
钙对干旱胁迫下草莓SOD、CAT、 PPO活性的影响
2009-12-02 03:28湖南农业大学东方科技学院湖南长沙410128
长江大学学报(自科版) 2009年2期
彭 淼 (湖南农业大学东方科技学院,湖南 长沙 410128)
钟晓红 (湖南农业大学园艺园林学院, 湖南 长沙 410128)
张 玲 (湖南省临武县农业局,湖南 临武 424300)
杨 塞 (湖南农业大学东方科技学院,湖南 长沙 410128)
钙对干旱胁迫下草莓SOD、CAT、 PPO活性的影响
彭 淼 (湖南农业大学东方科技学院,湖南 长沙 410128)
钟晓红 (湖南农业大学园艺园林学院, 湖南 长沙 410128)
张 玲 (湖南省临武县农业局,湖南 临武 424300)
杨 塞 (湖南农业大学东方科技学院,湖南 长沙 410128)
采用自然干旱胁迫的方法,以‘章姬’、‘丰香’为材料,研究了干旱胁迫下不同Ca2+浓度处理对草莓(FragariaAnanassaDuchesne)叶片超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、多酚氧化酶(PPO)的影响。结果表明,在干旱胁迫下,外源钙能在一定程度上维持酶的活性,以减轻活性氧对草莓的伤害,从而在一定程度上提高草莓抗旱性,且不同处理之间存在差异,以10 mmol/L CaCl2处理效果最好。
草莓(FragariaAnanassaDuchesne);干旱胁迫;钙;超氧化物歧化酶;过氧化氢酶;多酚氧化酶
草莓(FragariaAnanassaDuchesne)为多年生温带草本果树,果实柔软多汁、酸甜适度、营养丰富、外观美丽、香气浓郁,在国际水果市场一直备受青睐,有“水果皇后”之誉[1]。但是由于草莓根系分布浅、叶面积较大、蒸腾旺盛,容易造成植株水分亏缺而影响生长,给草莓产量、品质带来严重影响[2],特别是在我国南方,高温往往伴随着干旱,草莓的越夏也就成了生产中的难题。为了提高草莓的抗旱性,前人做过一些研究[3~5],但用施钙来提高草莓抗旱性的研究还未见报道。自从CaCl2作为抗蒸腾剂应用于农业生产实践以后,钙与植物抗旱性的关系越来越被研究者重视。众所周知,钙是构成细胞壁的一种大量元素,也是偶联胞外信号与胞内生理反应的第二信使。环境刺激通过信号转导影响植物的生长发育,已知启动Ca2+信使系统的一个中心环节是胞质中Ca2+浓度的改变,而维持细胞内Ca2+的低稳态水平是完成Ca2+的信使作用和细胞调节功能的前提条件[6]。20世纪80年代,Mukherjee等[7]揭开了钙在提高植物抗旱性及其他抗逆性研究的序幕。生理学研究发现,适宜浓度的外源钙能Ca2+能提高苜蓿[8]、金钗石斛[9]、裸大麦[10]、芒果[11]等植物的抗旱性。本研究主要探讨在干旱胁迫下,外源钙对草莓酶促保护系统的影响及与抗旱性的关系。
1 材料与方法
1.1 试验材料
供试材料取自湖南省农业科学院草莓基地,品种为‘章姬’和‘丰香’。2005年9月定植于25 × 30 cm 白色营养钵中,培养基质为1∶1混合的珍珠岩和河沙,每盆栽种1株,置于湖南农业大学园艺园林学院蔬菜基地塑料大棚内(避雨),定期浇灌营养液,营养液参照李富恒等[12]的方法配制。前期进行正常栽培管理。
1.2 试验方法
2006年5月18日进行,选取大小、长势一致的植株60盆,按照完全随机设计方法,每个单株作为1个处理,每处理2盆,设3次重复。叶片处理参照张建霞等[13]的方法加以修改,设4个处理,分别采用CaCl2(5、10、20、30 mmol/L,另加0.5%吐温-20)喷施,对照用双蒸水,于每天下午5时喷洒叶片,以叶片湿润而不滴水为度,连续喷洒3 d,然后浇足水,第4天开始进行干旱胁迫处理,干旱胁迫采用不浇水的自然方式处理。分别于胁迫后第1、4、7、10天取心叶外侧第3~5片功能叶进行指标测定。超氧化物歧化酶活性(SOD)和过氧化氢酶活性(CAT)采用王学奎的方法[14]。SOD活性单位以抑制氮蓝四唑(NBT)光化还原50%为1个酶活性单位(U),CAT活性单位以每分钟光密度下降0.1为1个酶活性单位。多酚氧化酶活性(PPO)测定采用邻苯二酚法[15],以每克鲜重吸光密值增加0.01为1个酶活性单位。
所得数据采用DPS3.0软件中的邓肯氏新复极差法进行统计分析。
2 结果与分析
2.1 干旱胁迫下不同浓度钙处理对草莓SOD活性的影响
从表1可以看出,随着干旱胁迫时间的延长,不同浓度钙处理与对照草莓叶片SOD活性均呈现先上升后下降的趋势。在干旱胁迫第1天,SOD活性均维持较低水平。在干旱胁迫第4天,各处理SOD活性开始上升,但是‘章姬’钙处理组与对照无显著差异,而‘丰香’各处理之间,以30 mmol /L CaCl2处理活性最低,甚至低于对照。干旱胁迫第7天,随着胁迫的进一步加剧,所有处理的SOD活性都达到高峰,20 mmol/L和30 mmol/L CaCl2处理‘章姬’与对照和其他处理相比均达到显著性差异,而‘丰香’则以10 mmol/L CaCl2处理活性最高,与对照相比达到了极显著性差异。到了干旱胁迫第10天,叶片抗氧化能力降低,质膜结构破坏严重,SOD活性不能很好的维持,开始下降,‘章姬’CaCl2处理组都高于对照,以10 mmol/L CaCl2处理SOD活性最高,与对照相比达到了极显著性差异;钙处理‘丰香’草莓组也高于对照,其中也以10 mmol/L CaCl2效果最好,与对照也达到了显著性差异。
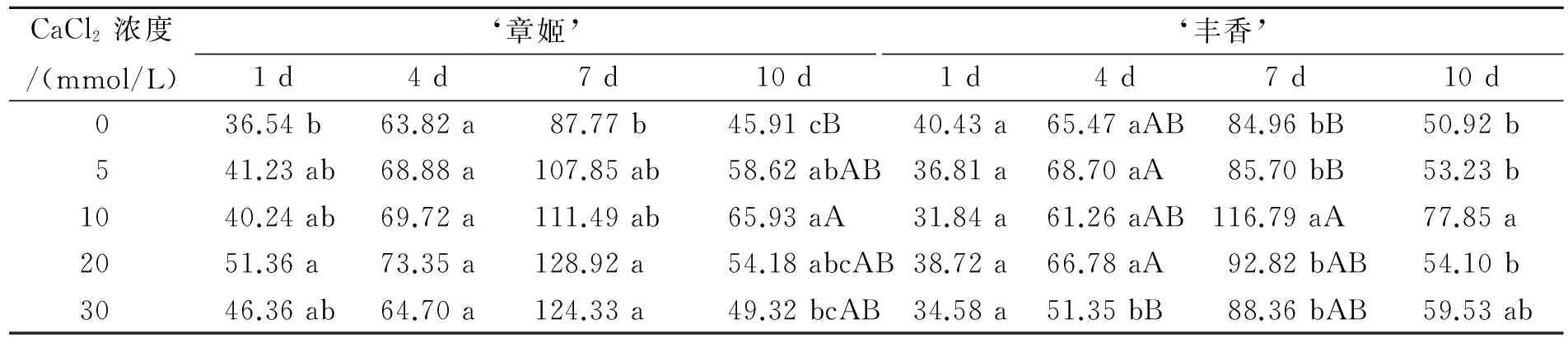
表1 不同Ca2+浓度下草莓的SOD活性比较Table 1 Comparison of SOD activity of strawberry under different concentrations of Ca2+ U/g·min-1
注:同列不同小写字母表示组间差异显著(Plt;0.05),不同大写字母表示组间极显著(Plt;0.01)。表2同。
2.2 干旱胁迫下不同浓度钙处理对草莓CAT活性的影响
由表2可知,CAT活性也呈现出与SOD活性变化一致的趋势,即先升后降,在第7天达到高峰,随后下降。在干旱胁迫第4天,‘章姬’草莓叶片CAT活性较第1天上升幅度较小,而‘丰香’则上升幅度较大,这也就是‘丰香’较‘章姬’抗旱的缘故。在干旱胁迫第7天,钙处理组均高于对照,10 mmol/L和30 mmol/L CaCl2处理‘章姬’与对照相比达到了极显著差异;除5 mmol/L CaCl2处理外,其余钙处理‘丰香’与对照达到了极显著差异。到了干旱胁迫第10天,所有钙处理‘章姬’与对照相比都达到了显著性差异,仍然维持着较高的水平,10 mmol/L和30 mmol/L CaCl2处理与对照还达到了极显著性差异;除5 mmol/L CaCl2处理外,其余钙处理‘丰香’与对照相比都达到了显著差异,10 mmol/L CaCl2处理还达到了极显著差异。结果表明,5~30 mmol/L CaCl2处理皆能在一定程度上提高草莓的CAT活性,减轻自由基对细胞膜的损害,其中以10 mmol/L CaCl2处理效果最佳。
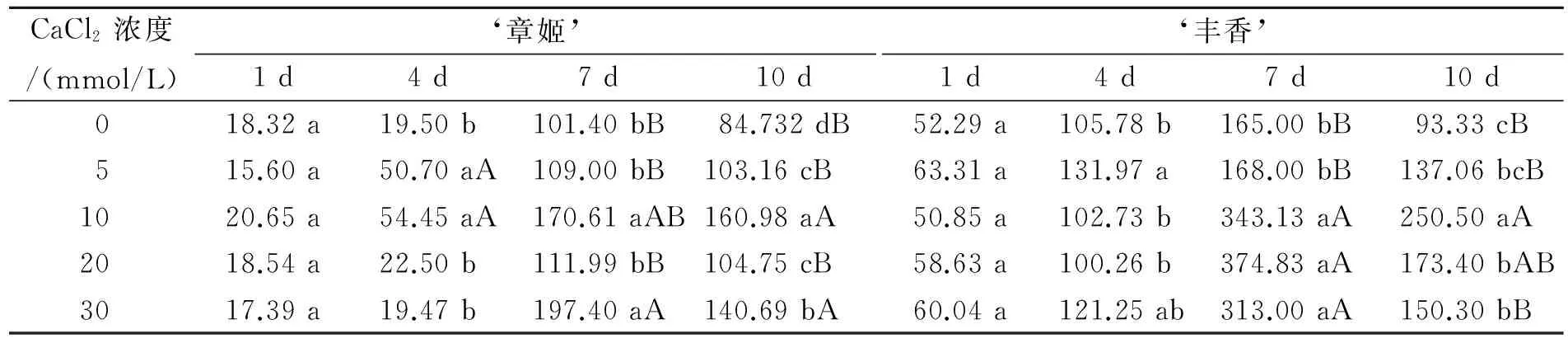
表2 不同Ca2+浓度下草莓的CAT活性比较Table 2 Comparison of CAT activity of strawberry under different concentrations of Ca2+ U/g·min-1
2.3 干旱胁迫下不同浓度钙处理对草莓PPO活性的影响
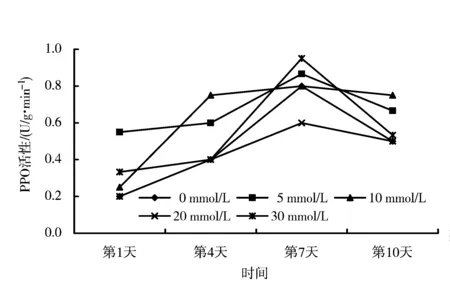
图1 不同干早胁迫程度下钙对‘章姬’草莓PPO活性的影响Figure 1 Effects of calcium on PPO activity of ‘Zhangji’ under drought stress
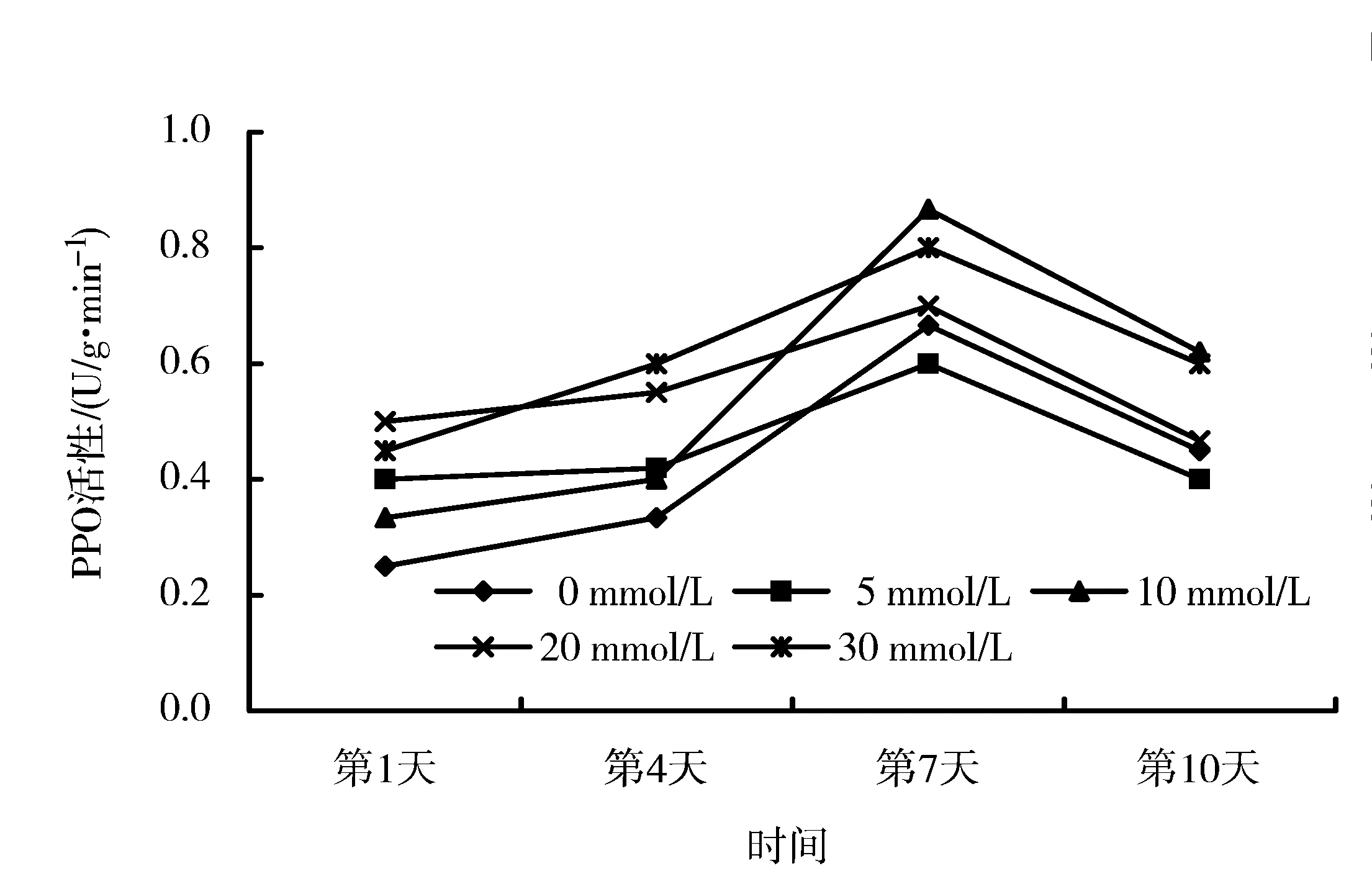
图2 不同干早胁迫程度下钙对‘丰香’PPO活性的影响Figure 2 Effects of calcium on PPO activity of ‘Fengxiang’ under drought stress
从图1、2可以看出,PPO活性与SOD、CAT的变化趋势一致,也是随着胁迫时间的延长先上升后下降,在第7天达到高峰,然后下降。从‘章姬’的变化曲线来看,以10 mmol/L CaCl2处理和30 mmol/L CaCl2处理变化最为剧烈,其中前者在胁迫第4天就达到了较高值,之后变化平稳,维持了较高活性,后者尽管在第7天达到高峰值,但在第10天却急剧下降。在第10天,除20 mmol/L CaCl2处理外均高于对照。‘丰香’草莓变化曲线也以10 mmol/L CaCl2处理变化剧烈,在第7天和第10天,活性均高于其他处理。与‘章姬’相似,20 mmol/L CaCl2处理‘丰香’效果也不佳。
3 讨论
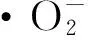
本研究结果表明,草莓叶片中SOD、CAT、PPO活性在干旱胁迫前期(第1~7天)呈上升趋势,在干旱胁迫第4天,各处理SOD活性开始上升,但是‘章姬’钙处理组与对照无显著差异,而‘丰香’各处理之间,以30 mmol/LCaCl2处理活性最低,甚至低于对照,这可能是因为对照未经钙处理,细胞膜受害程度较大,进而启动植物防御反应,SOD活性被诱导的缘故。到了胁迫第10天,叶片抗氧化能力的进一步降低,叶片遭受强烈的氧化伤害,膜脂过氧化加剧,3种酶的活性均表现出一致的下降趋势,即便是钙处理的草莓叶片也不能维持之前的较高酶活性,但通过钙处理后的草莓叶片酶活性要高于对照,这表明钙能有效诱导保护酶的活性,并有效调节好保护酶的活性,有效清除膜脂过氧化产生的活性氧,降低膜脂过氧化程度,维持膜的相对稳定性,恢复或维持正常的代谢水平,一定程度上减轻了干旱对植物的伤害。由于Ca2+对膜保护作用具有浓度效应,如果Ca2+长期以较高的浓度存在于细胞质和细胞核中,不能及时泵回到钙库或者胞外,就可能引起植物的“钙中毒”,它将会同磷酸反应生成沉淀而干扰以磷酸为基础的能量代谢。因此,如果不存在毒害作用,低浓度的Ca2+就能满足细胞的生理反应[19~21]。本研究中,不论是‘章姬’还是‘丰香’,都以10 mmol/L CaCl2处理效果最佳,与吴显芝[9]、吴德宽[10]等的研究结果一致。这充分说明,只有一定浓度的外源钙才能在一定程度上提高草莓抗旱性。同时,由于在干旱胁迫下,植物对Ca2+的吸收能力显著降低造成植物体内Ca2+缺乏[22],因此喷施CaCl2既可以作为施肥措施来满足草莓全面营养生长需要,又不失为一种经济简便的方法来减轻干旱胁迫给草莓带来的伤害,从而提高草莓的抗旱性。
[1]谭昌华,代汉平,雷家军.世界草莓生产与贸易现状及发展趋势(下)[J].世界农业,2003,(6):16~19.
[2]刘明池,小岛孝之,田中宗浩,等.亏缺灌溉对草莓生长和果实品质的影响[J].园艺学报,2001,28(4):307~311.
[3]吴少华.BR和KT对草莓抗旱性的影响[J].福建农业学报,2001,16(2):56~58.
[4]黄建昌,肖 艳.PP333提高草莓抗旱性研究[J].仲恺丧业技术学院学报,1996,9(1):67~72.
[5]黄建昌,肖 艳.自由基清除剂对草莓水分胁迫的保护作用[J].仲恺农业技术学院学报,1995,8(2):63~67.
[6]Poovaiah B W,Reddy A S N. Calcium and signal transduction in plants[J]. Critical Review of Plant Science,1993,12:185~221.
[7]Mukherjee S P,Choudhwri M A. Effect of water stress on some oxidative enzymes and senesence in vigna seedlings[J]. Physiol Plant,1981,52:37~42.
[8]姜义宝,李建华,方丽云,等.钙处理对苜蓿幼苗抗旱性的影响[J].草业学报,2008,30(1):117~120.
[9]吴显芝,乙 引.外源Ca2+对干旱胁迫下金钗石斛幼苗生长的影响[J].贵州师范大学学报(自然科学版),2006,24(2):23~26.
[10]吴德宽,吴渤海.干旱胁迫下钙对裸大麦叶片生理生化特性的影响[J].麦类作物,1997,17(2):42~44.
[11]贾虎森,蔡世英,李德全,等.土壤干旱胁迫下钙处理对芒果幼苗光合作用的影响[J].果树科学,2000,17(1):52~56.
[12]李富恒,王 艳.草莓无土栽培营养液的配制及管理[J].农业系统科学与综合研究,2001,17(3):210~212.
[13]张建霞,李新国,孙中海.外源钙对柑橘抗热性的相关生理生化指标的影响[J].华中农业大学学报(自然科学版),2005,24(4):397~400.
[14]王学奎.植物生理生化实验原理和技术(第2版)[M].北京:高等教育出版社,2006.169~173.
[15]王海波,李 疆,王孝娣,等.短时间高温处理下桃芽酚类物质及相关酶活性与体眠解除的关系[J].果树学报,2006,23(3):365~369.
[16]Giannopolitis C N,Ries S K. Superoxide dismutase II. Purification and quantitative relationship with water soluble protein in seedlings[J]. Plant Physiol,1977,59:315.
[17]段云青,王 艳,雷焕贵.镉胁迫对小白菜POD、PPO 和SOD 活性的影响[J].河南农业科学,2006,(7):88~91.
[18]袁清昌.钙提高植物抗旱能力的研究进展[J].山东农业大学学报,1999,30(3):302~306.
[19]Loneragan J F,Snowball K,Simmons W J. Response of plants to calcium concentration in solution culture[J]. Aust J Agric Res,1968,19:845~857.
[20]Minorsky P V. A heuristic hypothesis of chiiline injury on plants. A role for calcium as the primary physiological transducer of injury[J]. Plant Cell Environ,1985,8:75~83.
[21]龚 明,李 英,曹宗巽. 植物体内的钙信使系统[J].植物学通报,1990,7(3):19~29.
[22]缪 颖,曹家树,曾广文.渗透胁迫下大白菜吸收45Ca2+的变化[J].浙江农业大学学报,1998,24(2):126~130.
2008-09-04
彭 淼(1981-),男,湖南汨罗人,农学硕士,助教,研究方向为果树栽培生理与生态.
钟晓红,E-mail: xh-zhong@163.com.
10.3969/j.issn.1673-1409(S).2009.01.004
Q945.78
A
1673-1409(2009)01-S011-04
猜你喜欢
疯狂英语·新读写(2022年7期)2022-11-22
湘潮(上半月)(2019年6期)2019-05-22
音乐教育与创作(2019年9期)2019-05-16
阅读与作文(小学高年级版)(2017年10期)2017-10-11
浙江农业学报(2017年1期)2017-05-17
西南农业学报(2016年4期)2016-05-17
海峡影艺(2012年1期)2012-11-30
草食家畜(2012年2期)2012-03-20
现代农业科技(2009年17期)2009-03-08
