
嵩草草甸退化和恢复过程中主要牧草演替和地表特征变化
2010-06-08 07:52李以康林丽张法伟梁东营王溪曹广民
草业学报 2010年5期
李以康,林丽,张法伟,梁东营,2,王溪,曹广民*
(1.中国科学院西北高原生物研究所,青海 西宁810008;2.中国科学院研究生院,北京100039)
草原退化已成为我国重大的资源和环境问题之一[1,2]。青藏高原由于特殊的地理环境和恶劣的环境条件,生态系统脆弱、抗干扰能力差,草甸退化尤为突出[3,4]。通常认为放牧干扰是造成草甸退化的直接推动力[5-7],放牧活动导致植被演替,使生态系统的物种组成发生变化,植物种群的优势地位发生更替[6,8-10],从而产生不同的草甸类型。放牧牲畜通过取食和践踏等行为,减弱了优势种的竞争能力,导致了更多小尺度上的空间异质性[11]。重牧下的牲畜践踏行为损伤牧草、减少凋落物的现存量,增加土壤的紧实度、容重,降低土壤孔隙度、水稳性团聚体、透水性和透气性,导致雨后水涝和植物根区缺氧[12]。经过长期放牧的高寒草甸,在草甸的表面会形成不同大小和数目的黑斑和秃斑,严重的甚至表面还产生了菌斑,阻碍了物种的定殖和繁衍。草甸表面日益扩大的黑斑加速了退化。而小嵩草草甸发生塌陷和裂缝的地段,则成为物种入侵和繁衍的空斑,空斑既为草地植被更新提供了主要场所,又创造了维持草地物种多样性的重要环境[13]。
放牧利用下,天然草地几乎无时不处于演替之中,斑块的演替,特别是微斑块的变化更为活跃[14]。野外观察发现,禾草、矮嵩草(Kobresiahumilis)和小嵩草(K.pygmaea)主要依靠营养繁殖,都呈现出聚集分布的格局,往往在草甸中呈斑块状分布,而聚集分布也是种群抵抗不良环境的生态适应对策[15]。辛晓平等[16]的研究表明不同物种斑块空间格局动态不同,在恢复演替各个阶段中的地位与作用也不同;这些斑块的大小和数量的多少往往体现了演替和退化程度。Alados等[17]研究认为物种空间分布异质性的增加是组成物种自组织能力的结果,以前的研究多是以大尺度斑块的植被组成和物种变化为对象[18-20],对演替的格局和过程等进行分析[21],很少涉及到单个克隆植物的微斑块数量大小在放牧条件下的演替变化[14]。本研究以高寒草甸植被在退化到小嵩草草甸和采取封育恢复措施后的主要植被斑块变化,结合地表黑斑和秃斑等的变化为研究内容,力图揭示它们更替和变化的过程,使对高寒草甸在退化和恢复过程中的变化有一个更具体详细的认识。
1 材料与方法
1.1 样地选择
试验样地设置于青海省果洛州玛沁县和海北州祁连县。样地类型包括退化中的矮嵩草草甸、重牧小嵩草草甸,恢复中的小嵩草草甸和恢复较好的草甸。各样地由于牧民对草甸的放牧强度和人为管理措施的不同而不同,具体情况如下:
玉树县巴塘滩样地:位于山间滩地,植被由矮嵩草群落向小嵩草群落演替的过渡期。草地作为冬春草场进行正常放牧。
海北州祁连县俄博乡重牧样地:由于牧民大量放牧牦牛导致退化,植被盖度低,禾草零星分布。小嵩草为群落优势种。
海北州祁连县俄博乡封育样地:与重牧样地相邻,为牧民减轻放牧压力在原来重牧草甸的基础上自己封育形成,草甸已经得到恢复,与原来重牧的草甸形成鲜明的对比。
玛沁县大武镇样地:设置于山间滩地,封育前退化轻,为典型的小嵩草草甸。通过围栏封育,仅冬春季节进行放牧。设置围栏封育(表1)。

表1 样地基本状况Table 1 The basic instance of plots
1.2 取样方法
地表特征:采用样带法,在2008年4月进行了返青期样方调查。平行设置3条25 m长的样带(带间间隔10 m),每条样带上隔5 m沿线的两侧各设置1个25 cm×25 cm的样方,调查内容包括禾草、矮嵩草、小嵩草、地表黑斑、秃斑、群落总盖度等。斑块的出现几率以出现斑块的样方数和总的样方数的百分比表示;直径约1 cm以上的计为斑块,盖度以这一类斑块在样方中所占的总比例表示。
群落特征:2008年8月采样。采用样方法,随机选取6个面积为25 cm×25 cm样方进行植物群落特征(植物种分盖度以及群落盖度)调查。生物量测定采用收获法,齐地面剪去植物的地上部分,装在信封中带回实验室以80℃的温度烘干至恒重;盖度的测定采用目测法;重要值(IV)=(相对盖度+相对生物量)/2[22]
2 结果与分析
2.1 退化和恢复过程中主要种群重要值变化
野外观察和实验表明,退化中嵩草草甸处于矮嵩草向小嵩草转化阶段,小嵩草斑块占优势面积约占样地60%~70%,矮嵩草斑块占优势的面积约占30%。小嵩草丛径在3~35 cm之间,矮嵩草丛径在5~75 cm之间,大于小嵩草丛径。小嵩草重要值显著高于矮嵩草和禾草(P<0.05),高出矮嵩草48%,更远高于禾草重要值(表2);禾草生长受到限制后,矮嵩草作为牲畜喜食牧草,生长也受到极大影响。以禾草为优势种的嵩草草甸,由于放牧牲畜的选择性啃食作用,禾草生长和繁殖都受到极大限制,导致禾草种类减少,植株低矮,生物量和植被盖度降低,重要值远低于小嵩草重要值。
在重牧小嵩草草甸,矮嵩草斑块基本消失;小嵩草成为绝对优势种(重要值39%),远高于矮嵩草和禾草;禾草分布少,重要值低,在样地中只是零散分布,但在草甸塌陷处和裂缝里分布较多,这与塌陷和裂缝所形成的空斑和土质松软的条件有关(表2)。
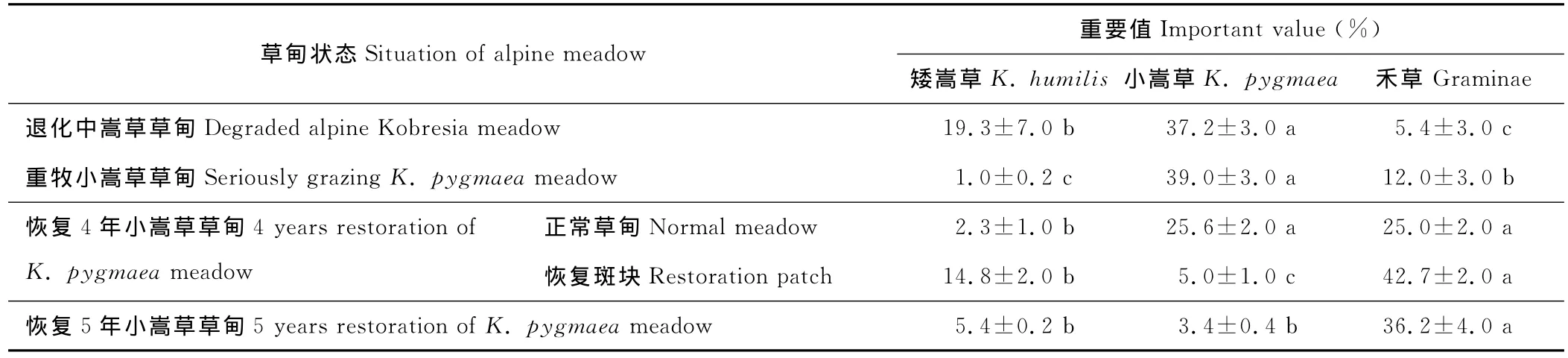
表2 嵩草草甸退化和恢复过程中小嵩草、矮嵩草和禾草之间重要值的比较Table 2 Comparative of K.pygmaea,K.humilis and Gramineae important value in degradation and restoration of alpine Kobresia meadow
对退化样地采取封育措施可以使草甸得到恢复。恢复4年小嵩草草甸是过去发生过草皮层塌陷的草地,塌陷坑面积占地面45%。封育后老化草皮上植被发生逆转,但小嵩草仍为建群种,剥蚀塌陷处以禾草为建群种,地面黑斑基本不见。小嵩草重要值比退化中嵩草草甸降低了31.2%,禾草生长得到很大恢复;矮嵩草恢复相对较慢,重要值较低。恢复较好的恢复斑块和样地中正常恢复的部分相比较,小嵩草斑块基本消失,矮嵩草生长得到极大恢复,禾草生长得到进一步加强,成为占据绝对优势的优势种,斑块恢复成为以禾草和矮嵩草为优势种的群落。
对退化不严重的小嵩草草甸进行5年的围栏封育后,与围栏外相比,小嵩草草甸从以小嵩草为优势种恢复成以禾草为优势种。莎草科植物的重要值仅为禾草的近一半,显著低于禾草的重要值(P<0.05),矮嵩草和小嵩草的斑块面积减小,分布也少。
从嵩草草甸退化到重牧小嵩草草甸,再恢复成为禾草嵩草草甸,禾草和矮嵩草的重要值呈现先降低后升高的趋势,而小嵩草的重要值则相反,先升高后降低(表2)。
2.2 退化和恢复过程中地表特征变化
草甸退化导致优势种改变、植被盖度变化、鼠类入侵、黑斑和秃斑增多等地表特征改变,这些改变一定程度上表征了不同退化阶段高寒草甸的外观特征。
嵩草草甸的退化过程中,在矮嵩草和小嵩草占优势的地方分别各选3条样带,比较小嵩草和矮嵩草样带结果(表3)。从矮嵩草占优势的样带退化到小嵩草占优势的样带,地表黑斑面积扩大近1倍,枯草盖度少了2/3,草甸植被盖度降低;2个样带小嵩草面积相差不大,但是退化导致矮嵩草的面积缩少了近3/4。矮嵩草斑块面积相对较大,但都出现中间部分植株死亡,长势渐衰。杂类草的盖度基本变化不大。退化使小嵩草斑块在样地中出现的几率增加(增加了13.3%),分布更加普遍和均匀,而矮嵩草斑块的出现几率缩小了2/3,显示出了强烈的退化趋势。菌斑开始在小嵩草样带中出现。矮嵩草草甸的退化已经表现出了小嵩草斑块增多的特点。
重牧小嵩草草甸以放牧牦牛为主,牦牛的践踏和啃食使草甸退化较重,地表黑斑遍布,菌斑面积在10%左右,地表塌陷多,濒临退化到杂类草-黑土滩阶段。小嵩草为优势植物,小嵩草斑块分布更加普遍(出现几率100%,表4);禾草和矮嵩草在放牧压力下盖度很低,样地植被盖度变小。
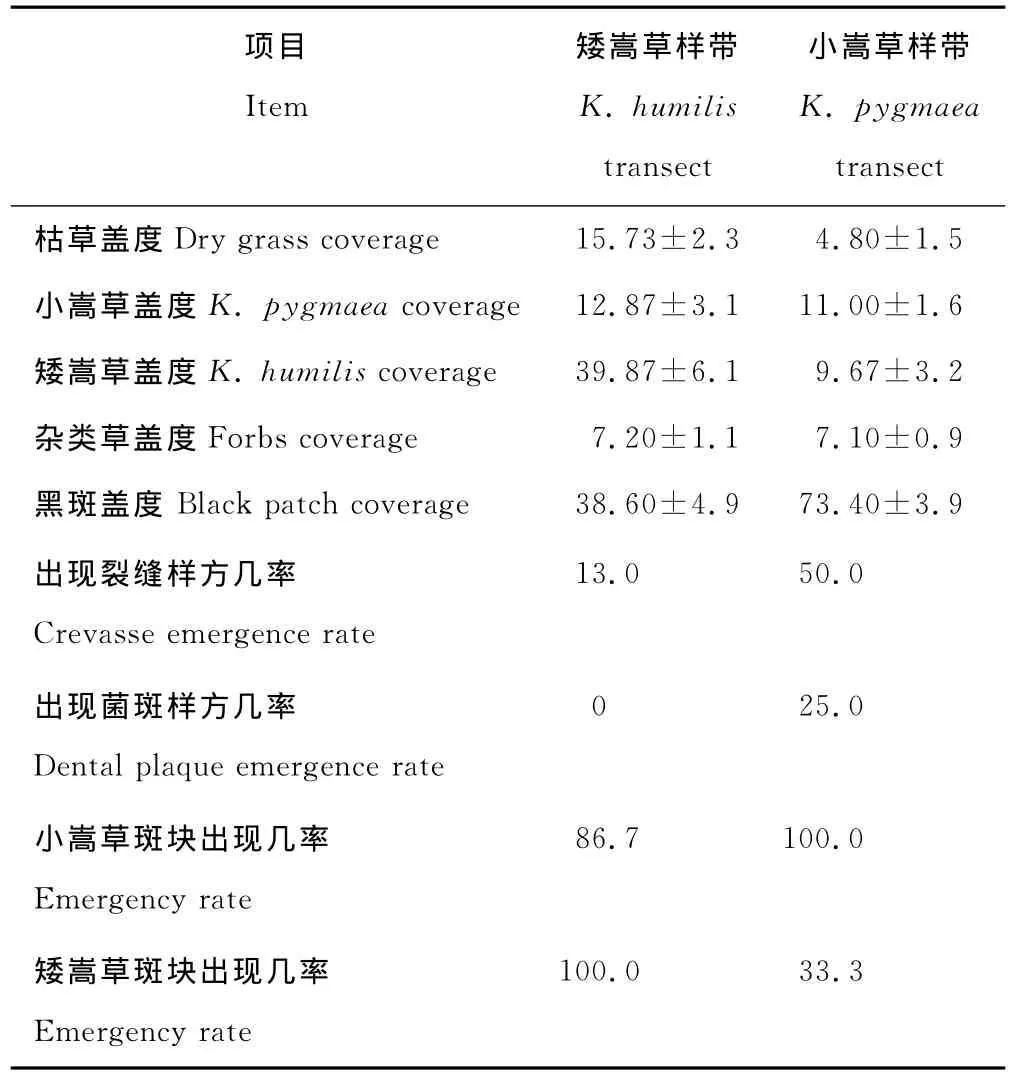
表3 嵩草草甸退化过程中小嵩草和矮嵩草斑块的地表特征Table 3 The soil surface characteristic of K.pygmaeaand K.humilis patch in the course of alpine Kobresia meadow degradation %
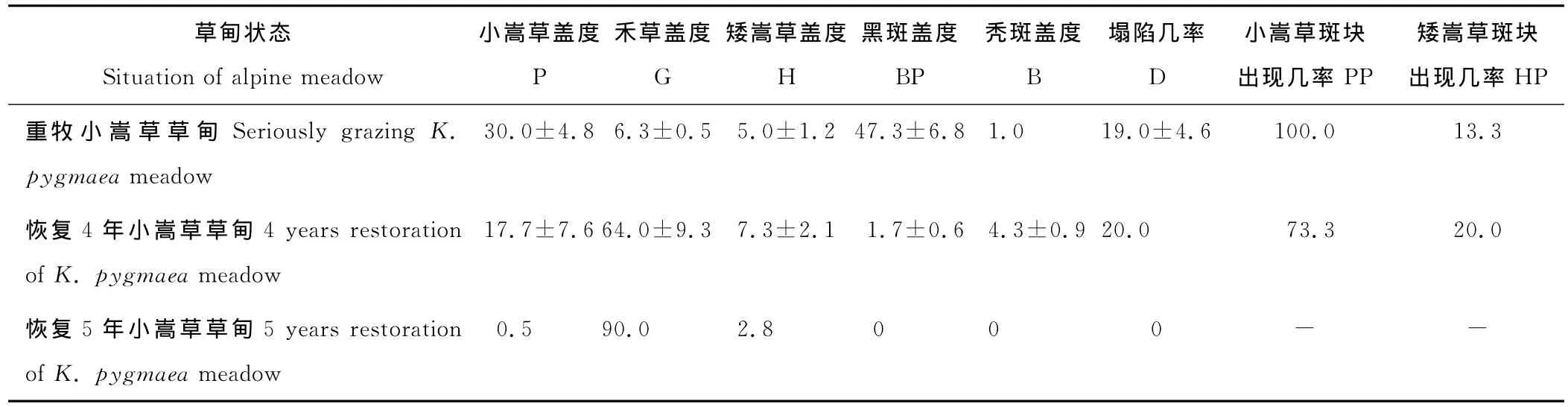
表4 重牧和不同恢复年限小嵩草草甸地表特征Table 4 The soil surface characteristic in seriously grazing and different restoration years of alpine K.pygmaea meadow %
对恢复4年小嵩草草甸返青期的样带调查结果表明(表4),禾草盖度远高于小嵩草和矮嵩草,地面植被得到了较好恢复,黑斑和秃斑的面积变小,小嵩草斑块在样方中出现的几率降低(相比较于退化中的小嵩草草甸,表3),矮嵩草斑块退化后还没有得到很好的恢复,斑块出现的几率不大。退化严重的草甸恢复需要的时间相对要长一些。
对恢复5年小嵩草草甸8月的样地调查表明:恢复样地植被总盖度为100%,地面较平整,没有黑斑和秃斑出现;优势种为垂穗披碱草,甘肃马先蒿(Pediculariskansuensis)、矮火绒草(Leontopodiumnanum)等退化草甸中常见的杂类草极少。莎草科的矮嵩草斑块面积为2%~3%,小嵩草斑块面积为0.5%;但是这两种斑块已经很少出现(表4)。禾本科的种类垂穗披碱草盖度在90%左右。经过5年的封育减牧措施,样地由典型的小嵩草草甸恢复成以禾草等植株较高植物为上层和低矮植物为下层两层片层结构的禾草嵩草草甸。
3 讨论
草地群落斑块的产生源于斑块间生境异质性及植物间生物学和生态学特征的差异[14]。植物通过斑块的形式扩展生长范围,获得空间优势,限制其他植物生长。禾草、矮嵩草和小嵩草能够靠根茎营养繁殖进行生长,在草甸上形成成丛斑块状分布的特点。斑块大小、数量受放牧家畜的选择性取食和践踏影响较大,并最终导致了优势植物的兴衰更替[23、24]。通常认为,放牧家畜对草甸植物的选择性啃食是导致牧草(禾本科)消失、植被演替[25]和嵩草类植物成为优势种的主要原因。矮嵩草属地面芽短根茎植物,是资源密集型的克隆植物,主要靠克隆生长进行繁殖和生长,耐放牧践踏;小嵩草属寒冷中生密丛短根茎地下芽植物,在高寒生境中以营养繁殖为主,有性繁殖为辅。禾草等较高优良牧草的生长和繁殖在放牧压力下受到抑制,矮嵩草和小嵩草得到更大的生长空间,斑块增多,斑块面积不断扩大,而在不同的外力作用下斑块面积和比例也处于动态演替之中。所以在不同演替阶段甚至同一阶段的不同地段,放牧压力和草甸鼠类活动的非均一性使矮嵩草和小嵩草斑块呈现出不同的演替过程。
在草地上出现空斑和植物的丛聚分布现象很常见[27]。草地放牧动物的采食、践踏和排出的代谢物都能引起植物的死亡或破坏,从而产生空斑。在放牧样地内空斑大量存在,而且随着放牧强度的增大,空斑数量和面积呈增加趋势[11]。在高寒草甸的退化过程中,退化草地的表层出现了板结皮,黑斑和秃斑的存在限制了土壤的水气交换功能,退化过程中小嵩草植被的发育和特殊的生物学特性(高地下/地上比)[23],使草毡表层死亡,失去弹性,地表出现的裂缝增多,裂缝的宽度和长度都扩大,促进草甸进一步退化,板结的表面限制了其它物种在空斑上定居。物种对放牧干扰的适应性有一定的限度,不同的物种适应能力不同。研究认为干扰可以促进种内集聚分布[26、27],而放牧干扰通过践踏和采食所产生的空斑[28],为物种的母株通过克隆繁殖产生大量克隆分株提供了拓殖空间[11]。矮嵩草和小嵩草具有耐牧性和克隆繁殖的特点,放牧干扰所产生的空斑为矮嵩草和小嵩草的发展创造了条件,成为过度放牧下的优势种。禾草在减轻放牧压力后,在自身的强大竞争优势下,抑制其他植物的生长和扩展,逐渐成为群落的优势植物。小嵩草和矮嵩草由于株高较小生长在群落下层,处于对光资源竞争的不利状态,主要靠克隆生长,在其他植物抑制之下,生长斑块越来越少,生长逐渐退缩。
干扰直接影响着生态系统的结构和功能演替,适度的干扰可以促进生物多样性和生物资源的保护[29]。在青藏高原高寒草甸,过度放牧导致草甸植被的演替顺序为禾草-矮嵩草群落→矮嵩草群落→小嵩草群落。在小嵩草群落阶段,如果放牧压力没有减轻,可能导致土壤立地条件进一步恶化,黑斑和秃斑增多,植被盖度减少,部分地段发生裂缝和塌陷,鼠兔进入,加速破坏草甸,最终形成杂类草“黑土型”退化草地。小嵩草群落成为草甸进一步退化成“黑土滩”的最后一道屏障。而“黑土滩”上建植人工草地的经验表明,建植后5-8年间的人工草地容易出现群落的逆向演替发展[30]。相比较于人工草地的恢复方式,在原有草甸基础上的恢复,由于原有植被种子库的存在和原生植被的部分存在,可能是一种更有效和快速的植被恢复方式。所以在草甸退化到小嵩草草甸时,要注意减轻放牧压力,防止草原状况进一步恶化。超载过牧是草地退化的直接原因,适度放牧是促进高寒草甸植被恢复和可持续利用的关键。
[1]李博.中国北方草地退化及其防治对策[J].中国农业科学,1997,30(6):1-9.
[2]李金花,李镇清,任继周.放牧对草原植物的影响[J].草业学报,2002,11(1):1-3.
[3]董全民,赵新全,马玉寿,等.牦牛放牧率和放牧季节对小嵩草高寒草甸土壤养分的影响[J].生态学杂志,2005,24(7):729-735.
[4]董锁成,周长进,王海英.“三江源”地区主要生态环境问题与对策[J].自然资源学报,2002,17(6):713-720.
[5]李政海,王炜,刘钟龄.退化草原围封恢复过程中草场质量动态的研究[J].内蒙古大学学报(自然科学版),1995,26(3):334-338.
[6]杨利民,李建东,杨允菲.草地群落放牧干扰梯度β多样性研究[J].应用生态学报,1999,10(4):442-446.
[7]柳小妮,孙九林,张德罡,等.东祁连山不同退化阶段高寒草甸群落结构与植物多样性特征研究[J].草业学报,2008,17(4):1-11.
[8]周兴民,王启基,张堰青,等.不同放牧强度下高寒草甸植被演替规律的数量分析[J].植物生态学与地植物学学报,1987,11(4):276-285.
[9]王亚军,魏兴琥,杨萍.超载放牧对那曲地区高山嵩草草甸植被退化的影响[J].兰州大学学报(自然科学版),2005,41(1):32-38.
[10]鲁为华,朱进忠,王东江,等.天山北坡围栏封育条件下伊犁绢蒿幼苗分布格局及数量动态变化规律研究[J].草业学报,2009,18(4):17-26.
[11]刘振国.内蒙古退化草原对不同类型干扰的相应研究[D].北京:中国科学院研究生院,2006.
[12]侯扶江,常生华,于应文,等.放牧家畜的践踏作用研究评述[J].生态学报,2004,24(4):784-789.
[13]刘桂霞,韩建国.草地生态学领域空斑研究进展[J].生态学杂志,2006,25(9):1136-1140.
[14]张卫国,黄文冰,杨振宇.草地微斑块与草地退化关系的研究[J].草业学报,2003,12(3):44-50.
[15]左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[16]辛晓平,徐斌,王秀山,等.碱化草地群落恢复演替空间格局动态分析[J].生态学报,2001,21(6):877-883.
[17]Alados C L,Aich E A,Komac B,etal.Self-organized spatial patterns of vegetation in alpine grasslands[J].Ecological Modeling,2007,201:233-242.
[18]辛晓平,徐斌,单保庆,等.恢复演替中草地斑块动态及尺度转换分析[J].生态学报,2000,20(4):587-593.
[19]祖元刚,马克明,张喜军.植被空间异质性的分形分析方法[J].生态学报,1997,17(3):334-337.
[20]张宏斌,杨桂霞,黄青,等.呼伦贝尔草甸草原景观格局时空演变分析-以海拉尔及周边地区为例[J].草业学报,2009,18(1):134-143.
[21]Wayne P.Sousa.The role of disturbance in natural communities[J].Annual Review of Ecology and Systematics,1984,15:353-391.
[22]任继周.草业科学研究方法[M].北京:中国农业出版社,1998:1-29.
[23]曹广民,杜岩功,梁东营,等.高寒嵩草草甸的被动与主动退化分异特征及其发生机理[J].山地学报,2007,25(6):641-648.
[24]周兴民,赵新全,曹广民,等.中国嵩草草甸[M].北京:科学出版社,2001.
[25]周华坤,赵新全,周立,等.青藏高原高寒草甸的植被退化与土壤退化特征研究[J].草业学报,2005,14(3):31-40.
[26]Skarpe C.Spatial patterns and dynamics of woody vegetation in an arid savanna[J].Journal of Vegetation Science,1991,2:565-572.
[27]Fehmi J S,Laca E A,Rice K J.The effect of small gaps in California annual grassland on above-ground biomass production[J].Grass and Forage Science,2001,56:323-329.
[28]Kleijn D,Steinger T.Contrasting effects of grazing and hay cutting on the spatial and genetic population structure ofVeratrumalbum,an unpalatable,long-lived,clonal plant species[J].Journal of Ecology,2002,90:360-370.
[29]陈利顶,傅伯杰.干扰的类型、特征及其生态学意义[J].生态学报,2000,20(4):581-586.
[30]王长庭,曹广民,王启兰,等.三江源地区不同建植期人工草地植被特征及其与土壤特征的关系[J].应用生态学报,2007,18(11):2426-2431.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
草地学报(2021年7期)2021-08-03
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
活力(2019年21期)2019-04-01
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
河北林业科技(2016年5期)2016-11-08
杂草学报(2015年2期)2016-01-04
