
土壤水分胁迫对大叶黄杨和月季光合生理特性的影响
2010-05-07 11:11:44谭雪红郭小平
水土保持通报 2010年4期
谭雪红,郭小平,王 亮
(1.北京林业大学水土保持学院,北京100083;2.徐州工程学院,徐州江苏221008)
土壤水分是保证植物正常生长发育的主要因素,是干旱区植物生长的最主要限制因子。在目前水资源日益短缺的情况下,如何选择耐旱的城市园林绿化植物、合理有效地利用有限水资源,是国内外学者广泛关注的问题[1-2]。土壤干旱对植物生长和代谢的影响是多方面的,其中对光合生理特性影响尤为突出和重要。近年来,有关植物光合生理过程及其环境影响因子的研究比较活跃,对多级水分胁迫下植物光合生理研究也已有报道[3-6]。土壤水分胁迫下植物光合生理特性研究为抗旱植物的选择、节水灌溉、管理等提供了依据,对于解决植物生长水分供需矛盾有重要的指导意义。
月季(Rosa chinensis)和大叶黄杨(Euonymus japonicus L)为我国园林绿化常用植物。月季具有观赏效果好形状独特、花香浓、花期长、花色多样等优点,具有很高的审美价值,在环城行道、公园、风景区等一切有绿地的地方,月季都能起到画龙点睛的美化功效;大叶黄杨是常绿灌木,具有叶色光亮,嫩叶鲜绿,极耐修剪等优点,是良好的绿篱植物,在我国各省,特别是中部和北部城市普遍栽植。目前关于月季和大叶黄杨光合生理特性及与环境关系方面的研究已见报道[7-9],但不同土壤水分胁迫下光合生理特性变化规律的研究尚未有报道。因此,本研究通过对月季和大叶黄杨在不同土壤水分胁迫下光合生理特性变化规律的分析,为干旱地区这两种植物的种植、管理提供一定的参考依据。
1 材料及方法
1.1 材料及处理
试验苗木大叶黄杨、月季来自北京附近苗圃基地,均为一年生苗木。试验地点设在北京林业大学校园内。采用盆栽的方法进行,花盆规格为30 cm×40 cm(直径×高),盆栽所用土壤来源于北京附近公路的中央隔离带,沙质壤土,容重为1.067 g/cm3,pH为7.74,速效磷含量为5.45 mg/kg,速效钾含量为186.65 mg/kg,土壤有机质为6.425‰,土壤田间持水量为28.9%。
于2009年5月下旬将长势一致的苗木移植至花盆中,每个树种各植15盆,每盆一株,共30盆。定期浇水,精心养护,使苗木适应盆栽环境,植株恢复生长2个月后一次性浇足水,选择长势较好的植株进行水分胁迫试验。采用自然干燥法控水,土壤相对含水量(土壤含水量占田间持水量的百分率)设为:90%~95%,60%~65%,40%~45%和20%~25%共 4个梯度,其中土壤相对含水量90%~95%为植物的正常供水水平,用CK表示;60%~65%,40%~45%和20%~25%分别代表轻度、中度和重度干旱胁迫处理,用W1,W2和W3表示。每个处理设置3个重复。采用秤重法监控土壤含水量,每天20:00时用灵敏度为1/1 000的电子天平称盆重,补充蒸发的水分。
1.2 测定指标
干旱胁迫后第10 d进行4种水分处理下叶绿素荧光和光合作用参数的测定。利用Hansatech公司生产的FMS-2型调制式叶绿素荧光分析仪测定暗适应后的最小荧光)、最大荧光()、可变荧光Fv()、PSII原初光能转换效率()、PSII潜在活性(),参考许大全的方法[10],在测定前一天晚上将花盆搬入暗室,植物经过一夜充分暗适应后,于次日凌晨7:00左右测定。利用Li-6400便携式光合作用测定仪测定两植物不同水分胁迫下的净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)、蒸腾速率()等光合作用参数,根据测定的光合速率和蒸腾速率的比值),计算出水分利用效率(WUE)。测定时间从8:00—18:00,每2 h测定一次。所有测定在同一天进行,测定选用植株中部生长健壮的成熟叶片,每次测定3次重复,取平均值。
1.3 数据处理
利用Excel进行试验数据处理和图表的绘制,利用SPSS 13.0软件进行方差分析(ANOVA)和t检验。
2 结果分析
2.1 土壤水分条件对光合作用的影响
2.1.1 主要光合参数的日变化 自然条件下,植物光合作用随着内外因子的变化而时刻发生变化。月季Pn日变化对不同程度水分胁迫的响应表现出一定差异,W3时,Pn明显降低,且日变化不明显;CK,W1和W2处理时,Pn有 “午休”现象,即在 12:00—14:00降到最低,但两峰值出现的时间不同,水分胁迫下,有上午峰值提前,下午峰值滞后的现象。Tr,Gs和Pn有相似的变化趋势,但 Gs在 CK,W1和 W2时,10:00—12:00出现第一峰值之后,具有明显的下降趋势,第二峰值不明显。Ci在CK,W1和W2时,16:00—18:00有明显的峰值。WUE的变化较为复杂,W2处理时,16:00—18:00呈现明显峰值,其它处理日变化不明显。
大叶黄杨在CK和W1处理时,Pn表现为“单峰”型,峰值都出现在 10:00—12:00,W2时表现出“双峰”型,W3时没有明显的日变化。Tr除W3处理时变化不明显外,其余3种处理都表现为“单峰”型,只是峰值出现的时间略有不同。Gs日变化规律和Tr相似。Ci有明显的峰值,但CK时峰值明显滞后。CK时的WUE一直大于3种水分胁迫,4种处理都表现出“双峰”型的特点,其日变化有一定的差异,总的来看,W3处理的WUE最低。
试验结果表明,月季和大叶黄杨主要光合参数的日变化表现出较大的不同。首先,CK和W1处理时,月季的 Pn,Tr和Gs日变化曲线表现为“双峰”型,而大叶黄杨表现为“单峰”型;其次,月季CK,W1和W2时的Pn,Tr和Gs日变化曲线排列紧密,和W3处理时的曲线之间距离较大,而大叶黄杨CK时的日变化曲线和3种胁迫的日变化曲线距离较大;第三,月季的WUE日变化曲线表现为不同处理的交替变化,而大叶黄杨CK时的日变化曲线始终处于最上部(WUE值始终最大)。
2.1.2 主要光合参数日均值 由图1可以看出,4种水分处理下,Pn,Tr和Gs都表现为:月季>大叶黄杨;WUE表现为:CK处理时,大叶黄杨>月季,其它3种水分胁迫下,月季>大叶黄杨;Ci表现为:CK处理时,月季>大叶黄杨,其它3种水分胁迫下,大叶黄杨>月季。随着水分胁迫的加重,月季和大叶黄杨的Pn和Tr都逐渐降低,和CK处理相比,从W1到W3,月季 Pn降低幅度为 13.1%,22.9%,87.6%;大叶黄杨为51.1%;62%;81%;月季 Tr降低幅度为21.4%,28.3%,83.1%;大叶黄杨为 40.8%,49%,68.9%;可以看出无论是Pn还是Tr,月季在W3处理时开始大幅度降低,而大叶黄杨在W1处理时就开始大幅度降低;大叶黄杨的WUE逐渐降低,而月季在W2处理时WUE达到最高,这说明适度的水分胁迫可以增加月季的WUE;月季的Gs逐渐降低,大叶黄杨在W2时出现了略高于W1的情况;月季的Ci先降低,W3处理时开始升高,而大叶黄杨有逐渐增加的趋势。方差分析表明,月季和大叶黄杨4种水分处理间各光合参数都差异显著(P<0.05);t检验也表明月季和大叶黄杨之间净光和速率、蒸腾速率、水分利用效率和气孔导度差异显著(P<0.05),而胞间CO2浓度差异不显著(P=0.376>0.05)。
2.2 土壤水分条件对叶绿素荧光的影响
叶绿素荧光反应是植物光化学反应的指示物,与物种、季节、环境、样品情况和其它影响植物生理作用的因素有关。因此,可测量叶绿素荧光的变化来反映植物对环境胁迫的反应[11]。
Fo和Fm分别指光系统Ⅱ(PSⅡ)反应中心处于完全开放和完全关闭时的荧光产量,Fv为可变荧光,反映原初电子受体(QA)的还原情况[12]。由图2可以看出,随着土壤水分胁迫的加强,Fo逐渐上升,Fm和Fv逐渐下降,但两种植物变化的幅度不同。与CK相比,从W1控水处理到W3处理,月季Fo增加幅度分别为5.59%,11.2%和45.4%;大叶黄杨为23.6%,29.9%,46.6%;月季 Fm降幅为 1.5%,1.9%和2.3%;大叶黄杨为10.1%,12.3%,38.4%;月季Fv降幅为2.9%,4.4%和11.9%;大叶黄杨为18%,22.1%,58.2%。可以看出,大叶黄杨的变化幅度要大于月季。方差分析表明,月季的Fo在 W1和CK,W2处理间的差异不显著(P=0.087,0.2>0.05),而大叶黄杨Fo在W1和W2处理间差异不显著(P=0.06>0.05),其余各处理间差异显著。Fm和Fv具有相同的规律,月季的Fm和Fv在W3和CK 差异显著(P=0.009,0.007<0.05),其余各处理间差异不显著,大叶黄杨的Fm和Fv在W1和W2处理间差异不显著(P=0.59,0.71>0.05),其余各处理间差异显著。
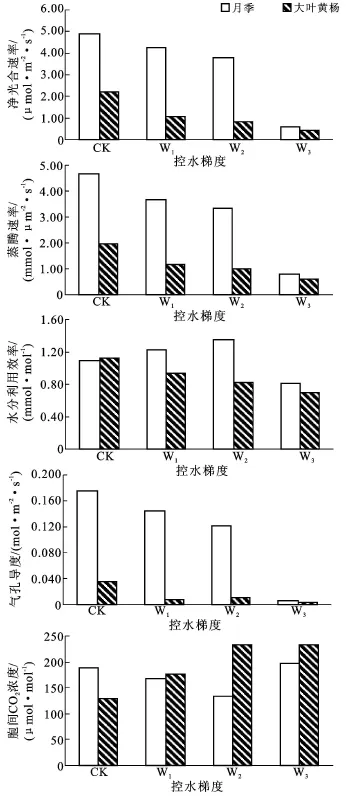
图1 水分胁迫对月季、大叶黄杨主要光合参数日均值的影响注:CK,W1,W2,W3分别为正常供水,轻度干旱胁迫,中度干旱胁迫,重度干旱胁迫处理。下同。
荧光参数Fv/Fm是反映PSⅡ的光化学转化效率的一个常用参数,称为PSⅡ原初光能转化效率,Fv/Fo表示光反应中心PSⅡ的潜在活性,非环境胁迫条件下这两参数极少变化,不受物种和生长条件的影响,胁迫条件下会有下降趋势[12]。CK时,月季和大叶黄杨的Fv/Fm值差别不大,
但随着土壤水分胁迫的加重,两植物都有逐渐下降的趋势,而且Fv/Fm的差异越来越明显,这说明大叶黄杨PSⅡ活性中心受损程度可能要远远大于月季。和CK相比,月季从W1到W3降幅为1.2%,2.4%和9.6%,大叶黄杨为 8.6%,11.1%,32.2%,大叶黄杨的降幅较大(图 2)。方差分析表明,月季W1和CK ,W2(P=0.086,0.161>0.05)差异不显著,其余各处理间差异显著,而大叶黄杨各处理间均达到显著差异。Fv/Fo和Fv/Fm有类似的规律,但下降幅度有一定的差异,由图2可以看出,随着水分胁迫的增加,其下降趋势更加明显,大叶黄杨的下降幅度更大,方差分析表明无论月季还是大叶黄杨,各处理间都达到显著差异的水平。
3 结论与讨论
Pn,Tr,Gs和Ci是植物光合作用状况的最重要指标,植物遭受土壤水分胁迫,其光合作用必然受到影响,最终影响植物生长。土壤水分胁迫下,引起植物叶片Pn降低的植物自身因素主要有气孔部分关闭导致的气孔限制和叶肉细胞光合活性下降导致的非气孔限制两类,前者使Ci降低,而后者使Ci升高[4,13-14]。
在W2及以下程度水分胁迫处理下,月季Gs和Ci随着胁迫的加重而降低,表明其光合能力的下降主要由于气孔因素限制;W3处理时Gs降低而Ci升高(图1),表明月季Pn降低的原因已经不只是可逆的气孔收缩,而是光合器官受到损伤。随着水分胁迫的加重,大叶黄杨Gs降低而Ci升高,表明W1处理时,Pn的降低主要由非气孔因素所导致,其叶片光合器官已经受到损伤。从月季和大叶黄杨光合参数的日变化可以看出月季Pn和Tr出现了“午休”现象,大叶黄杨没有,而“午休”有利于保持叶片水分,提高植物的抗旱性。W3处理是土壤水分对月季伤害的转折点,而W1处理是土壤水分对大叶黄杨伤害的转折点。当植物遭受水分胁迫时,其WUE必定会发生相应的变化,以保持植物生长与水分消耗的平衡,由于土壤水分胁迫同时对Pn和Tr产生了影响,所以WUE的高低就取决于两者被影响的程度。月季W3时水分利用效率最高,而大叶黄杨在CK时最高,这充分说明了W1已经对大叶黄杨叶片造成了伤害。
水分胁迫对植物光合作用的影响是多方面的,不仅直接引发光合机构结构和功能的异常,同时也影响光合电子的传递[15]。水分胁迫下,月季和大叶黄杨叶绿素荧光参数 Fo 升高,Fm ,Fv,Fv/Fm ,Fv/Fo 降低 ,说明水分胁迫影响了两植物PSⅡ的活性、光合电子的传递过程及CO2同化的正常进行。但月季和大叶黄杨各参数的变化有很大差异,随着水分胁迫的加重,大叶黄杨各参数的变化幅度均大于月季。各荧光参数不同水分处理下的方差分析表明,与CK相比,月季在W3处理时才达到极显著差异的水平(P<0.01),而大叶黄杨在W1处理时就达到极显著差异(P<0.01)。
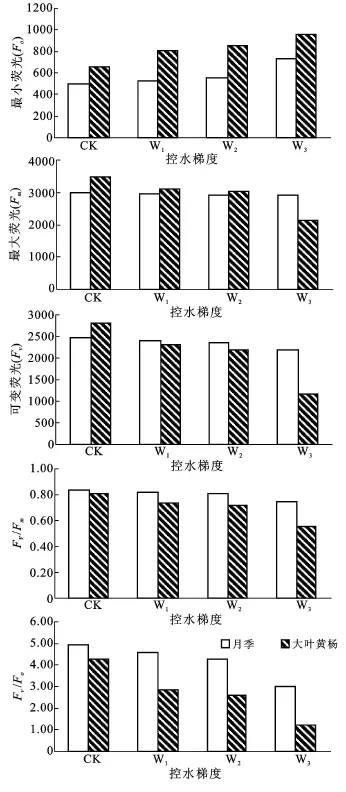
图2 水分胁迫对月季、大叶黄杨主要叶绿素荧光参数的影响
月季和大叶黄杨在水分胁迫下各荧光参数和光合作用参数变化取得了较为一致的结论。即水分胁迫对月季和大叶黄杨的光合生理产生较大影响,但二者变化幅度有所差异,大叶黄杨对土壤水分胁迫更为敏感。
[1] 李志军,罗青红,伍维模,等.干旱胁迫对胡杨和灰叶胡杨光合作用及叶绿素荧光特性的影响[J].干旱区研究,2009,26(1):45-52.
[2] 王颖,魏国印,张志强,等.7种园林树种光合参数及水分利用效率的研究[J].河北农业大学学报,2006,29(6):44-48.
[3] 朱教君,康宏樟,李智辉,等.水分胁迫对不同年龄沙地樟子松幼苗存活与光合特性影响[J].生态学报,2005,25(10):2527-2533.
[4] 柯世省.干旱胁迫对夏蜡梅光合特性的影响[J].西北植物学报,2007,27(6):1209-1215.
[5] 柯世省,杨敏文.水分胁迫对云锦杜鹃光合生理和光温响应的影响[J].园艺学报,2007,34(4):959-964.
[6] 王强,陈存根,钱红格,等.水分胁迫对6种苗木光合生理特性的影响[J].水土保持通报,2009,29(2):144-149.
[7] 张利萍,张志平,傅松玲,等.蝴蝶兰、月季新品种生理特性的初步研究[J].安徽农业大学学报,2005,32(4):523-528.
[8] 王勇,杨培岭,任树梅,等.两种抗旱剂对大叶黄杨光合特性的耦合效应[J].应用生态学报,2007,18(3):514-518.
[9] 冯强,胡聃,李娜,等.典型城区与郊区环境大叶黄杨气体交换及叶绿素荧光特性比较[J].生态学报,2009,29(7):3477-3484.
[10] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
[11] 喻方圆,徐锡增.植物逆境生理研究进展[J].世界林业研究,2003,16(5):6-11.
[12] 黄华宏,陈奋学,童再康,等.矮生杉木光合特性及叶绿素荧光参数研究[J].北京林业大学学报,2009,31(2):69-73.
[13] Monneveux P,Rekika D,Acevedo E,et al.Effect of drought on leaf gas exchange,carbon isotope discrimination,transpi-ration efficiency and productivity in field grown durum wheat genotypes[J].Plant Science,2006,170:867-872.
[14] Farquhar G D,Sharkey T D.Stomatal conductance and photosyn-thesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[15] 胡学华,蒲光兰,肖千文,等.水分胁迫下李树叶绿素荧光动力学特性研究[J].中国生态农业学报,2007,15(1):75-77.
猜你喜欢
农村实用技术(2020年7期)2020-12-15 18:30:04
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
山东林业科技(2019年4期)2019-09-09 09:25:04
中国果业信息(2019年1期)2019-01-05 17:41:42
生物学教学(2017年9期)2017-08-20 13:22:32
高原山地气象研究(2016年2期)2016-11-10 06:06:27
安徽工业大学学报(自然科学版)(2014年4期)2014-07-11 01:45:54
食品工业科技(2014年6期)2014-05-10 06:04:50
塔里木大学学报(2014年3期)2014-03-11 18:47:27
食品工业科技(2014年13期)2014-03-11 18:17:01
