
生物质炭对镉铅污染土壤线虫群落的影响
2025-02-19 00:00:00马振华时倩茹宁欣杰魏宏杨王璨张静静张彪杨素勤
中国农业科技导报 2025年2期


摘要:为研究连续施用改性生物质炭对农田镉铅污染土壤修复及线虫群落的影响,以改性生物质炭为材料,开展连施改性生物质炭2、3、4和5年的定位试验。通过测定小麦籽粒和土壤有效态镉铅含量,探究改性生物质炭持续钝化土壤镉铅的能力,并利用高通量测序技术,探究改性生物质炭对土壤线虫群落的影响。结果表明,改性生物质炭能显著降低小麦籽粒和土壤有效态镉铅含量,其中,小麦籽粒镉含量降幅为21%~35%,小麦籽粒铅含量降幅为27%~39%;土壤有效态镉铅含量降幅分别为21%~43%和15%~28%。施用改性生物质炭降低食真菌土壤线虫滑刃属(Aphelenchoides)丰度,而植物寄生性土壤线虫默林属(Merlinius)、针属(Paratylenchus)等占比均增至40%以上。土壤中线虫c-p2类群的比例大幅降低,而c-p3类群占比增加;自由生活线虫成熟指数减少,植物寄生类线虫成熟指数增加,表明不同营养类群、不同生活史的土壤线虫对重金属污染的反应特性不同。整体而言,连施不同年限处理间线虫群落变化不明显。以上表明,小麦籽粒镉含量在连续施用改性生物质炭4年达到最低,小麦籽粒铅含量施用改性生物质炭2年即可低于GB 2762—2017限值0.2 mg·kg-1。施入改性生物质炭可调节线虫群落结构,从而影响土壤环境,在重金属污染土壤综合利用与修复方面具有较强的应用价值。
关键词:改性生物质炭;土壤重金属;土壤线虫;高通量测序doi:10.13304/j.nykjdb.2023.0321
中图分类号:S154 文献标志码:A 文章编号:1008‐0864(2025)02‐0201‐10
镉和铅在土壤中具有较强的隐蔽性,易累积于植物体内影响作物生长,并通过食物链威胁人类健康[1]。生物质炭是在限氧或缺氧条件下生物质热裂解得到的产物,具有比表面积大、孔隙结构发达、功能团丰富等特点[2],在土壤改良[3]、重金属钝化[4]等方面具有广阔的应用前景。近年来,研究者发现,生物质炭可在短时间内钝化土壤中的镉铅[5],但植株中镉含量的降幅因施炭量不同而有所变化[6]。土壤pH和阳离子交换量(cation exchangecapacity,CEC)在连施生物质炭3年后依旧显著提高,但其效应存在一定的滞后性[7]。因此,研究施用不同年限生物质炭对污染土壤镉铅钝化效果具有较强的实践价值。此外,长期施用生物质炭可调节污染土壤微生物群落结构,提升微生物多样性。杨素勤等[8]研究发现,改性生物质炭能调节土壤细菌和真菌群落结构,对土壤质量有明显影响。
线虫相比于其他微生物具有独特的土壤生态功能,不同种群的土壤线虫在食物网中的营养级不同[9],且具备分布广、数量多、世代周期短等优点,常作为指示土壤健康状况的优选生物[10]。Zhang等[11]研究显示,生物质炭增加食真菌线虫比例的同时减少了植物寄生性线虫的比例。杨贝贝等[12]发现,添加生物质炭增加了土壤中食细真菌线虫数量,降低了植物寄生性线虫比例。然而,就生物质炭在未受污染区对线虫群落的影响而言,相关研究结果并不完全一致。牛亚茹[13]研究表明,食细真菌线虫占比随生物质炭的施入而降低,但植物寄生性线虫的占比无明显变化。Domene等[14]发现,施用较低水平的生物质炭可促进植物寄生性线虫的增加。由此判断,生物质炭、土壤和线虫之间存在复杂的互作关系,探究施用生物质炭对土壤线虫群落结构的改变具有重要意义。
线虫群落结构的变化也可较灵敏地响应土壤污染状况。Renčo等[15]研究表明,线虫的丰度、分类群数量以及线虫群落的营养复杂性随土壤污染程度的加深显著降低;邓普荣等[16]分析3种修复措施对镉污染水稻农田线虫群落的影响发现,土壤线虫的成熟指数等生活史策略指数存在显著差异。由此可见,研究施用不同年限生物质炭对污染土壤线虫群落结构的影响可为阐明生物质炭长期施用下污染土壤生态特征和功能变化提供参考依据。本研究以磷酸改性稻壳生物质炭为材料,设置连施2、3、4和5年的田间定位试验,探讨改性生物质炭对污染土壤镉铅的钝化效果及小麦籽粒镉铅积累水平,同时借助高通量测序技术,探究改性生物质炭对土壤线虫群落结构的影响,为改性生物质炭长期施用条件下污染土壤生态特征和功能变化提供科学依据。
1 材料与方法
1.1 试验材料
试验材料为改性稻壳生物质炭,购于郑州博奥农业科技有限公司,经磷酸改性pH为5.5,镉和铅含量分别为0.08 和1.75 mg·kg-1,比表面积为76.6 m2·g-1,施用量11.19 t·hm-2·a-1。
1.2 试验设计
试验地点位于豫北某冶炼厂周边的污染农田。土壤类型为褐土,耕层土壤pH 7.71,镉含量达2.34 mg·kg-1,铅含量为181.42 mg·kg-1,参照GB 15618—2018[17],属安全利用类耕地。
试验于2015—2019年进行,以未施加改性生物质炭为对照区(CK),设置改性生物质炭连施2(Y15-16)、3(Y15-17)、4(Y15-18)和5年(Y15-19)后停施,共计5 个处理,每个处理重复3 次,各处理间均设置50 cm的保护行。
1.3 样品采集及指标测定
在2022年小麦成熟季,采集点对点土壤−小麦样品。小麦籽粒洗净、烘干后粉碎过筛,用于测定小麦籽粒中的镉铅含量。取耕作层土壤(0—20 cm),混匀后部分土样冷冻暂存,用于分离线虫;其余土样风干、研磨过筛,用于测定土壤理化指标。
1.3.1 土壤pH 的测定 按照水土比2.5∶1.0,采用pH计测定[18]。
1.3.2 镉铅含量的测定 土壤镉铅含量通过X射线荧光重金属分析仪(E-max 700 Plus,佳谱科技)测定,选用土壤标准物质GBW07423、GBW07426、GBW07385 进行质量控制,准确度92%~105%,精密度1%~11%;土壤有效态镉铅含量参照GB/T 23739—2009[19],通过二乙三胺五乙酸(diethylenetriaminepentaacetic acid,DTPA)提取剂浸提,采用X射线荧光重金属分析仪测定;小麦籽粒中的镉铅含量通过HNO3-HClO4法消解,采用石墨炉原子吸收分光光度法测定[20]。
1.3.3 土壤线虫高通量测序 PCR 扩增引物采用NF1F(5’-GGTGGTGCATGGCCGTTCTTAGTT-3’)和18Sr2bR(5’-TACAAAGGGCAGGGACGTAAT-3’),每个样本3个重复,待PCR产物检测定量后利用Illumina Miseq测序仪进行测序(上海美吉生物医药科技有限公司)。土壤线虫高通量测序数据利用Uparse(version 7.0)进行OTU(operational taxonomicunits)分析。
1.4 数据处理
小麦、土壤样品数据采用SPSS 25.0、Origin2022进行统计、分析和绘图,采用Duncan法检验各处理间的差异(Plt;0.05)。土壤线虫样品数据采用生物云平台分析制图。
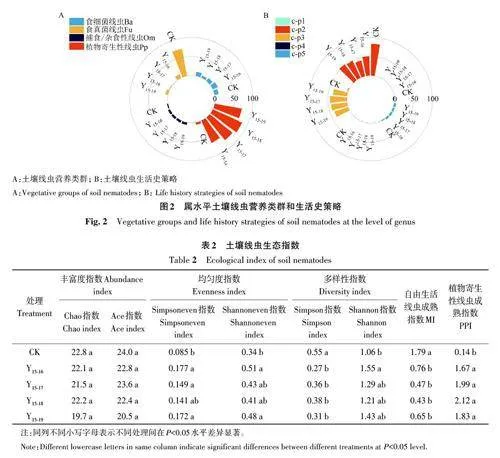
土壤线虫多样性生态指数采用线虫营养类群多样性和生活史多样性[21]。营养类群多样性是根据线虫的食性特征划分为食细菌线虫(becterivores, Ba)、食真菌线虫(fungivores, Fu)、植物寄生性线虫(plant-parasites, Pp)和捕食/杂食性线虫(predators/omnivores, Om)[22]。生活史多样性是根据线虫生活史策略不同,划分5 个c-p(colonizer persister)类群,c-p值越小代表世代繁殖时间越短、产卵量越大、代谢越快、耐环境压力越强[23]。
土壤线虫生态指数通过丰富度指数(Chao指数、Ace 指数)、均匀度指数(Simpsoneven 指数、Shannoneven 指数)、多样性指数(Simpson 指数、Shannon 指数)、自由生活线虫成熟指数(maturityindex of free-living,MI)和植物寄生性线虫成熟指数(maturity index of plant parasitic,PPI) 进行表征,并利用距离矩阵进行土壤线虫群落主坐标分析(principal coordinate analysis,PcoA)。
2 结果与分析
2.1 小麦籽粒及土壤有效态镉铅含量分析
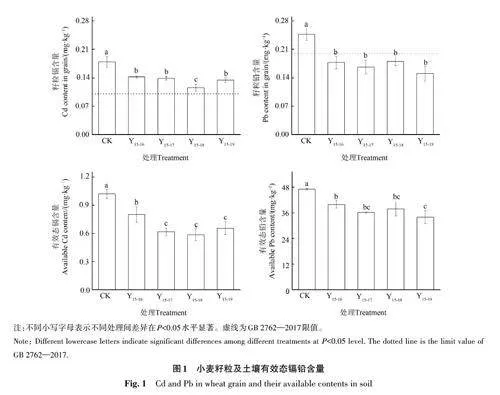
由图1可知,施用不同年限改性生物质炭处理的小麦籽粒镉铅含量均显著低于对照,小麦籽粒镉含量降幅为21%~35%。其中,连施改性生物质炭4年后(Y15-18),小麦籽粒中的镉含量最低,相比于对照降幅为35%,且这一处理与其他不同年限(Y15-16,Y15-17,Y15-19)处理相比,差异显著(Plt;0.05)。小麦籽粒铅含量随改性生物质炭施用年限不同均有不同程度降低,降幅为27%~39%,且均低于GB2762—2017[24]规定的限值(0.2 mg·kg-1),但施用改性生物质炭的处理间差异不显著。
土壤有效态镉铅含量在施用改性生物质炭处理后均显著低于对照,土壤有效态镉铅含量降幅分别为21%~43% 和15%~28%,且随改性生物质炭施用年限增加略有降低。就土壤有效态镉而言,相比于2年连施(Y15-16)处理,连施3年(Y15-17)处理显著降低,但继续施加改性生物质炭(Y15-18、Y15-19)其有效态镉含量无显著降低。土壤有效态铅含量在连施改性生物质炭5年后(Y15-19)达到最低值,为33.87 mg·kg-1,相比于对照,降幅达28%。然而,连施3年以上的3个处理(Y15-17、Y15-18、Y15-19)间土壤有效态铅含量差异不显著。
2.2 土壤线虫群落组成分析
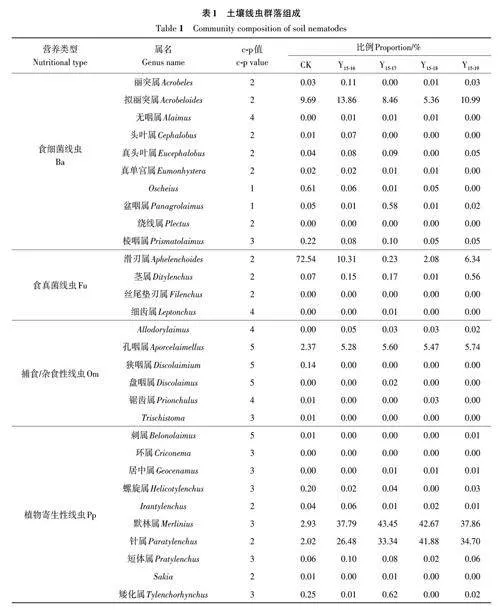
该研究检测到的OTU 分属于3 纲、8 目、26科、46 属、61 种。其中,Y15-16 处理OTU 数最多;OTU数量最少的是Y15-18 处理。属水平主要土壤线虫群落组成见表1,滑刃属(Aphelenchoides)线虫在对照区占比最高,达72.54%,施加改性生物质炭后大幅降低,在Y15-17 处理中最低,仅为0.23%。默林属(Merlinius)和针属(Paratylenchus)线虫在施用改性生物质炭的处理中占绝对优势,其中默林属(Merlinius)在Y15-17 处理中占比最高,达43.45%,针属(Paratylenchus)在Y15-18 处理中占比最高,达41.88%。
在土壤线虫营养类群方面(图2A),食细菌线虫(Ba)在各处理间差异不明显。但对照区的食真菌土壤线虫(Fu)占比较高,随改性生物质炭的施入这一比例大幅降低,在Y15-17 处理达到最低值。未施加改性生物质炭的污染土壤中(对照区)植物寄生性线虫(Pp)群落受明显抑制,占比仅为5.52%,随改性生物质炭的施用植物寄生性线虫群落占比有不同程度的提升,在Y15-18处理占比最高,达84.60%。捕食/杂食类土壤线虫(Om)随改性生物质炭施入有所提高,但提升幅度有限。
就线虫生活史策略而言(图2B),无论施加改性生物质炭与否,土壤线虫群落以c-p2和c-p3类群为主,二者之和高于86%。对照区土壤线虫c-p2群落占比最高,达84.47%,随改性生物质炭施入这一比例大幅下降;与c-p2变化状况相反,土壤线虫c-p3类群在对照区中仅为3.67%,改性生物质炭施入后这一比值迅速升高,说明线虫生活史策略对环境变化较敏感,改性生物质炭的施入可使线虫生活史由r-策略者向k-策略者转移,土壤生态环境状况趋于好转。土壤线虫c-p5类群占比状况与营养类群中的捕食/杂食类相似。土壤线虫c-p1和c-p4类群占比均低于1%。
2.3 土壤线虫生态指数
土壤线虫生态指数见表2,相较于对照区,施入改性生物质炭后土壤线虫Simpson多样性指数显著降低,Shannon 多样性指数、Simpsoneven 和Shannoneven均匀度指数均有不同程度升高,说明本研究中的改性生物质炭可提高土壤线虫群落的多样性和均匀度,但连施不同年限改性生物质炭处理间差异不明显。
土壤中自由生活线虫成熟指数(MI)在对照区较高,达1.79,随改性生物质炭施入MI显著降低,降幅在57%~76%。植物寄生性线虫成熟指数(PPI)在未施加改性生物质炭的污染土壤中(对照区)仅为0.14,施入改性生物质炭后显著增加,相较于MI指数其变幅较大,是对照区的12~15倍。由以上可知,镉铅复合污染土壤在改性生物质炭施入后,自由生活线虫和植物寄生性线虫等不同生活史的土壤线虫反应各异,土壤线虫群落结构出现不同程度的改变。
2.4 土壤线虫群落主坐标分析
土壤线虫群落主坐标分析(PcoA)见图3,2个主坐标轴分别解释74.59%和8.36%的变异信息。施用不同年限改性生物质炭的处理间土壤线虫群落结构较相似,且均与对照有明显不同。施用改性生物质炭的处理间,主成分1(principalcomponent 1,PC1)显著分离了Y15-16 与Y15-18 处理,说明Y15-16 处理的土壤线虫群落组成与Y15-18 处理具有显著差异。
2.5 环境因子对土壤线虫群落的影响
对丰度前30的线虫属与环境因子进行冗余分析( redundancy analysis,RDA)结果如图4所示,土壤线虫群落的相对丰度在第1主轴(X 轴)方差解释度为70.56%,在第2主轴(Y 轴)为2.11%,总的方差解释度达72.67%。其主要的影响因子集中于第1主轴的土壤有效态镉、有效态铅以及土壤pH。其中,这些主要影响因子与滑刃属(Aphelenchoides)呈正相关,与默林属(Merlinius)和针属(Paratylenchus)呈明显负相关。
属水平土壤线虫与环境因子相关性分析如图5所示,环境因子中的土壤pH、土壤有效态镉、土壤有效态铅含量均与滑刃属、棱咽属(Prismatolaimus)、螺旋属(Helicotylenchus)呈极显著正相关,与默林属、针属呈极显著负相关,与Pristionchus、狭咽属(Discolaimium)呈显著正相关;而土壤总镉含量只与棱咽属呈正相关。在上述土壤线虫中,棱咽属和螺旋属对环境因子响应较为一致,Pristionchus 和狭咽属以及默林属和针属也有相似结果。此外,土壤有效态镉、有效态铅相较于土壤总镉、总铅含量对线虫影响较大,整体表现为土壤有效态镉gt;土壤有效态铅gt;土壤总镉gt;土壤总铅。
3 讨论
重金属由土壤转运至作物可食用部位受多个环节控制,如重金属在土壤中的存在状态[25]、重金属被吸收后存储于根细胞液泡中[26]、作物茎叶部位对重金属的阻隔[27]等。本研究采用的生物质炭经磷酸改性后,比表面积显著增大,更利于稳定土壤中的镉铅离子,降低土壤有效态镉铅含量;磷酸改性生物质炭施用后土壤中有效态镉铅含量明显降低。课题组在前期研究中发现,磷酸改性生物质炭可降低土壤镉铅的弱酸提取态、还原态,增加残渣态,且随施用量增加更明显[28];同时碳酸盐结合态镉较高比例的降低,也有PO34-与重金属离子生成沉淀的参与[8]。土壤重金属形态的变化可能与生物质炭中磷素的存在有一定关系。磷酸二氢钾施入土壤后X射线衍射分析显现新的衍射峰,经图谱物相鉴定为重金属−磷化合物的特征峰[29]。付煜恒等[30]分析磷酸盐对镉铅复合污染土壤的钝化效果,也证实这一效果的主要原因是难溶性羟基金属矿和磷酸盐沉淀的形成。
本研究中,施用改性生物质炭后小麦籽粒中的镉铅含量显著降低,其中小麦籽粒中的铅含量在施用改性生物质炭后均符合GB 2762—2017[24]的规定,镉含量略高于GB 2762—2017[24]的限值0.1 mg·kg-1,表明本研究所施用的改性生物质炭在土壤钝化重金属方面效果显著。此外,连施不同年限改性生物质炭的处理间,小麦籽粒镉铅含量也有不同。
土壤生物群落特征的变化常用于反映土壤的健康水平[31]。其中,土壤小型动物主要是捕食、食腐、食真菌或食细菌的动物,包括线虫等。线虫是土壤动物的优势类群,在食物网中的营养级有所不同,可以综合反映土壤环境的物理、化学和生物学特性的细微变化[32‐33]。根据土壤线虫对重金属的响应状况,线虫群落可分为极度敏感型、中度敏感型和不敏感型[34]。食细菌和食真菌线虫对污染环境具有一定耐受性。此外,它们可以驱动并加速土壤氮的矿化和硝化过程,从而促进作物对氮的吸收和生长[35‐36]。王赢利等[37]和成碧君等[38]研究发现,重金属污染越重的土壤食细真菌线虫在线虫总数中占比越高。本研究对照区的食细真菌线虫也符合这一特点。分析以上结果,其原因可能是由于在污染土壤中滑刃属对镉铅耐性较强,相比之下,该类线虫比例有所增加。研究表明,食细菌线虫拟丽突属(Acrobeloides)、食真菌线虫滑刃属、真滑刃属(Aphelenchus)对重金属污染物具有一定的耐受性[39-42]。
本研究中,施加改性生物质炭的处理植物寄生性土壤线虫占比相较于对照区变化较大。王赢利等[37]发现,随着污染指数的增加,植物寄生性线虫数量减少且PPI丰度降低,说明在重金属污染的土壤中植物寄生类线虫较为敏感。而施加生物质炭后镉铅钝化、毒性降低,植物寄生性线虫占比随之增加。研究表明,生物质炭会促进寄生性线虫的增加,如Domene等[14]发现,植物寄生性线虫与生物质炭施用量有关,低水平的施入量将促进植物寄生性线虫比例的增加,而较高水平的施入量结果与其相反。此外,植物寄生性土壤线虫以取食作物根部为生,其类群与作物根系生长状况相关。李孟洁[43]研究发现,植物寄生性土壤线虫在作物成熟期数量达到最大。与此同时,生活在土壤颗粒表面水膜中的自由生活线虫,由于受小麦生长对水分的消耗,生存环境受限在一定程度上致使其比例降低[44]。但植物寄生类土壤线虫危害作物的生长,特别是根结线虫、孢囊线虫[45]以及粒线虫,其比例增加将危害作物生长甚至影响产量,本研究中未检测到孢囊线虫、根结线虫和粒线虫。但默林属、针属占比较大,其与孢囊线虫、根结线虫等植物内寄生病原线虫不同,二者均属于外寄生类[46]土壤线虫,且在土壤中广泛存在,目前对小麦的影响尚不清楚,仍需进一步研究。
本研究中,施用改性生物质炭能明显改变土壤线虫群落,但连施改性生物质炭不同年限处理间线虫群落变化不明显。其中,环境因子中的土壤有效态镉、有效态铅、土壤pH均与螺旋属呈极显著正相关。螺旋属对较高含量铅仍具有耐受性[47]。同时这些环境因子与默林属、针属呈极显著负相关,说明污染土壤中镉铅有效态比其全量更大程度上影响土壤线虫群落结构。
参 考 文 献
[1] 黄卫, 庄荣浩, 刘辉, 等. 农田土壤镉污染现状与治理方法研
究进展[J]. 湖南师范大学自然科学学报, 2022, 45(1): 49-56.
HUANG W, ZHUANG R H, LIU H, et al .. Recent advances of
the current situation and remediation methods of cadmium
contamination in paddy soil [J]. J. Nat. Sci. Hunan Norm.
Univ., 2022, 45(1): 49-56.
[2] 杨莉, 付婧, 文子伟, 等. 6种低温生物质炭的制备及结构表
征[J]. 吉林农业大学学报, 2021, 43(5): 565-573.
YANG L, FU J, WEN Z W, et al .. Preparation and structure
characterization of six kinds of low temperature biochar [J]. J.
Jilin Agric. Univ., 2021, 43(5): 565-573.
[3] 孙宇龙, 张永利, 苏有健, 等. 生物质炭对土壤物理结构性
状和水分特征影响的研究进展[J]. 江苏农业科学, 2022,
50(23): 25-32.
SUN Y L, ZHANG Y L, SU Y J, et al .. Effects of biochar on
soil physical structure properties and hydrologic properties: a
review [J]. Jiangsu Agric. Sci., 2022, 50(23): 25-32.
[4] 杨文浩, 李佩, 周碧青, 等. 生物炭缓解污染土壤中植物的
重金属胁迫研究进展[J]. 福建农林大学学报(自然科学版),
2019, 48(6): 695-705.
YANG W H, LI P, ZHOU B Q, et al .. Biochar-mediated
alleviation of heavy metal stress in plants growing in
contaminated soils: a review [J]. J. Fujian Agric. For. Univ.
(Nat. Sci.), 2019, 48(6): 695-705.
[5] 左静, 陈德, 郭虎, 等. 小麦秸秆生物质炭对旱地土壤铅镉
有效性及小麦、玉米吸收的影响[J]. 农业环境科学学报,
2017, 36(6): 1133-1140.
ZUO J, CHEN D, GUO H, et al .. Effects of biochar on Cd Pb
availability and uptake by maize and wheat in upland soil [J].
J. Agric. Environ. Sci., 2017, 36(6): 1133-1140.
[6] 施琪, 鲁然英, 常德政, 等. 增施生物质炭对镉污染土壤的
修复效果研究[J]. 江西农业学报, 2019, 31(10): 83-87.
SHI Q, LU R Y, CHANG D Z, et al .. Effects of increasing
biochar application on remediation of Cd contaminated soil [J].
Acta Agric. Jiangxi, 2019, 31(10): 83-87.
[7] 肖和友, 朱伟, 王海军, 等. 连续施用不同生物质炭对植烟土
壤特性和烤烟品质的影响[J]. 中国烟草科学, 2021, 42(3):
19-25.
XIAO H Y, ZHU W, WANG H J, et al.. Effects of continuous
application of different biochar on tobacco-planting soil, flue-cured
tobacco leaf quality [J]. China Tob. Sci., 2021, 42(3): 19-25.
[8] 杨素勤, 魏森, 张彪, 等. 连续施用改性生物质炭对镉铅土
壤修复效果及其对微生物群落结构的影响[J]. 农业环境科
学学报, 2022, 41(7): 1460-1471.
YANG S Q, WEI S, ZHANG B, et al .. Remediation effect of
continuous application of modified biochar on cadmium-and
Lead-contaminated soil and its effect on microbial community
structure [J]. J. Agro-Environ. Sci., 2022, 41(7): 1460-1471.
[9] 杜晓芳, 李英滨, 刘芳, 等. 土壤微食物网结构与生态功
能[J]. 应用生态学报, 2018, 29(2): 403-411.
DU X F, LI Y B, LIU F, et al .. Structure and ecological
function of soil micro-food web [J]. Acta Appl. Ecol., 2018,
29(2): 403-411.
[10] LU Q F, LIU T T, WANG N Q, et al .. A review of soil
nematodes as biological indicators for the assessment of soil
health [J]. Front. Agric. Sci. Eng., 2020, 7(3): 275-281.
[11] ZHANG X K, LI Q, LIANG W J, et al .. Soil nematode response
to biochar addition in a Chinese wheat field [J]. Pedosphere,
2013, 23(1): 98-103.
[12] 杨贝贝, 朱新萍, 赵一, 等. 生物炭基肥施用对棉田土壤线
虫群落结构的影响[J]. 中国土壤与肥料, 2020(4): 66-71.
YANG B B, ZHU X P, ZHAO Y, et al .. Effect of biochar based
application on soil nematode community structure in cotton
field [J]. Soil Fert. Sci. China, 2020(4): 66-71.
[13] 牛亚茹. 施用生物质炭对日光温室黄瓜生长及土壤微生物
和线虫群落结构的影响[D]. 南京: 南京农业大学, 2016.
NIU Y R. Effect of biochar on cucumber growth and soil
microbial and nematode community structure in sunlight
greenhouse [D]. Nanjing: Nanjing Agricultural University, 2016.
[14] DOMENE X, MATTANA S, SÁNCHEZ-MORENO S. Biochar
addition rate determines contrasting shifts in soil nematode
trophic groups in outdoor mesocosms: an appraisal of underlying
mechanisms [J/OL]. Appl. Soil Ecol., 2021, 158: 103788 [2023-
03-23]. https://doi.org/10.1016/j.apsoil.2020.103788.
[15] RENČO M, ČEREVKOVÁ A, HLAVA J, et al .. Life in a
contaminated environment: how soil nematodes can indicate
long-term heavy-metal pollution [J/OL]. J. Nematol., 2022,
54(1): 20220053 [2023-03-23]. https://doi.org/10.2478/jofnem-
2022-0053.
[16] 邓普荣, 姚志, 刘勇波. 基于线虫生物多样性的稻田镉污染土
壤修复成效研究[J]. 环境科学研究, 2022, 35(10): 2378-2387.
DENG P R, YAO Z, LIU Y B. Effects of cadmium
contamination remediation on the biodiversity of soil nematodes in
paddy fields [J]. Res. Environ. Sci., 2022, 35(10): 2378-2387.
[17] 中华人民共和国生态环境部, 国家市场监督管理总局. 土
壤环境质量 农用地土壤污染风险管控标准(试行): GB
15618—2018 [S]. 北京: 中国标准出版社, 2018.
[18] 鲍士旦. 土壤农化分析[M]. 第三版. 北京: 中国农业出版社,
2000: 163-165.
[19] 刘凤枝,刘铭,蔡彦明,等. 土壤质量 有效态铅和镉的测定 原子
吸收法: GB/T 23739—2009 [S]. 北京: 中国标准出版社, 2009.
[20] 中华人民共和国国家卫生和计划生育委员会. 食品安全国
家标准 食品中镉的测定: GB 5009.15—2014 [S]. 北京: 中国
标准出版社, 2014.
[21] 李琪, 梁文举, 姜勇. 农田土壤线虫多样性研究现状及展
望[J]. 生物多样性, 2007, 15(2): 134-141.
LI Q, LIANG W J, JIANG Y. Present situation and prospect of
soil nematode diversity in farmland ecosystems [J]. Biodiversity
Sci., 2007,15(2): 134-141.
[22] YEATES G W, BONGERS T, DE GOEDE R G M, et al ..
Feeding habits in nematode families and genera:an outline for
soil ecologists [J]. J. Nematol., 1993, 25(3): 315-331.
[23] BONGERS T. The maturity index: an ecological measure of
environmental disturbance based on nematode species
composition [J]. Oecologia, 1990, 83(1): 4-19.
[24] 中华人民共和国国家卫生和计划生育委员会, 国家食品药
品监督管理总局. 食品安全国家标准 食品中污染物限量:
GB 2762—2017 [S]. 北京: 中国标准出版社, 2017.
[25] 汪鹏, 王静, 陈宏坪, 等. 我国稻田系统镉污染风险与阻
控[J]. 农业环境科学学报, 2018, 37(7): 1409-1417.
WANG P, WANG J, CHEN H P, et al .. Cadmium risk and
mitigation in paddy systems in China [J]. J. Agro-Environ. Sci.,
2018, 37(7): 1409-1417.
[26] ZHAO F J, WANG P. Arsenic and cadmium accumulation in
rice and mitigation strategies [J]. Plant Soil, 2020, 446(1): 1-21.
[27] CLEMENS S, AARTS M G M, THOMINE S, et al .. Plant
science: the key to preventing slow cadmium poisoning [J].
Trends Plant Sci.,2013, 18(2): 92-99.
[28] 周志云. 磷酸改性生物炭和氯化物混施对小麦吸收铅镉的
影响[D]. 郑州: 河南农业大学, 2018.
ZHOU Z Y. Effects of phosphoric-acid modified biochar
combined with chlorine on lead and cadmium absorption in
wheat [D]. Zhengzhou: Henan Agricultural University, 2018.
[29] 景鑫鑫. 几种钝化剂修复铅镉污染石灰性土壤的效果研
究[D]. 郑州: 河南农业大学, 2015.
JING X X. Effects of different stabilizers on the remediation for
heavy metal contaminated alkaline soil [D]. Zhengzhou: Henan
Agricultural University, 2015.
[30] 付煜恒, 张惠灵, 王宇, 等. 磷酸盐对铅镉复合污染土壤的
钝化修复研究[J]. 环境工程, 2017, 35(9): 176-180.
FU Y H, ZHANG H L, WANG Y, et al .. Immobilization of soil
contaminated by lead and cadmium using phosphate [J].
Environ. Eng., 2017, 35(9): 176-180.
[31] 孙新, 李琪, 姚海凤, 等. 土壤动物与土壤健康[J]. 土壤学报,
2021, 58(5): 1073-1083.
SUN X, LI Q, YAO H F, et al .. Soil fauna and soil health [J].
Acta Pedol. Sin., 2021, 58(5): 1073-1083.
[32] 张晓珂, 梁文举, 李琪. 我国土壤线虫生态学研究进展和展
望[J]. 生物多样性, 2018, 26(10): 1060-1073.
ZHANG X K, LIANG W J, LI Q. Recent progress and future
directions of soil nematode ecology in China [J]. Biodiversity
Sci., 2018, 26 (10): 1060-1073.
[33] NEHER D A. Role of nematodes in soil health and their use as
indicators [J]. J. Nematol., 2001, 33(4): 161-168.
[34] HAN D C, ZHANG X K, TOMAR V V S, et al.. Effects of heavy
metal pollution of highway origin on soil nematode guilds in North
Shenyang, China [J]. J. Environ. Sci., 2009, 21(2): 193-198.
[35] LI H X, HU F. Effect of bacterial-feeding nematode inoculation
on wheat growth and N and P uptake [J]. Pedosphere, 2001,
11(1): 57-62.
[36] XIAO H F, GRIFFITHS B, CHEN X Y, et al .. Influence of
bacterial-feeding nematodes on nitrification and the ammoniaoxidizing
bacteria (AOB) community composition [J]. Appl.
Soil Ecol., 2010, 45(3): 131-137.
[37] 王赢利, 王宏洪, 廖金铃, 等. 电子垃圾拆解地重金属污染
对稻田土壤线虫群落结构的影响[J]. 农业环境科学学报,
2015, 34(5): 874-881.
WANG Y L, WANG H H, LIAO J L, et al.. Effects of heavy metal
contamination on nematode communities in paddy soils of an ewaste
recycling area [J]. J. Agro-Environ. Sci., 2015, 34(5):
874-881.
[38] 成碧君, 刘良坡, 张红梅, 等. 重金属对煤矿区土壤线虫群
落结构的影响[J]. 山西农业科学, 2022, 50(12): 1680-1688.
CHENG B J, LIU L P, ZHANG H M, et al .. Effects of heavy
metals on soil nematode community structure in coal mine
areas [J]. Shanxi Agric. Sci., 2022, 50(12): 1680-1688.
[39] 吕莹. 线虫群落对大连滨海石油与土壤重金属污染及绿地生
态系统恢复的指示作用研究[D]. 大连: 辽宁师范大学, 2012.
LYU Y. The study of responses of nematode communites to
crude oil contamination, soil heavy metal pollution and
recovery of urban greenland in Dalian [D]. Dalian: Liaoning
Normal University, 2012.
[40] NAGY P, BAKONYI G, BONGERS T, et al .. Effects of
microelements on soil nematode assemblages seven years after
contaminating an agricultural field [J]. Sci. Total Environ.,
2003, 320(2): 131-143.
[41] ZHANG W D, XIAO Y, WANG X F, et al .. Soil nematode
community characteristics around the Gangue hill of Fushun
West Open-pit mine [J]. Helminthologia, 2011, 48(2): 116-123.
[42] DOROSZUK A. Populations under stress: analysis on the
interface between ecology and evolutionary genetics in
nematodes [D]. Wageningen: Wageningen University, 2007.
[43] 李孟洁. 小麦−玉米轮作体系中长期施肥对土壤线虫群落
结构的影响[D]. 郑州: 郑州大学, 2018.
LI M J. Effects of long-term fertilization on soil nematode
community structure in wheat-maize rotation system [D].
Zhengzhou: Zhengzhou University, 2018.
[44] 杨盼盼,黄菁华,张欣玥,等.长期施肥对渭北旱塬麦田土壤
线虫群落特征的影响[J]. 干旱地区农业研究, 2022, 40(5):
242-251.
YANG P P, HUANG J H, ZHANG X Y, et al .. Effects of longterm
fertilization on soil nematode community of wheat field in
Weibei dryland, China [J]. Agric. Res. Arid Areas, 2022, 40
(5):242-251.
[45] 金娜, 刘倩, 简恒. 植物寄生线虫生物防治研究新进展[J].
中国生物防治学报, 2015, 31(5): 789-800.
JIN N, LIU Q, JIAN H. Advances on biological control of plantparasitic
nematodes [J]. Chin. J. Biol. Control, 2015, 31(5):
789-800.
[46] 段玉玺. 植物线虫学[M]. 北京: 科学出版社, 2011: 52-53.
[47] JORÉ A R M, CARMEN G, MIGUEL E, et al.. Effect of mine
tailing on the spatial variability of soil nematodes from lead
pollution in La Union (Spain) [J]. Sci. Total Environ., 2014,
473-474:518-529.
猜你喜欢
今日农业(2022年14期)2022-09-15 01:44:26
能源工程(2021年5期)2021-11-20 05:50:44
宝钢技术(2021年4期)2021-09-10 07:23:24
生物质化学工程(2021年1期)2021-01-26 09:22:30
中国造纸(2020年9期)2020-10-20 05:33:36
环境影响评价(2020年3期)2020-09-18 07:15:52
中成药(2018年2期)2018-05-09 07:20:04
现代园艺(2017年19期)2018-01-19 02:49:46
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:02
天然产物研究与开发(2016年6期)2016-06-05 10:29:30
