
黑河中游干旱荒漠地面节肢动物群落季节变异规律
2010-06-08 07:52刘继亮李锋瑞刘七军牛瑞雪
草业学报 2010年5期
刘继亮,李锋瑞,刘七军,牛瑞雪
(中国科学院寒区旱区环境与工程研究所 临泽内陆河流域研究站,甘肃 兰州730000)
干旱荒漠约占全球陆地面积的1/4~1/5,是主要的陆地生态系统类型之一[1,2]。在中国,干旱荒漠主要分布在西北地区,约占国土陆地面积的1/3[3]。干旱荒漠的自然环境特征是:气候干旱、降水稀少、植被稀疏、土壤贫瘠、地表水资源匮缺,是我国生态环境最脆弱的区域之一,也是生态环境保护与建设的重点区域。在干旱荒漠生态系统,由于动植物的生长和繁育受到干旱缺水和高温等环境因素的限制,植被的净初级生产力较低,因而通常认为干旱荒漠生态系统的生物群落结构简单、动物种类贫乏。然而,近年来在北美洲的Mojave、Chihuanhuan和Sonoran荒漠、非洲的Namib荒漠以及亚洲的Negev和Arabian荒漠开展的一系列研究结果显示,荒漠生态系统中地面和土壤动物的种类仍然是相当丰富的,几乎囊括了陆地生态系统中的大多数动物类群[1,4-6]。
地面节肢动物是荒漠生态系统的重要组分之一,它与土壤节肢动物和微生物一起参与了荒漠生态系统凋落物的分解和碳氮等关键元素的地球生物化学循环过程[6-9]。此外,地面节肢动物也是荒漠鸟类、蜥蜴和啮齿类动物的捕食对象,在维持荒漠生态系统的生态服务功能、生物多样性和食物网络结构等方面起着十分关键的作用[6,9-12]。因此,深入开展干旱荒漠生态系统地面节肢动物群落结构、数量和功能群多样性的季节变异规律研究,将为进一步开展荒漠生态系统地上与地下生态过程的相互作用机制研究奠定重要的基础。
近年来,尽管国际上对土壤动物的多样性及其在陆地生态系统中的功能已经进行了相对较多的研究[13,14],但是国内这方面的系统性研究工作尚不多见。以往国内土壤动物的研究主要集中在东北寒温带和中温带地区以及华北和华南暖温带、亚热带和热带地区[15,16],而对西北干旱区内陆河流域的研究甚少。因此,以中国第二大内陆河流域——黑河流域中游的干旱荒漠生态系统为研究对象,重点对荒漠生态系统的地面节肢动物群落的个体数量、生物量和营养功能群多样性进行了系统的定位研究,以期解析地面节肢动物群落组成、数量和功能群多样性的季节变异规律。
1 材料与方法
1.1 研究地点
本研究在中国生态系统网络临泽内陆河流域研究站的荒漠生态系统长期定位观测场进行。该观测场(面积约3 hm2)于2003年5月用刺丝围封后用于荒漠生态系统植被及土壤水分和养分变化动态的长期监测样地。观测场位于黑河流域中游的人工绿洲边缘(39°24′N,100°07′E),地势平坦,海拔1 350 m,属于大陆性干旱荒漠气候。冬季漫长寒冷,春季干旱少雨,降雨主要集中在7、8、9三个月,秋季比较湿润。年平均降水量117.1 mm,年平均温度7.6℃,年平均蒸发量2 390 mm[17]。2008年及近5年(2004-2008)的月降水量和月平均温度的变化动态如图1所示。
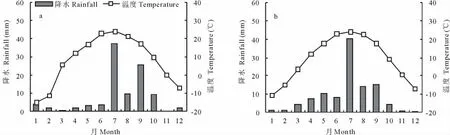
图1 2008年各月降水量及月平均气温(a)及近5年(2004-2008)各月降水量和月平均气温(b)Fig.1 Monthly rainfall and mean temperatures in the study area in 2008(a)and over the 5 years between 2004 and 2008 (b)
研究区的地带性土壤为灰棕漠土,土壤母质为第四纪砂砾洪积-冲积物。0~20 cm土层的主要理化性质是:粗砂(>2 mm)占(9.5±6.9)%;中细砂(0.05~2 mm)占(82.6±12.6)%;粘粉粒(<0.05 mm)占(5.9±2.8)%;p H 值(7.86±1.07);容重(1.43±0.21)g/cm3;有机碳含量(1.21±0.04)g/kg;总氮含量(0.13±0.01)g/kg;总磷含量(0.75±0.01)g/kg。研究区是典型的干旱荒漠灌丛植被,总盖度(15.7±0.02)%,优势灌木种是红砂(Reaumuriasoongorica)和泡泡刺(Nitrariasphaerocarpa),草本层植物主要有多年生沙葱(Alliummongolicum)、蝎虎霸王(Zygophyllummucronatum)及一年生草本植物盐生草(Halogetonglomeratus)和黄蒿(Artemisiascoparia)等[18]。
1.2 野外观测方法和动物样品鉴定
在荒漠生态系统观测场内,随机选取50 m×100 m的样区作为地面节肢动物调查样区,采用国际通用的陷阱捕获法(pitfall trap)调查样区内的动物群落组成与数量[19-21]。具体取样方法是:在样区内设置4条平行的样线(间距10 m),在每条样线上等距离布设陷阱收集器6个(间距10 m),共计24个陷阱收集器。由于研究区降水集中在7、8、9三个月,存在明显的干季和湿季,所以确定在2008年的春季(6月上旬)和秋季(9月中旬)2个典型时期对地面节肢动物群落进行调查,每次调查采样期均为14 d[22]。
将捕获到的节肢动物样品带回实验室,采用实体显微镜和生物显微镜进行种类鉴定和个体数量统计。对节肢动物的分类鉴定主要依据尹文英编写的《中国土壤动物检索图鉴》[23]、郑乐怡和归鸿编写的《昆虫分类》[24]和胡金林编写的《中国农林蜘蛛》[25]等权威分类著作。在本研究中,绝大多数动物样品鉴定到科,但捕获到的螳螂目昆虫仅有1头,且样品残缺,仅鉴定到目。
1.3 数据分析
将已鉴定出的地面节肢动物样品用万分之一的微量天平称量湿重。依据节肢动物不同类群在群落总个体数和总生物量中所占比例多少将其划分为优势类群(>10%)、常见类群(介于1%~10%)和稀有类群(<1%)。同时,依据有关文献和分类专著将收集到的地面节肢动物依据其取食类型不同划分为植食性、捕食性、腐屑食性和杂食性4个营养功能群[26,27]。Hurlbert稀疏法是一种常见的研究物种丰富度变化特征的方法[28],本研究采用物种稀疏曲线来分析地面节肢动物丰富度的季节变异规律。采用配对样本t检验法对6和9月2个调查期地面节肢动物的个体数量、群落类群数和生物量的差异进行比较。
采用非线性多维标度法(non-metric multidimensional scaling,NMDS)对6和9月地面节肢动物群落进行了分类排序[29]。NMDS是近年来发展起来的一种适用于非线性复杂数据结构分析的迭代排序方法,越来越多的研究表明它能够很好地揭示连续梯度变化关系[30-33]。在本研究中,NMDS排序所使用的数据库是每个收集器收集的节肢动物的个体数量和生物量,排序分析过程选用Sorensen(Bray-Curtis)指数来比较6和9月动物群落结构的差异,并计算了不同动物类群与排序轴的Kendall相关系数(相关系数用tau来表示)。排序分析结果给出了Stress值,它用来表征由NMDS确定的n个初始排序点在多大程度上反映了n个实体之间的实际差异的大小。Stress<5,表示排序结果很好;Stress<10,表示排序结果较好;Stress<20,表示排序结果尚可;Stress>20,则表示排序结果较差[28]。
2 结果与分析
2.1 荒漠地面节肢动物群落的结构与数量特征
在6和9月2个调查期共捕获节肢动物3 082头(只),隶属2纲12目26科,其中昆虫纲动物占94.3%,蛛形纲动物占5.7%(表1)。根据个体数量划分,节肢动物群落的优势类群是长椿科、蚁科和拟步甲科动物,它们的个体数量分别占群落个体总数的30.9%,27.8%和19.5%;常见类群包括象甲科、蝗总科、皿网蛛科、平腹蛛科和蟹蛛科,其个体数量总和占群落个体总数的10.6%;稀有类群包括19个科,其个体数量总和占群落个体总数的5.1%。根据生物量划分,优势类群是拟步甲科和象甲科,其生物量分别占群落总生物量的76.9%和12.8%;常见类群包括夜蛾科幼虫、步甲科、平腹蛛科和蚁科,其生物量总和占群落总生物量的7.7%;稀有动物类群包括21个科,其生物量总和占群落总生物量的2.6%。
2.2 节肢动物群落结构与数量的季节变异特征
采用Hurlbert稀疏曲线分析和统计分析结果均表明,6和9月地面节肢动物群落的组成、数量和多样性存在显著差异(图2,3)。6月地面节肢动物的丰富度高于9月,6月群落的个体数、生物量和类群数也显著高于9月(群落个体数:t=4.25,P=0.003;群落生物量:t=4.73,P<0.001;群落类群数:t=6.03,P<0.001)。另外,6和9月地面节肢动物群落4个不同营养功能群的个体数量和生物量组成也存在明显差异(图4)。无论是从个体数量还是从生物量来看,在2个时期腐屑食性类群都是荒漠地面节肢动物的重要功能类群。从个体数量的变化看,从6月到9月,植食性和腐屑食性2个功能群所占比重明显降低,捕食性功能群所占比重明显增加,而杂食性功能群所占比重变化不明显(图4a,b)。从生物量的变化看,6月到9月腐屑食性功能群所占比重明显降低,植食性和杂食性所占比重明显增加,而捕食性所占比重略有增加(图4c,d)。
2.3 地面节肢动物群落分布的NMDS排序分析
利用6和9月地面节肢动物的个体数量数据进行NMDS排序分析结果表明,排序轴1和2存在明显的正交性(73%,r=0.519),这2个排序轴累计解释了94.7%的数据变异(排序轴1:R2=0.66;排序轴2:R2=0.287),说明前2个排序轴较好地反映了地面节肢动物群落结构(图5a)。对生物量的NMDS排序分析结果表明,排序轴1和2也存在显著的正交性(85%,r=0.389),前2个排序轴累计解释了94.1%的数据变异(排序轴1:R2=0.66;排序轴2:R2=0.287),同样较好地反映了地面节肢动物群落结构(图5b)。在6和9月2个调查期对个体数量和生物量的NMDS排序分析计算出的Stress值分别为8.5和8.7,表明NMDS排序分析是合理的。个体数量的NMDS排序轴1和生物量的NMDS排序轴2分别反映了6月和9月地面节肢动物群落结构的差异,而个体数量的排序轴2和生物量的排序轴1分别反映了同一调查时期不同生境之间地面节肢动物群落结构的差异。此外,对不同类群节肢动物与排序轴1和2的Kendall相关分析结果表明,不同类群的个体数量和生物量存在明显的季节变异性,这种变异性的大小可以用不同类群的个体数量和生物量与排序轴的tau值的大小来反映(表2)。从6月到9月,腐屑食性的拟步甲科以及植食性的蝗总科和长椿科的个体数量和生物量均明显下降,而象甲科的个体数量和生物量明显增加;捕食性的步甲科的生物量明显增加、球蛛科的个体数量显著降低,而蟹蛛科的个体数量显著增加;杂食性的蚁科的个体数量和生物量均明显增加(表2)。

表1 荒漠地面节肢动物不同类群个体数量(生物量)的相对多度Table 1 Relative abundance of individuals(biomass)of different groups of the ground arthropod communities %
3 讨论
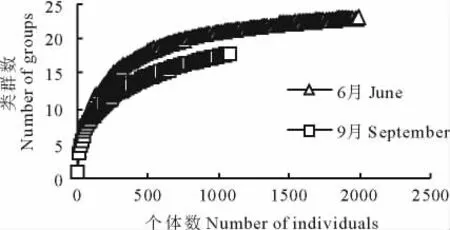
图2 6和9月地面节肢动物丰富度稀度曲线Fig.2 Hurlbert rarefaction curves of the ground dwelling arthropod assemblages in June and September censuses
本研究采用陷阱法对黑河中游典型干旱荒漠草地的地面节肢动物群落进行了系统定位观测,首次发现干旱荒漠生态系统的地面节肢动物群落由27个不同类群动物组成,其中鞘翅目的拟步甲科动物是研究区地面节肢动物群落的主要优势类群。因此,拟步甲科动物是干旱荒漠生态系统的重要组分之一,在干旱荒漠生态系统物质分解和以碳氮等元素为核心的生物地球化学循环过程中发挥着关键作用[6,9,10]。国内学者张大治等[34]在宁夏白芨滩、刘永江等[35],刘新民和杨劼[36]在宁夏中卫沙坡头干旱沙漠生态系统和内蒙古半干旱科尔沁典型沙地生态系统的调查结果均表明,拟步甲科是我国荒漠半荒漠地区地面节肢动物群落的主要类群,这与笔者在黑河中游的观测结果相一致。此外,一些学者还研究发现,拟步甲科动物适应干旱少雨的 环 境,它 主 要 分 布 在 荒 漠 半 荒 漠 地 区[37,38],Stapp[39]在北美荒漠草原、Mazía等[40],Sackmann和Flores[33]在南美荒漠草原的研究表明环境多变的春季拟步甲科的种类和数量明显高于秋季。本研究结果显示,干旱少雨的春季拟步甲科动物的个体数量和生物量均显著高于温暖湿润的秋季。
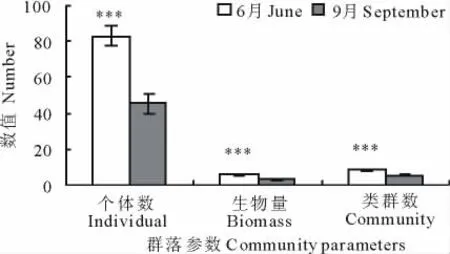
图3 地面节肢动物群落的个体数、类群数和生物量的季节变异特征Fig.3 Seasonal variation in the numbers of individuals and groups as well as biomass of the ground dwelling arthropod communities
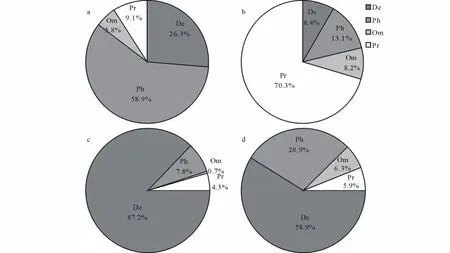
图4 6和9月2个调查期地面节肢动物不同营养功能群的个体数量和生物量相对多度的变异Fig.4 Differences in relative abundances of individuals and biomass of different functional groups of the ground dwelling arthropod communities between June and September censuses
研究结果还表明,荒漠地面节肢动物可划分为腐屑食性、植食性、杂食性和捕食性4个主要的营养功能类群。然而,4个营养功能类群的组成存在明显的季节变异性,并主要反映在以下3个方面:一是春季腐屑食性和植食性2个功能类群在群落组成中所占比重较大,而秋季捕食性类群所占比重较大;二是春季腐屑食性类群在群落总个体数中所占比重要远远高于其他3个类群,但在秋季其所占比重显著下降;三是尽管春季植食性和杂食性2个功能类群所占比重较低,但在秋季它们所占比重显著增加。许多研究同样发现土壤动物对环境条件季节变化的生理生态适应能力不同,因而在荒漠生态系统中土壤动物群落的功能群多样性存在明显的季节变化特征[1,6,41]。例如,Polis[1]在非洲Namib荒漠的研究结果显示,从干旱期到湿润期,捕食性类群与食腐屑类群和杂食性类群的比值从1∶2下降为1∶7.4。在本研究中,从春季干旱期到秋季湿润期,地面节肢动物群落中捕食性类群与食腐屑类群和杂食性类群的个体数量比值从1∶2.3下降到1∶5.5,生物量比值从1∶0.05下降到1∶0.09,类群数比值从1∶6.1下降到1∶9.6,该结果与在Namib荒漠的研究结果相一致。由于荒漠地面节肢动物营养功能群的数量和生物量组成与分解食物网结构密切相关,因此营养功能群的数量和生物量组成的季节变化将会在一定程度上影响荒漠生态系统的物质循环和能量流动过程[6,41]。
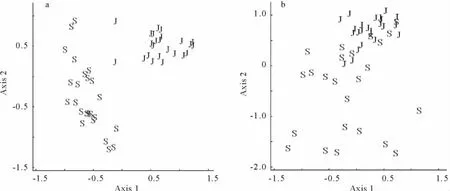
图5 6月和9月地面节肢动物个体数量和生物量的NMDS排序图Fig.5 NMDS plots indicating 2-dimensional distances of the ground dwelling arthropod communities between June and September censuses
综合应用物种稀疏曲线分析法和NMDS排序分析法对荒漠地面节肢动物群落分布特征的研究表明,节肢动物群落的个体数量、生物量、类群数以及群落丰富度存在显著的季节差异。此外,依据6和9月不同类群与排序轴的Kendall相关系数的正负和大小,可将27个地面节肢动物类群对季节变化的响应模式划分为4种基本类型。第1种响应类型是,从6月到9月动物个体数量和生物量显著增加,属于该种响应类型的动物类群有蚁科(属于杂食性动物)和象甲科(属于植食性动物)等。这说明温暖湿润的秋季是这些动物类群的成虫活动时期[23,42]。第2种响应类型是,从6月到9月动物个体数量和生物量显著下降,属于该种反应类型的动物类群有拟步甲科、蝗总科和长椿科等,这些类群主要属于腐屑食性和植食性动物。说明这些类群的动物能适应干旱少雨的环境。已有的研究表明,春季是拟步甲科动物许多种类的繁殖季节[10,38]。第3种响应类型是,从6月到9月动物个体数量或生物量有明显的变化,属于该种反应类型的动物类群有步甲科、球蛛科和蟹蛛科等。由于这些类群均属于捕食性动物,所以其种群数量的季节变化可能是因食物资源的季节变化引起的[6,26]。第4种响应类型是,从6月到9月动物个体数量和生物量无明显的变化,大部分常见和稀有类群属于该种反应类型,这说明气候因子(主要是气温和降水)的季节变化对它们的生长和繁殖过程的影响程度较小。
综上分析表明,气候因子(主要是降水和温度)的季节变化可能直接或间接地导致了动物栖息地的生物(如植被)和非生物环境(地表温度和土壤水分含量等)条件的季节变化,从而进一步对节肢动物种群和群落的生长和繁殖过程产生了深刻影响[37,42,43]。然而,不同动物类群在对气候的季节变化的响应模式和适应机理上存在明显的差异,这充分反映了它们在生活史、营养方式、繁殖特征和生物学特性等方面的综合性差异[44]。深入研究不同类群荒漠地面节肢动物的生活史、营养方式、繁殖特征和生物学特性,对于进一步解析荒漠地面节肢动物群落结构、组成与功能群多样性的时空分布变异规律是非常重要的。
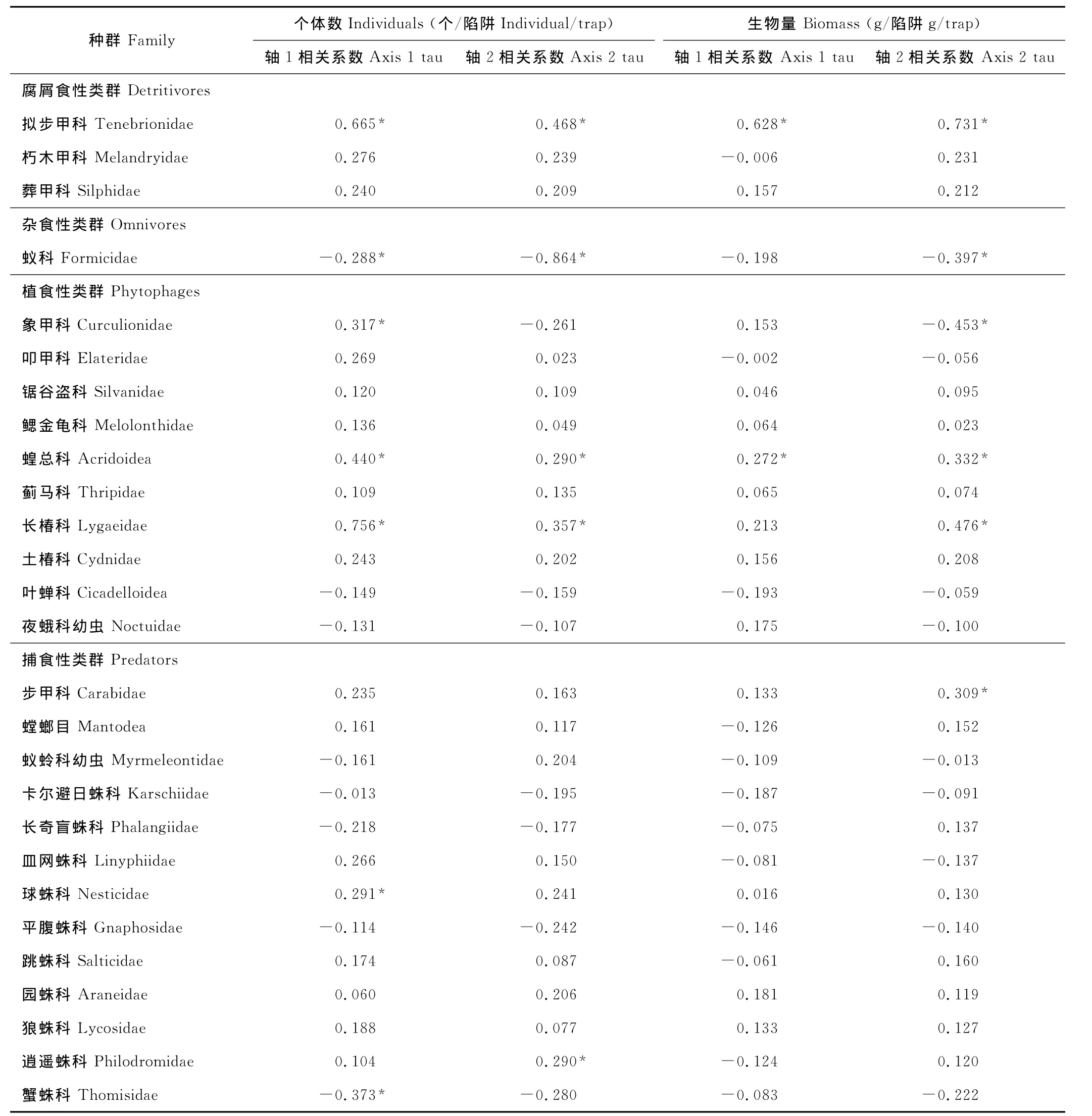
表2 不同类群地面节肢动物与NMDS排序轴1和2的Kendall相关系数Table 2 Kendall correlation coefficients with the first and second ordination axes of different groups of the ground dwelling arthropod communities(n=48)
4 结论
拟步甲科动物是黑河中游干旱荒漠生态系统地面节肢动物群落最重要的组分,在维持干旱荒漠生态系统的生态服务功能、生物多样性和食物网络结构等方面发挥着关键作用。荒漠地面节肢动物群落结构及其功能群组成和数量表现出明显的季节变异性,主要体现在以下3个方面:一是春季地面节肢动物群落的个体数量、生物量和类群丰富度显著高于秋季;二是腐屑食性的拟步甲科、植食性的长蝽科、蝗总科和捕食性的球蛛科动物的主要活动期是在春季,而杂食性的蚁科、捕食性的蟹蛛科和步甲科及植食性的象甲科的主要活动期是在秋季;三是春季捕食性类群与杂食性和腐屑食性类群的比值要高于秋季。
[1]Polis G A.The Ecology of Desert Communities[M].Tucson:The University of Arizona Press,1991.
[2]Whitford W G.Ecology Desert Systems[M].San Diego:Academic Press,2002.
[3]潘晓玲,马应军,高炜,等.中国西部干旱区生态环境演变过程[J].中国沙漠,2004,24(6):663-673.
[4]Crawford C S.Biology of Desert Invertebrates[M].New York:Springer-Verlag,1981.
[5]Tigar B J,Osborne P E.Patterns of arthropod abundance and diversity in an Arabian desert[J].Ecography,1997,20:550-558.
[6]Shachak M,Gosz J R,Pickett S T A,etal.Biodiversity in Drylands:Toward a Unified Framework(Long-Term Ecological Research Network Series)[M].New York:Oxford University Press,2005.
[7]Swift M J,Heal O W,Anderson J M.Decomposition in Terrestrial Ecosystems[M].Berkeley and Los Angeles:University of California Press,1979.
[8]吴鹏飞,刘兴良,刘世荣.米亚罗亚高山草甸冬春两季土壤动物群落特征的比较[J].草业学报,2009,18(5):123-129.
[9]Whitford W G.The importance of the biodiversity of soil biota in arid ecosystems[J].Biodiversity and Conservation,1996,5:185-195.
[10]Seastedt T R,Crossley D A Jr.The influence of arthropods on ecosystems[J].Bioscience,1984,34(3):157-161.
[11]尚占环,任国华,龙瑞军.土壤种子库研究综述——规模、格局及影响因素[J].草业学报,2009,18(1):144-154.
[12]任国栋,于有志.中国荒漠半荒漠的拟步甲科昆虫[M].保定:河北大学出版社,1999.
[13]Hooper D U,Bignell D E,Brown V K,etal.Interactions between aboveground and belowground biodiversity in terrestrial ecosystems:Patterns,mechanisms,and feedbacks[J].BioScience,2000,50:1049-1061.
[14]Wardle D A,Bardgett R D,Klironomos J N,etal.Ecological linkages between aboveground and belowground biota[J].Science,2004,304:1629-1633.
[15]尹文英.中国土壤动物[M].北京:科学出版社,2000.
[16]傅声雷.土壤生物多样性的研究概况与发展趋势[J].生物多样性,2007,15(2):109-115.
[17]何志斌,赵文智.黑河流域荒漠绿洲过渡带两种优势植物种群空间格局特征[J].应用生态学报,2004,15(6):947-952.
[18]张德魁,马全林,刘有军,等.河西走廊荒漠区一年生植物组成及其分布特征[J].草业科学,2009,26(12):37-41.
[19]Brennan K E C,Majer J D,Reygaert N.Determination of an optimal pitfall trap size for sampling spiders in a Western Australian Jarrah forest[J].Journal of Insect Conservation,1999,3:297-307.
[20]Gotelli N J,Colwell R K.Quantifying biodiversity:Procedures and pitfalls in the measurement and comparison of species richness[J].Ecology Letter,2001,4:379-391.
[21]Ward D F,New T R,Yen A L.Effects of pitfall trap spacing on the abundance,richness and composition of invertebrate catches[J].Journal of Insect Conservation,2001,5:47-53.
[22]Ziesche T M,Roth M.Influence of environmental parameters on small-scale distribution of soil-dwelling spiders in forests:What makes the difference,tree species or microhabitat?[J].Forest Ecology and Management,2008,255:738-752.
[23]尹文英.中国土壤动物检索图鉴[M].北京:科技出版社,1992.
[24]郑乐怡,归鸿.昆虫分类[M].南京:南京师范大学出版社,1999.
[25]胡金林.中国农林蜘蛛[M].天津:天津科学技术出版社,1984.
[26]Coleman D C,Crossley D A Jr,Hendrix P F.Fundamentals of Soil Ecology[M].Oxford:Elsevier Academic Press,2004.
[27]贺达汉,赵紫华,张大治.草原景观下昆虫群落及种群对生境破碎化的反应[J].草业学报,2009,18(6):235-241.
[28]钱迎倩,马克平.生物多样性研究的原理与方法[M].北京:中国科学出版社,1994:141-165.
[29]McCune B,Grace J B.Analysis of Ecological Communities[M].Oregon:MJM Software Design,2002.
[30]Hyrenbacha K D,Veitb R R,Weimerskirch H,etal.Community structure across a large-scale ocean productivity gradient:Marine bird assemblages of the Southern Indian Ocean[J].Deep Sea Research Part I:Oceangraphic Research Papers,2007,54:1129-1145.
[31]林英华,宋百敏,韩茜,等.北京门头沟废弃采石矿区地表土壤动物群落多样性[J].生态学报,2007,27(1):4832-4839.
[32]Hebshi A J,Duffy D C,Hyrenbach K D.Associations between seabirds and subsurface predators around Oahu,Hawaii[J].Aquatic Biology,2008,4:89-98.
[33]Sackmann P,Flores G E.Temporal and spatial patterns of tenebrionid beetle diversity in NW Patagonia,Argentina[J].Journal of Arid Environments,2009,73:1095-1102.
[34]张大治,贺达汉,于有志,等.宁夏白芨滩国家级自然保护区地表甲虫群落多样性[J].动物学研究,2008,29(5):569-576.
[35]刘永江,关宏斌,郭砺.科尔沁沙地土壤动物研究初探[J].中国沙漠(增刊1),1999,19(1):115-119.
[36]刘新民,杨劼.干旱、半干旱区几种典型生境大型土壤动物群落多样性比较研究[J].中国沙漠,2005,25(2):216-222.
[37]Thomas D B Jr.Tenebrionid beetle diversity and habitat complexity in the eastern Mojave desert[J].The Coleopterists Bulletin,1983,37(2):135-147.
[38]任国栋,于有志,侯文君.中国荒漠半荒漠地区拟步甲的组成和分布特点[J].河北大学学报(自然科学版),1999,19(2):176-183.
[39]Stapp P.Microhabitat use and community structure of darkling beetles(Coleoptera:Tenebrionidae)in shortgrass prairie:Effects of season shrub and soil type[J].American Midland Naturalist,1997,137(2):298-311.
[40]Mazía C N,Chaneton E J,Kitzberger T.Small-scale habitat use and assemblage structure of ground-dwelling beetles in a Patagonian shrub steppe[J].Journal of Arid Environments,2006,67:177-194.
[41]Whitford W G.Abiotic controls on the functional structure of soil food webs[J].Biology and Fertility of Soils,1989,8:1-6.
[42]Schumacher A,Whitford W G.Spatial and temporal variation in Chihuahuan desert ant faunas[J].The Southwestern Naturalist,1976,21(1):1-8.
[43]Ayal Y.Trophic structure and the role of predation in shaping hot desert communities[J].Journal of Arid Environments,2007,68:171-187.
[44]Staley J T,Hodgson C J,Mortimer S R,etal.Effects of summer rainfall manipulations on the abundance and vertical distribution of herbivorous soil macro-invertebrates[J].European Journal of Soil Biology,2007,43:189-198.
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15
中国石化(2021年8期)2021-11-05
科学大众(中学)(2021年3期)2021-05-17
绿色中国(2019年14期)2019-11-26
江苏农业科学(2019年5期)2019-09-02
课外生活(小学1-3年级)(2018年10期)2018-11-19
小猕猴学习画刊(2018年9期)2018-09-22
广东农业科学(2017年5期)2017-08-29
文史春秋(2016年3期)2016-12-01
西南农业学报(2016年4期)2016-05-17
