
舍饲肉羊生长期能量和蛋白质代谢特点及需要量的研究
2010-05-06 14:33:00魏炳栋邱玉朗万伶俐杨华明
饲料博览 2010年7期
魏炳栋,邱玉朗,万伶俐,杨华明,李 林
(吉林省农业科学院畜牧分院,吉林 公主岭 136100)
放牧养羊的饲养方式导致草地退化日益加剧,严重危害了我国的生态环境,而饲养方式的转变是有效解决这一问题的关键所在,也是解决人、环境、羊三者和谐健康发展的有效手段。肉羊生产由放牧改为舍饲后,其营养代谢特点及需要量发生很大变化。
我国对舍饲肉羊能量和蛋白质的代谢特点及需要量也做过一些研究,但由于饲养环境和方式的不同,研究结果差别很大。为此本试验选用东北细毛羊为试验对象,因其羊体大、生长发育快、耐粗饲适应性强,结合当地饲料资源特点,采用消化代谢试验和呼吸测热试验,研究舍饲肉羊生长期能量和蛋白质代谢特点及需要量,为当地舍饲养羊业发展提供基础参数。
1 材料与方法
1.1 试验动物
试验在吉林省农业科学院畜牧分院动物营养代谢实验室进行,选用东北细毛羊公羊6只,按体重分为两组(30和40 kg),分别进行了两期消化代谢试验和一期呼吸测热试验。试验时间为2008年8~11月。
1.2 试验日粮
试验日粮参考内蒙古细毛羊饲养标准,由精料、羊草和苜蓿草粉组成,其饲喂比例为2:3:1。精料组成及日粮各组分的营养物质含量见表1和表2。

表1 精料组成及营养成分%
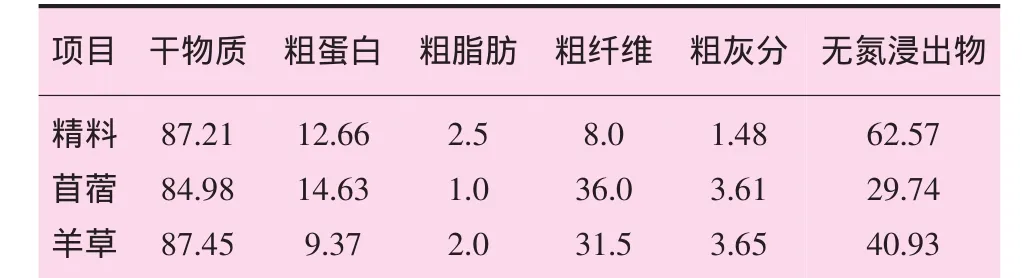
表2 日粮各组分营养物质含量 %
1.3 试验方法
1.3.1 消化代谢试验
消化代谢试验采用全收粪、尿法。设预试期5 d,正试期3 d,试验在代谢笼中进行。饲料、粪、尿样品中的氮、干物质和能值含量均按照张丽英的方法进行测定[1]。
1.3.2 呼吸测热试验
生长代谢呼吸测热试验与第二期消化代谢试验同步进行,呼吸小室温度控制在23±1℃,湿度控制在60±5%,呼吸测热过程中微机每3 min测量1次小室的气体流量、气压、湿度、O2、CO2和 CH4气体的浓度,每1 h测量1次空气中的O2和 CO2浓度,并将气体容积自动校正到标准状态。预试期5 d,正试期3 d。呼吸测热试验结果参照杨嘉实计算方法进行[2]。
1.3.3 绝食代谢
将试验羊绝食72 h后,在呼吸测热小室中进行呼吸测热试验,绝食代谢试验结果的计算参照杨嘉实的计算方法进行[2-3]。
2 结果与分析
2.1 干物质的消化率
试验羊干物质采食量及消化率见表3。
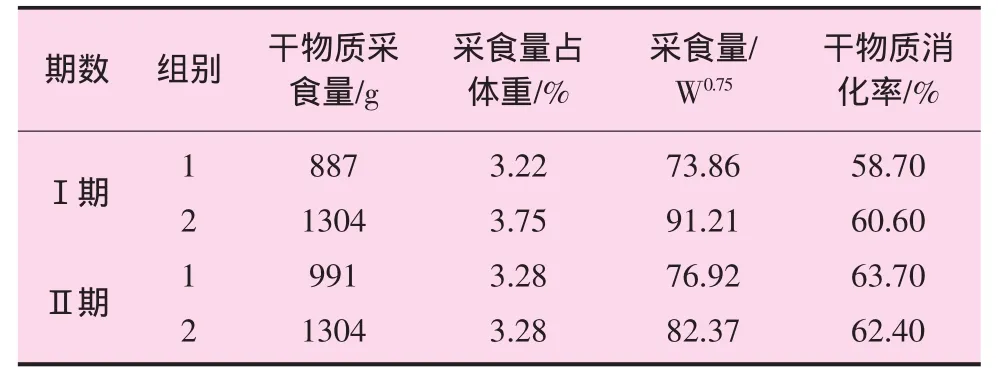
表3 试验绵羊干物质采食量及消化率
从表3可以看出,第1组的干物质采食量第Ⅱ期较第Ⅰ期略高,第2组干物质采食量两期相同,用绿草、青贮和不同干草进行的采食量和消化率研究表明,绵羊和山羊的干物质采食量分别为 55.8和 65.3 g·kg-1·W-0.75,绵羊的采食量每天为 56.9 g·kg-1·W-0.75[4]。杨维仁等报道大尾寒羊的干物质采食量为 86.6 g·kg-1·W-0.75[5]。本试验结果较国外资料高,与大尾寒羊干物质采食量相差不大,但是在报道的绵羊干物质采食量 40~120 g·kg-1·W-0.75的范围内;干物质采食量占体重比在报道的绵羊干物质采食量占体重的1.8%~3.8%的范围内,干物质消化率第Ⅰ期与杨维仁等报道的大尾寒羊前期消化率59.1%相接近,但第Ⅱ期低于后期干物质消化率70.98%。
2.2 能量代谢及呼吸测热试验
试验羊在第Ⅱ期消化代谢试验期间,进行了呼吸测热试验,结果见表4。从表4可以看出,试验羊产热主要是由脂肪分解引起的。
杨在宾等在对小尾寒羊的能量代谢和需要量的研究中,指出48 kg的空怀母羊的能量表观消化率69.9%,呼吸商为 0.872,HP 为 377 kJ·kg-1·W-0.75,与本试验结果相似[6]。
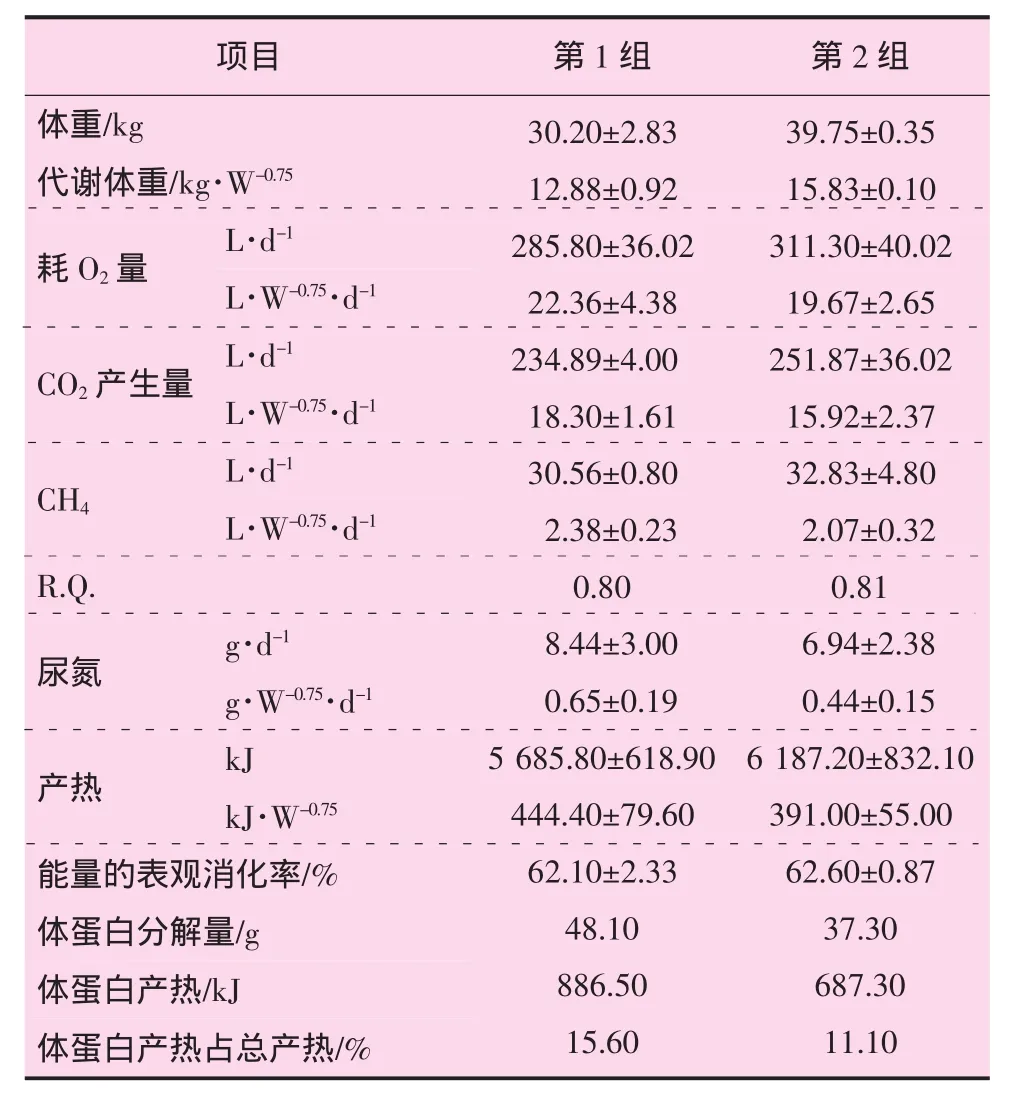
表4 舍饲肉羊生长代谢呼吸测热数据
根据生长期呼吸测热试验结果和绝食产热结果,利用能量代谢公式计算,得出舍饲肉羊能量代谢数据,见表5。
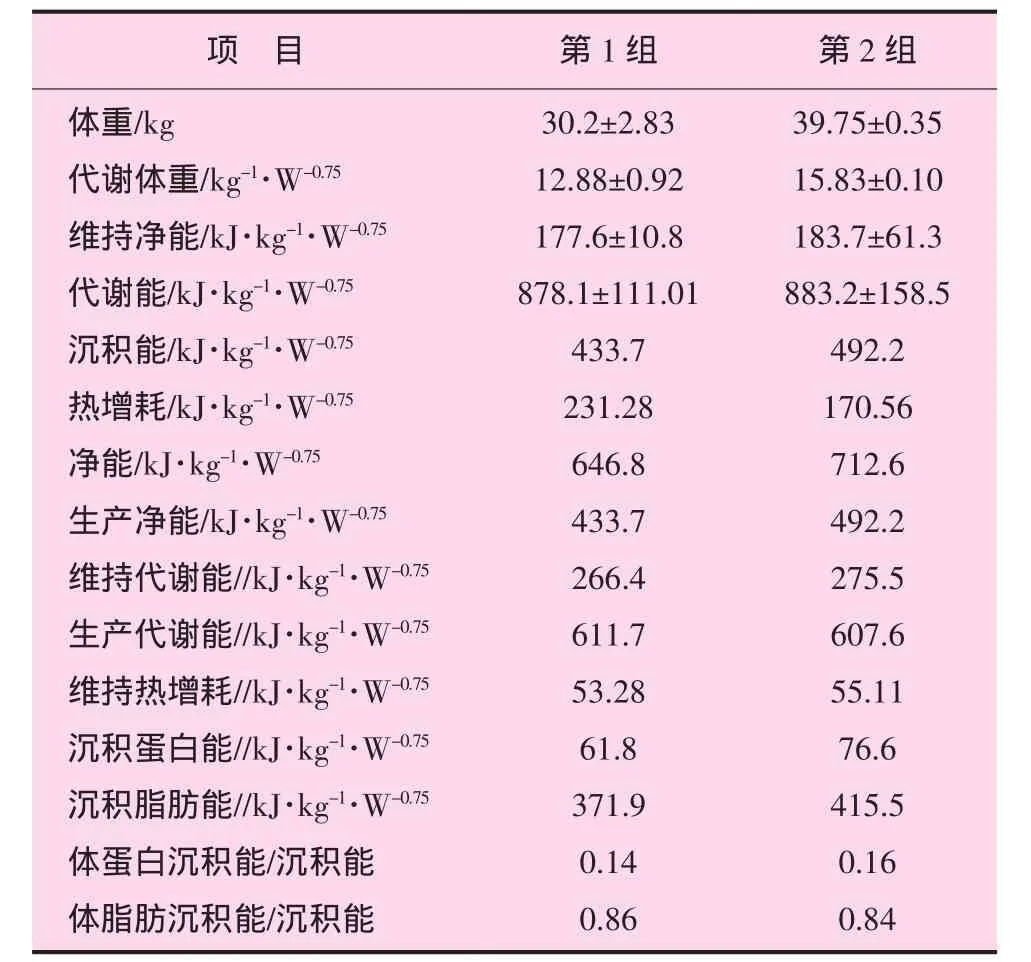
表5 舍饲肉羊能量代谢数据
赵玉民等应用自制的密闭式呼吸测热装置,对育成期的细毛羊母羊,分3个体重阶段进行24 h绝食代谢测试,得出维持代谢能(MEm)为 460.2~481.2 kJ·kg-1·W-0.75[7]。杨在宾等进行的青山羊泌乳期母羊的能量需要量研究,得出青山羊泌乳前期、后期及整个泌乳期的MEm分别为456、540和502 kJ·kg-1·W-0.75[8]。杨诗兴等测得育成期湖羊的 MEm为 38 kJ·kg-1·W-0.75[9]。赵新全等在青海海北州高寒草甸生态系统定位站采用呼吸面具的方法对9只试验羊进行呼吸测热,得出藏羊的MEm为442.75 kJ·kg-1·W-0.75,维持热增耗为 94.14 kJ·kg-1·W-0.75[10]。综合以上报道,维持代谢能的需要量各报道差异较大,主要是由于测定方法、日粮组成和外界环境的不同所致。近些年来,国外的许多学者都对山羊泌乳期的能量需要进行了大的研究,欧美等都相继制订出了本国山羊品种的能量需要量标准,并提出了评定反刍动物能量需要及营养价值的新体系[11-14]。本试验所得生长期两个体重阶段(30和 40 kg)的结果与前人研究结果一致。
2.3 代谢能利用效率的计算
代谢能利用效率,即K=NE/ME,可根据净能的种类,将K分为Km(代谢能用于维持的效率)、Kf(脂肪利用效率)、Kp(蛋白利用效率)及Kg(用于生产的效率)。Km值采用许振英、Thorober提出的代谢能用于维持的效率为80%估算,可得Km=0.8,进一步推算出维持代谢能MEm=NE/Km。
舍饲肉羊K值见表6。
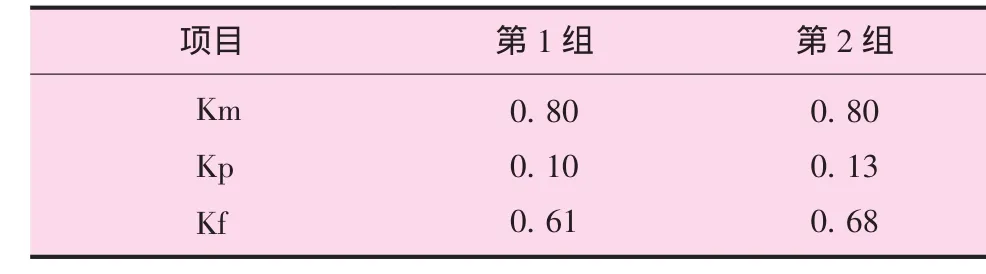
表6 舍饲肉羊的K值
2.4 舍饲肉羊生长期代谢能需要量的估算
根据析因法可知,生长代谢能包括维持代谢能和生长代谢能,见式(1)。

式(1)中 MEg为生长代谢能,MEg=NEg/Kpf;NEg为增重净能;Kpf为增重代谢能转化为增重净能的效率;Kpf=0.78ME/GE+0.0006,得出Kpf=0.4;NEg=1000△W(2.5+0.35W)。
本试验得出的舍饲绵羊生长代谢能的需要量见式(2):

式(2)中 NEg=△W(2.5+0.35W),Kpf=0.4,△W为日增重(g);W为体重(kg)。
3 小 结
舍饲东北细毛羊用于维持蛋白质需要量和生长蛋白质需要量为:
可消化粗蛋白需要量(RDCPm)=6.4 g/kg.W0.75。
生长期蛋白质需要量(RCP)=10.21+0.18△W/(0.49+0.11△W)
RDCP=6.4+0.11△W/(0.49+0.11△W)
舍饲东北细毛羊生长期代谢能需要量为(MER)=271KJ/W0.75+NEg/Kpf
其中,NEg=△W(2.5+0.35W),Kpf=0.4,△W为日增重(g);W 为体重(kg)。
[1] 张丽英.饲料分析及饲料质量检测技术[M].北京:中国农业大学出版社,2007.
[2] 杨嘉实,冯仰廉.畜禽能量代谢[M].北京:中国农业出版社,2004.
[3] 韩兴泰.影响反刍动物绝食代谢的因素[J].青海畜牧兽医,1990(4):31-34.
[4] 蔡建森,刁其玉.舍饲肉羊的营养需要量[C].第三届中国羊业发展大会会议论文集,北京:中国农业科技出版社,2006.
[5] 杨维仁,杨在宾,李凤双,等.大尾寒羊能量和蛋白质需要量及代谢规律的研究[J].中国养羊,1997(1):26-27.
[6] 杨在宾,杨维仁,张崇玉,等.小尾寒羊和大尾寒羊能量与蛋白质代谢规律研究[J].中国草食动物,2004,24(5):11-13.
[7] 赵玉民.中国美利奴羊育成母羊能量和蛋白质需要研究[D].延边大学,2005.
[8] 杨在宾,李凤双,杨维仁.青山羊泌乳期母羊的能量需要量研究[J].畜牧兽医学报,1993,24(5):391-398.
[9] 杨诗兴,彭大惠,张文远,等.湖羊能量与蛋白质需要量的研究[J].中国农业科学,1988,2(2):73-80.
[10] 赵新全,皮南林.藏系绵羊维持代谢能(MEm)测定[C].高寒草甸生态系统国际学术讨论会论文集,1986.
[11] Williams C B,Jenkins T G.Adynamic model of metabolizable energy utilization in growing and mature cattleⅢ.Model evaluation[J].Journal of Animal science,2003,81(6):1 390-1 398.
[12] Ludden P A,Wechter T L,Hess B W.Effects of oscillating die-tary protein on nutrient digestibility,nitrogen metabolism,gastrointestinal organ mass in sheep[J].Journal of Animal science,2002,80(11):3 021-3 026.
[13] Swanson K C,Freetly H C,Ferrell C L.Nitrogen balance in lambs fed low-quality brome hay and infused with differing proportions of casein in the rumen and abomasums[J].Journal of Animal science,2004,82(2):502-507.
[14] Ferrell C L,Kreikemeier K K,Freetly H C.The effect of supplemental energy,nitrogen,and protein on feed intake,digestibility and nitrogen flux across the gut and liver in sheep fed low-quality forage[J].Journal of Animal science,1999,77(12):3 353-3364.
猜你喜欢
现代畜牧科技(2021年10期)2021-11-19 08:42:16
中国饲料(2021年17期)2021-11-02 08:15:18
中国饲料(2020年16期)2020-11-06 06:57:54
湖南饲料(2019年4期)2019-10-17 02:05:30
动物营养学报(2018年7期)2018-07-16 03:05:30
饲料博览(2016年6期)2016-04-05 14:11:14
中国畜牧兽医文摘(2015年9期)2015-12-29 03:38:14
动物营养学报(2015年10期)2015-12-01 03:30:54
动物营养学报(2015年10期)2015-12-01 02:26:22
饲料博览(2012年8期)2012-04-13 09:27:10
