Amax、LSP均显著高于(P<0.05)2个雀稗品种,杂交臂形草的Amax、Φ均显著高于杂交狼尾草和桂牧一号,具有较高的光饱和点,同时又有较高的最大光合速率,而LCP则显著小于桂牧一号,表明其利用弱光的能力比桂牧一号要强,Rday、LSP则无显著差异,说明对当地环境具有很好的适应性。杂交狼尾草和桂牧一号各项指标均无显著差异(P>0.05)。乌邦雀稗和黑籽雀稗的各项指标均无显著差异,但乌邦雀稗的Amax、Rday、LSP比黑籽雀稗稍大,Φ、LCP稍小,说明乌邦雀稗利用光的能力比黑籽雀稗稍强一些。

表1 5个热带牧草品种的光响应曲线参数
2.2 光合速率和蒸腾速率光合作用是植物物质生产的基础,光合速率的高低决定了光合能力的强弱,是植物生物产量形成的关键。在所给光合有效辐射(PAR)的范围内5个牧草品种的光合速率均随光合有效辐射的增强而增加,而后趋于变缓,即单位数量PAR的增加对植物Pn的增大贡献随PAR的增大而减小。而随光合有效辐射的增加,3个杂交牧草品种的Pn增大幅度比2个雀稗品种要大,2个雀稗品种到达光饱合点的光合有效辐射要比3个杂交牧草品种低。在光合有效辐射超过 800 μ mol/(m2·s)之后,杂交种与雀稗种的光响应曲线差异渐渐增大。而5个牧草品种的蒸腾速率与光合速率相似,均随光合有效辐射的增强而增加,以杂交臂形草的蒸腾速率最高,达6.44 mmol/(m2·s),其次为杂交狼尾草和桂牧一号,分别为 5.42、5.04 mol/(m2·s),2个雀稗品种相近,均在3.70 mol/(m2·s)左右(图1)。
2.3 气孔导度与水分利用效率气孔是植物叶片与外界进行气体交换的主要通道。植物在光下进行光合作用,经由气孔吸收CO2和发生蒸腾作用,气孔可以根据环境条件的变化来调节开度的大小而使植物在损失水分较少的条件下获取最多的CO2。5个牧草品种中,以杂交臂形草的气孔导度最大(图1),可达0.443 mol/(m2·s),其次是杂交狼尾草和桂牧一号,最后是2个雀稗品种,只有0.25、0.26 mol/(m2·s)。
随着光照强度的增加,5个牧草品种的同化作用伴随着水分的散失均呈迅速上升的趋势,在光合有效辐射超过600 μ mol/(m2·s)时,光合速率上升的趋势减缓(图1),而蒸腾速率仍保持相对较高的上升趋势,因此各叶片的水分利用效率迅速达到饱和,并且随着光照的进一步加强而呈现逐渐下降趋势。结果表明,5个牧草品种的水分利用效率相近,以2个雀稗品种的水分利用效率稍低,在较强的光照条件下随着光照的加强,水分利用能力有所下降。
3 讨论与结论
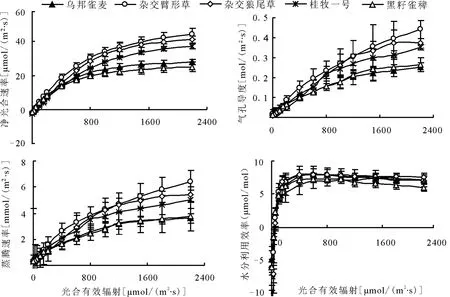
图1 5个牧草品种净光合速率、蒸腾速率、气孔导度、水分利用效率对不同光合有效辐射的响应
有效光合辐射是植物光合作用的能量来源,与光合过程的各生理特性密切相关[18]。表观量子效率和光补偿点是植物利用弱光能力的一项重要指标。表观量子效率越大,表明植物吸收与转换光能的色素蛋白复合体可能较多,利用弱光的能力越强[19]。研究结果表明,5个牧草品种的表观量子效率为0.044~0.057,光补偿点均在100 μ mol/(m2·s)以下,最大的桂牧一号也只有 55.8 μ mol/(m2·s),其次是杂交狼尾草,为 47.8 μ mol/(m2·s),最小的乌邦雀稗只有30.1 μ mol/(m2·s)。可见这5个牧草品种对弱光的利用率均较强,在低光强的环境下也有较好地生长。光饱和点则可以揭示植物对强光的喜好程度,光饱和点越高的植物越能有效地利用全日照的强光,3个杂交品种的光饱和点都较高,均在1 000 μ mol/(m2·s)以上,而 2 个雀稗品种的光饱和点则较低,分别为 702.1和605.6 μ mol/(m2·s),说明3个杂交品种对全日照的强光利用能力较2个雀稗品种要高,这可能是种间差异造成。叶片的最大净光合速率表示了植物的最大光合作用能力,光响应曲线反映了5个供试材料的光合能力的大小,即杂交臂形草的光合能力>杂交狼尾草>桂牧一号>乌邦雀稗>黑籽雀稗。
研究结果表明,在所给光条件下,5个牧草品种的光合速率并未出现下降现象,表明均没有光抑制现象发生,在与最强自然光强相似的光强作用情况下,5个牧草品种仍然能够维持较高的光合速率[大于20 μ mol/(m2·s)],表明这 5个牧草品种均具有很强的光合能力和对极端高光的适应能力,这一结果与温达志等[20]的研究结果相似。杂交臂形草的光合速率最高,光适应范围最宽,利用弱光的能力及最大光合能力均强于桂牧一号和杂交狼尾草,而2个雀稗品种最低,光适应范围近似,乌邦雀稗利用光的能力则较黑籽雀稗稍高。5个牧草品种在PAR为 400~2 200 μ mol/(m2·s)内 WUE 均可维持在 6~8 μ mol/(m2·s),表明在桂林地区5个热带牧草品种光合特性方面均存在一定的竞争优势。
[1]廖维和.从广西区情看草业在国民经济中的地位和作用[J].草业科学,1999,16(2):49-54.
[2]蒋建生.广西草地资源开发利用策略初探[J].草业科学,1996,13(1):70-72.
[3]姚庆群,白昌军,王文强,等.22份禾本科牧草种质适应性评价[J].草业科学,2009,26(6):84-89.
[4]肖润林,单武雄,方宝华.喀斯特峰丛洼地桂牧一号杂交象草对不同追施氮肥水平的响应[J].生态学杂志,2008,27(5):735-739.
[5]韦兰英,袁维圆,尤业明,等.岩溶石漠化区牧草植物地上部分生物量的动态变化[J].草业科学,2009,26(10):73-79.
[6]侯海军,郭建春,胡新文.“热研11号”黑籽雀稗植株再生体系的建立[J].热带作物学报,2008,29(2):202-205.
[7]王文强,付玲玲,白昌军.热研 11号黑籽雀稗开花生物学特性[J].中国农学通报,2007,23(8):495-498.
[8]刘金祥,陈伟云,肖生鸿.黑籽雀稗的光合生理特性研究[J].草业学报,2009,18(6):254-258.
[9]Hare M D,Saengkham M,Kaewkunya C,et al.Effect of cutting on yield and quality ofPaspalum atratumin Thailand[J].Tropical Grasslands,2001,35:144-150.
[10]Hare M D,Kaewkunya C,Tatsapong P,et al.Method and time of establishingPaspalum atrtumseed crops in Thailand[J].Tropical Grasslands,2001,35:19-25.
[11]Hare M D,Tatsapong P,Saipraset K.Seed production of two brachiaria hybrid cultivars in north-east Thailand.1.Method and time of planting[J].Tropical Grasslands,2007,41:26-34.
[12]林栋,吕世海,冯朝阳,等.华北山地阳坡中生灌草植被对CO2浓度和温度变化的光合响应[J].草业科学,2008,25(4):135-140.
[13]莫凌,韦兰英,张中峰,等.桂北中华猕猴桃光合蒸腾特性及其影响因子研究[J].西南农业学报,2008,21(4):968-971.
[14]Walker D A.Automated measurement of leaf photosynthetic O2evolution as a function of photon flux density[J].Philosophical Transactions of the Royal Society of London:Series B,Biological Science,1989,323(1216):313-326.
[15]陆佩玲,罗毅,刘建栋,等.华北地区冬小麦光合作用的光响应曲线的特征参数[J].应用气象学报,2000,11(2):236-241.
[16]蹇洪英,邹寿青.地毯草的光合特性研究[J].广西植物,2003,23(2):181-184.
[17]杨兴洪,邹琦,赵世杰.遮荫和全光生长的棉花光合作用和叶绿素荧光特征[J].植物生态学报,2005,29(1):8-15.
[18]孙伟,王德利,王立,等.狗尾草蒸腾特性与水分利用效率对模拟光辐射增强和CO2浓度升高的响应[J].植物生态学报,2003,27(4):448-453.
[19]吴承祯,侯智勇,洪伟,等.桉树无性系光合光响应研究[J].福建林学院学报,2008,28(3):198-202.
[20]温达志,张德强,周国逸,等.四种禾本科牧草植物光合特性的初步研究[J].热带亚热带植物学报,2000(增刊Ⅲ):59-66.
猜你喜欢
中国饲料(2022年5期)2022-04-26
今日农业(2021年10期)2021-07-28
江苏农业科学(2020年12期)2020-07-20
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20
广东饲料(2016年3期)2016-12-01
西藏科技(2015年11期)2015-09-26
新课程·中旬(2014年10期)2014-12-26
作物研究(2014年6期)2014-03-01
当代畜禽养殖业(2014年1期)2014-02-27

- 草业科学的其它文章
- 世界信息
- 甜高粱在青海高原种植的初步研究