
中国南方典型稻作梯田土壤有机碳积累及其来源表征
2025-02-16 00:00:00韩玥赵正田静龙国刚龚婧雯王廷好沈易毛子仪李恋卿张旭辉潘根兴
农业资源与环境学报 2025年1期



摘要:研究长期耕作管理下土壤有机碳(SOC)固持能力及其积累机制是当前农业可持续发展基础研究的前沿。我国南方山区稻作梯田是国际社会公认的全球重要农业遗产,本研究旨在剖析其千年可持续水稻生产下有机碳的积累特征,探索基于自然的农田固碳技术途径。以我国南方山区5处典型稻作梯田系统为研究对象,采集0~20 cm(表土)、20~40 cm(心土)和40~60 cm(底土)深度土壤样品,分析土壤有机碳形态与分布特征,同时采用生物标志物分子提取和GC-MS鉴定技术,分析木质素及氨基糖含量变化来解析有机碳的不同来源。结果表明:供试5处梯田表土有机碳积累水平差异极大,发育于沉积岩的四川新山梯田和高坎梯田黏粒含量高达40%,各层有机碳含量在15.58~47.78 g·kg-1范围内,而发育于花岗岩的江西上堡梯田、浙江云和梯田和湖南紫鹊界梯田的砂粒含量高达40%~66%,各层土壤有机碳含量在7.41~30.86 g·kg-1范围内。在深度上,土壤有机碳分布存在分层性:表土有机碳含量(17.98~47.78 g·kg-1)大幅高于心土、底土;相较于表土,心土层SOC含量降低24.8%~62.2%,底土层SOC含量则降低47.1%~76.0%。表土具有较高的木质素含量及较低的氧化程度,较高的MBC(微生物生物量碳)/SOC值、POC(颗粒态有机碳)/MAOC(矿物结合态有机碳)值及较低的DOC(溶解性有机碳)/SOC值;相反,心土、底土有机碳含量较为匮缺(除高坎梯田外,均低于20 g·kg-1),MAOC占比较高。所有样品的总有机碳含量与木质素酚含量间呈极显著的正相关关系(R2=0.909,Plt;0.01)。研究表明,植物源有机碳因颗粒态物理保护作用而在黏质水稻土中积累,这是千年稻作梯田有机碳稳定积聚的主要机制。
关键词:稻作梯田;土壤有机碳;木质素;氨基糖;有机碳固定;来源解析
中图分类号:S153.6 文献标志码:A 文章编号:2095-6819(2025)01-0139-10 doi: 10.13254/j.jare.2023.0827
土壤有机碳库(Soil Organic Carbon,SOC)是陆地系统中最大的碳库,是全球碳循环的重要组成部分,全球1 m深土壤中储存有机碳达15 000亿t[1]。因此,认识土壤有机碳库的本质及其在不同土壤中的稳定机制是土壤学研究的前沿领域[2]。植物-微生物相互作用控制着土壤有机碳输入、积累与持留,植物和微生物来源有机碳组分的分布、转化、保存和稳定影响着土壤有机碳库大小及稳定性[3-4]。因此,阐明植物源和微生物源碳的积累和分布可以更好地理解土壤有机碳变化。
自然生态系统中,土壤有机碳主要来源于植物残体和微生物代谢产物在土壤中的持留[5-6]。植物源有机碳进入土壤后,不同程度地被微生物分解、转化或利用,其未分解组分残留于土壤中[7]。因此,土壤有机碳的积累依赖于植物来源有机碳的残留。特别是以往认为木质素等芳香族化合物是难降解的有机碳组分,是土壤有机碳积累的主要贡献者。然而,特定条件下木质素也可以被分解,不能达到长期稳定[8]。相反,微生物在分解、转化和利用植物源有机物的同时,其代谢产物或死亡的残体也残留于土壤,通常能与矿物质结合达到稳定化[9]。因而,这部分有机碳往往比植物源有机碳更稳定,对稳定有机碳库的长期贡献可能更大。尽管如此,目前对不同生态系统中土壤有机碳水平与植物源和微生物源有机碳的持留关系还未有全面的了解。
我国具有悠久的水稻种植历史,稻田土壤具有较大的固碳潜力,对全球碳循环具有重要意义[10]。在人口压力较大的丘陵山区,沿等高线建造的梯田更利于粮食生产,同时还能减少径流和泥沙流失,从而保持水土[11]。通过千年来的持续生产和发展,梯田被证明是一种基于自然的山区可持续农业解决方案。Cao等[12]的研究表明,在我国耕地总面积中,梯田(梯地)面积占有不可低估的份额。梯田的建设、利用和管理会影响区域内碳动态、粮食安全和农业生态系统服务功能[13-14]。梯田化对有机碳储量的正向和负向影响均有报道[15-16],但其对有机碳的储存特征、变化的影响机制尚不清楚。我国南方地区的丘陵山地,存在着不同时期开垦的古老稻作梯田,至今仍持续发挥着水稻生产和乡村振兴的巨大支撑作用[17]。在千年尺度的水稻种植和耕作管理下,梯田土壤发育为特殊的人为土——水稻土,其有机碳固存特征是什么?固碳能力受何种机制控制?不同来源的有机碳相对贡献如何?了解这些问题对于认识梯田可持续生产的本质和梯田农业对控制气候变化的作用十分必要。
1 材料与方法
1.1 研究区概况
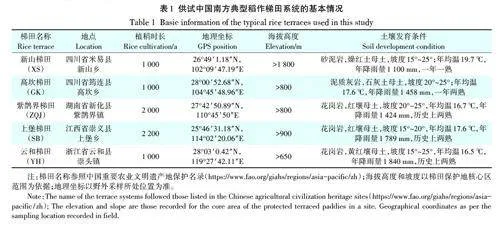
根据资料及数据统计分析[18],南方古老梯田集中分布于101°48′~119°29′E、22°33′~29°46′N 之间的亚热带山区和丘陵地区,地跨云南、四川、贵州、广西、湖南、江西、福建和浙江等8个省份,涵盖红壤、黄壤、紫色土、石灰土和冲积土等土壤发生类型,养育着汉、壮、苗、哈尼、傈僳和畲等众多民族聚落,最古老的梯田迄今已有2 500多年历史。本研究选取了我国南方5个典型稻作梯田,分别为:四川米易新山傈僳族梯田(XS)、四川筠连县高坎僰人梯田(GK)、湖南新化县紫鹊界梯田(ZQJ)、江西崇义县上堡梯田(SB)和浙江云和县的云和梯田(YH)。采样点的基本信息见表1。
1.2 样品采集与处理
土壤采样在水稻收获后进行,在2021年10月—2022年8月期间完成。通过查阅资料及走访当地农民,调查了解梯田区植稻历史,确定采样研究的核心区域和代表性采样田块。在核心区域内,选取坡度坡向基本一致、梯田形态相对完好的稻作梯田进行土壤样品采集。一个梯田系统选取3个代表性样田,每个采样田相距至少100 m。采样时,去除地表凋落物,用不锈钢钻土器分别采集0~20 cm(表土层)、20~40cm(心土层)和40~60 cm(底土层)等分层土壤样品。每样田梅花形随机采集3点混匀构成1个混合样本。将样品装入密封袋后置于内有冰袋的保鲜箱中,取样后48 h内送至实验室。
抵达实验室后,拣出植物根系及砾石,混匀后四分法将样品分成2 份,一份直接筛过2 mm 孔筛,-20 ℃保存用于测定土壤微生物指标;另一份自然风干,按鲁如坤[19]的土壤农化分析法,分别磨碎过2、1mm和0.15 mm孔筛,存于自封袋中,用作土壤理化分析和有机质提取-生物标志物测定。
1.3 分析测定方法
土壤基本理化性质测定参照鲁如坤[19]的方法。土壤矿质颗粒粒径采用比重计法;土壤pH(H2O)以土水比1∶2.5用pH计(Metter-Toledo pH meter)测定;土壤有机碳(SOC)采用重铬酸钾氧化-外加热法测定;土壤总氮(TN)采用半微量凯氏定氮法测定;土壤有效磷(AP)采用NH4F-HCl浸提-钼锑抗比色法测定;土壤速效钾(AK)采用NH4NO3 浸提-火焰光度法测定。土壤微生物生物量碳(MBC)采用氯仿熏蒸-K2SO4 提取-TOC 仪(multi N/C 3100 TOC,Analytik Je⁃na,德国)测定。
土壤颗粒态有机碳(POC)和矿物结合态有机碳(MAOC)的分离:称取10 g 过2 mm 筛的风干土样于50 mL离心管中,加入20 mL 5 g·L-1六偏磷酸钠(土液比1∶2),以90 r·min-1的频率摇床振荡18 h,振荡后悬液倾倒在53 μm 孔筛上,反复用超纯水冲洗筛上物质,直到过筛水清澈为止。筛上物质固持的碳即为POC,收集至50 mL 离心管中;过筛的悬浊液静置沉降,倒去上清液,转移至离心管中,沉淀出的物质所固持的碳即为MAOC。这些分离物均在冷干机中冷冻干燥后,测定其有机碳浓度。
木质素酚类分析依据Otto 等[20]提出和Chen等[21]介绍的氧化铜(CuO)氧化和GC-MS定量鉴定法。该方法在GC/MS 分析得到的图谱中,依据样本中出现的木质素单体酚峰值与内标物峰值的相对数值换算出样本中木质素单体酚的浓度(mg·kg-1),再以样本有机碳含量(SOC)为基准换算丰度值(mg·g-1 SOC),最后,采用下列公式估计植物源碳的比例[22]:
参照文献[25]的方法提取和测定土壤氨基葡萄糖(GluN)、氨基半乳糖(GlaN)、胞壁酸(MurN)和甘露糖胺(ManN)的浓度。有机碳中微生物残体碳(MRC)丰度按下式估算[26]:
1.4 数据处理
所有数据均以3个重复样田的平均值±标准差表示。数据处理和分析采用Excel 2019和SPSS 20.0软件。绘图采用Origin 2021软件。采用单因素方差分析(One way ANOVA)和Duncan检验来检验不同深度间所有土壤指标的差异,显著性概率设为Plt;0.05。
2 结果与分析
2.1 土壤基本理化性质
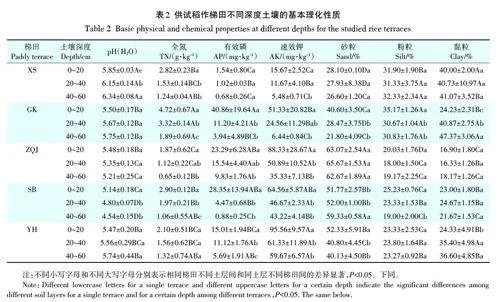
供试5个稻作梯田系统不同深度土壤基本理化性质见表2。所测土壤均为酸性,pH 在4.54~6.34之间。这些梯田系统间土壤矿质颗粒组成存在显著性差异,XS和GK梯田表土为壤质黏土,粉、黏粒含量显著高于其他三地;而YH梯田为黏壤土,SB梯田和ZQJ梯田则为砂质黏壤土;所有样田不同土层间,仅在GK梯田中观察到黏粒淋失淀积现象。供试梯田表土全氮含量介于1.87~4.72 g·kg-1,以GK梯田为最高;表土有效磷含量介于1.54~40.86 mg·kg-1,也以GK梯田为最高;然而,表土速效钾含量介于15.67~95.56 mg·kg-1,以YH梯田为最高。所有梯田系统的土壤全氮、有效磷和有效钾含量均随土层深度增加而降低。
2.2 土壤有机碳含量及形态分布
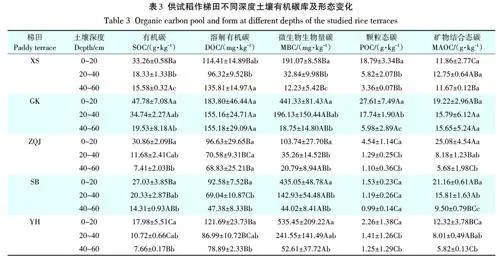
供试稻作梯田系统不同深度土壤有机碳含量及形态分布如表3所示。表土SOC含量在17.98~47.78g·kg-1,POC 含量在1.53~27.61 g·kg-1,DOC 含量在92.58~183.80 mg · kg-1,MBC 含量在103.74~535.45mg·kg-1。SOC和MBC含量随土壤深度增加而降低。在同一梯田,相对于表土,心土层土壤SOC含量降低24.8%~62.2%,MBC 含量降低54.9%~82.8%;底土层SOC含量降低47.1%~76.0%,MBC降低80.0%~95.8%。DOC含量在YH梯田和SB梯田随土层深度降低而降低,而在其他梯田系统,不同深度间无显著性差异。POC与MAOC的含量均随土壤深度增加而降低。在XS梯田和GK梯田,表土POC含量高于MAOC含量。然而,随土壤深度增加,POC含量急剧降低,MAOC含量逐渐大于POC含量;而在YH、SB和ZQJ梯田中,不同土壤深度POC含量均小于MAOC含量。
2.3 土壤木质素酚类和氨基糖的丰度
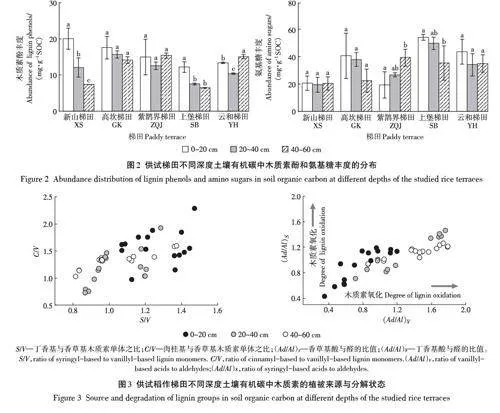
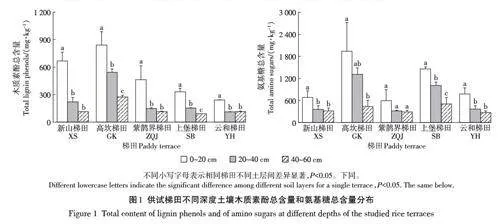
供试稻作梯田不同深度土壤的木质素酚类和氨基糖含量以及计算得到的在有机碳中的丰度变化如图1和图2所示(木质素酚类和氨基糖的各单体浓度见附表1和附表2(扫描首页OSID码查看)。土壤木质素酚含量随土层深度增加呈降低趋势,表土层显著高于心土层和底土层;就丰度而言,在XS和SB梯田,木质素酚类丰度在表土层高于心土层与底土层;且不同梯田系统间并没有明显的差异分布规律。不同的是,土壤氨基糖含量随土层深度增加呈下降趋势,表土层显著高于心土层和底土层。氨基糖的丰度在不同土层深度间无显著差异,但不同梯田间差异较大。例如,底土层氨基糖丰度在有机碳含量较低的ZQJ、SB和YH等梯田系统中高于XS和GK梯田。
供试稻作梯田不同深度土壤木质素的植被来源与氧化状态如图3所示。不同深度土壤中丁香基/香草基(S/V)和肉桂基香草基(C/V)值相对集中在同一区域,指示各层间木质素植被来源无明显差异。表土香草基酸醛比([ Ad/Al)V]和丁香基酸醛比([ Ad/Al)S]值显著低于心土层和底土层(Plt;0.05),提示表土木质素酚的氧化分解程度相对较低,即相对于心土和底土较为新鲜。
采用公式(1)~公式(3)分别估算得到植物源和微生物源有机碳的相对贡献,其在供试稻作梯田不同深度土壤间的分布信息见附图1(扫描首页OSID 码查看)。在表土层和心土层,有机碳含量较高的XS、GK和ZQJ等梯田中植物源有机碳相对贡献高于微生物源有机碳贡献,在YH和SB等表层有机碳含量较低的梯田中则是微生物源有机碳相对贡献略高于植物源贡献。总体而言,随着土壤深度增加,土壤有机碳含量降低,植物源有机碳的贡献也随之降低(Plt;0.01),而微生物源有机碳的相对贡献无明显变化。
3 讨论
3.1 稻作梯田土壤有机碳积累与碳库分布
水稻土是长期人为水耕熟化的农耕实践下培育的特殊人为土壤[27]。尽管耕作数百年甚至上千年,耕层土壤有机碳含量可能持续增长[28]。本研究中耕层土壤有机碳含量变化与梯田稻作历史没有明显的对应关系。在供试的5个典型梯田系统中,表土有机碳含量介于17.98(云和梯田)~47.78 g·kg-1(高坎梯田),尽管变化范围极大,但在土层深度上均存在极显著的表聚性,稻田的耕作管理主要发生于耕层,由于水稻土犁底层的存在,导致输入的有机碳高度聚集于表层[29]。根据表3中土壤有机碳的分布深度,有机碳的表聚强度(有机碳含量的深度递降斜率)与表土有机碳水平呈极显著正相关关系,这表明长期的耕作培育使土壤有机碳越来越向耕层聚集。
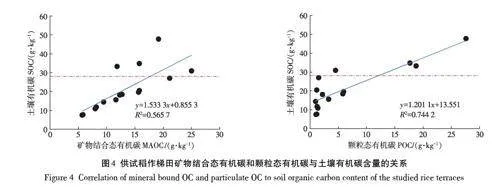
在旱地土壤中,有机碳的固持与黏粒的物理保护能力有关。本研究中(表3),无论是不同土层深度间,还是不同梯田间,有机碳含量都与相应的黏粒含量没有显著的统计相关关系。按照不同土层深度统计,估算的有机碳固存量[有机碳质量(g)/黏粒质量(g)]介于(0.12±0.04)(表土)、(0.06±0.02)(心土层)和(0.04±0.02)(底土层)。这说明稻作梯田耕层和心土层有机碳固存不适用黏粒保护理论(0.02~0.03)[30]。本研究中,MAOC 也与黏粒含量没有显著的相关关系。上述结果均说明,是团聚体的物理保护能力而不是黏粒的结合保护能力影响着稻田中有机碳的固持[31-32]。本研究中所有土壤样品有机碳含量与MAOC和POC都存在显著的正相关关系(图4)。值得注意的是,当不同梯田土壤样品中有机碳含量在28 g·kg-1(图4中虚横线)以上时,控制有机碳水平的形态不是MAOC含量,而是POC含量。最新研究显示,颗粒态有机碳的积累可以较好解释土地利用[33]和生态系统管理下土壤固碳[21,34],且可以进一步联系到土壤的生态系统服务功能[35]。
上述结果提示,特定土地利用下矿物质-有机质-微生物间的复杂相互作用控制着稻田土壤有机碳积累[36]。土壤DOC的含量代表易利用的有机碳含量,而MBC 的含量代表有机碳的微生物利用[37]。本研究中不同梯田DOC/SOC 值和DOC/MBC 值均表现为表土显著低于心土、底土,而MBC/SOC值表现为表土高于心土,底土(表3),说明耕层有机碳处于相对积累状态,而心土、底土有机碳处于相对分解状态,耕层土壤的有机碳微生物可利用性高于心底土。不同深度土壤MAOC、POC、MBC含量和DOC含量有一定的显著差异(表3),说明心土、底土有机碳含量匮缺,因为稻田土壤中存在犁底层,阻隔了有机碳的输入。耕层有机碳,特别是作物秸秆残茬在分解后,小部分可溶性有机物随水在田面横向迁移和剖面纵向渗滤[38]。这部分溶解的可移动有机质与颗粒态有机质和矿物结合态有机质的相对关系可以代表剖面中有机质的积累和稳定[39-40]。表土的DOC/POC值低于心土层,而POC/MAOC 值则相反(表3)。不同梯田间,砂粒含量gt;30%、黏粒含量lt;40% 的SB、ZQJ 和YH 梯田,其表土的DOC/POC 值高于XS 和GK 梯田,而POC/MAOC值则相反。供试梯田母土土壤发生类型涵盖红壤、黄壤和石灰土[41],其黏粒矿物种类存在的差异可能掩盖了矿物结合态有机碳含量与黏粒含量的关系,但在砂粒含量较高而黏粒含量较低时,有利于有机质的微生物矿化而不利于有机质的结合稳定。与发育于细质地河海沉积物的平原水稻土[28]不同,ZQJ、SB和YH等梯田发育于花岗岩风化物,尽管稻作已持续千年以上,但仍不能有效积累有机碳。
3.2 梯田土壤有机碳积累的植物和微生物来源贡献
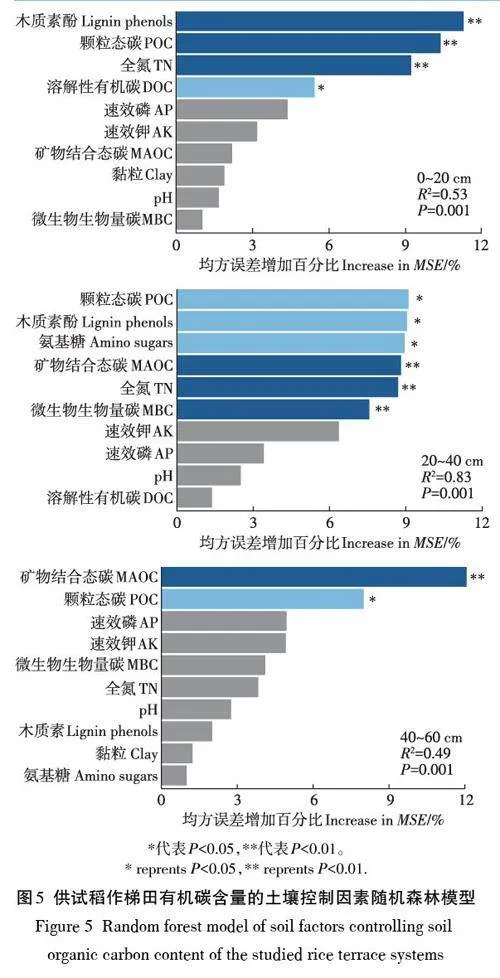
土壤有机碳库的固存取决于植物源有机碳的直接输入与微生物对植物残体的分解利用的平衡[42]。已有研究表明,POC是植物残体来源的新鲜有机碳的载体[43],而MAOC来源于植物残体微生物分解残余物及微生物代谢产物[9]。在稻田土壤中,POC主要来源于水稻根系或其他植物残体输入[44]。供试梯田系统表土木质素含量与其POC含量(R2=0.835,Plt;0.01)和SOC含量(R2=0.814,Plt;0.01)均呈显著正相关。这种关系在心土层和底土层减弱。随机森林模型(图5)揭示,控制总有机碳积累水平的极显著(Plt;0.01)因素分别是:木质素酚含量、颗粒态碳和全氮(表土层),矿物结合态碳、全氮和微生物生物量碳(心土层)和矿物结合态碳(底土层)。
木质素酚含量在表土层显著高于心土层,但不同土层深度的土壤氨基糖丰度不存在显著差异,且底土层氨基糖的丰度在有机碳含量低的ZQJ、SB和YH梯田中高于XS和GK梯田。这表明,供试梯田土壤中有机碳的水平主要受控于植物源有机碳的固存,有机碳匮缺的土壤中微生物来源上升为主要贡献。木质素酚单体中,丁香基/香草基(S/V)的值和肉桂基/香草基(C /V)的值常用来指示土壤有机质来源的植被类型[45],而香草基的酸醛比([ Ad/Al)V]和丁香基的酸醛比([ Ad/Al)S]用来评估土壤木质素酚的降解状态[23]。耕层土壤(Ad/Al)V和(Ad/Al)S值较低,代表木质素单体的氧化分解程度较低,也就是说植物来源的碳在表土中输入效率大于其分解水平,这促进了耕层有机碳持续积累[32]。不同土壤间C/V 和S/V 值无显著差异,说明植物源有机质的来源结构稳定,符合长期水耕稻作的持续影响。且供试梯田不同深度样品的木质素酚含量与有机碳含量间存在极显著的线性相关性(图6),这进一步提示,木质素为代表的来自水稻根系残体有机碳为主要来源,而不是微生物残余物有机碳,可以诠释黏质梯田水稻土(XS和GK梯田)中的有机碳固存。这与Gherardi等[46]和Dai等[47]关于植物源碳驱动了森林土壤和稻作农田中SOC 积累的报道相一致。因此,木质素代表的植物源有机碳的物理保护是梯田水稻土有机碳长期稳定积累的原因。
4 结论
(1)供试的5个南方山区典型高山梯田系统,虽然稻作农业持续了千年以上,但土壤有机碳含量丰缺不等,发育于花岗岩的梯田(柴鹊界、上堡、云和梯田)因质地较粗,有机碳积累大幅度低于发育于灰岩和泥质岩的梯田(新山、高坎梯田)。
(2)不同梯田的有机碳剖面分布呈明显的分层性:表土有机碳含量较高,且木质素为代表的植物源有机碳及颗粒态碳有相当的份额,因而微生物可利用性较高,心土、底土有机碳处于匮缺状态,以矿物结合态碳为主。
(3)从有机碳来源和形态分布上来看,不同梯田存在相似性,表明木质素丰度可以指示稻田有机碳的积累水平及其来源。有机碳分布与土壤的生态系统服务功能及农田生产力的关系还需要进一步研究。
参考文献:
[1] LAL R, MONGER C, NAVE L, et al. The role of soil in regulation of
climate[J]. Philosophical Transactions of the Royal Society of London
Series B, Biological Sciences, 2021, 376(1834):20210084.
[2] 潘根兴, 丁元君, 陈硕桐, 等. 从土壤腐殖质分组到分子有机质组学
认识土壤有机质本质[J]. 地球科学进展, 2019, 34(5):451-470.
PAN G X, DING Y J, CHEN S T, et al. Exploring the nature of soil
organic matter from humic substances isolation to SOMics of molecular
assemblage[J]. Advances in Earth Science, 2019, 34(5):451-470.
[3] SOKOL N W, SANDERMAN J, BRADFORD M A. Pathways of
mineral-associated soil organic matter formation:integrating the role of
plant carbon source, chemistry, and point of entry[J]. Global Change
Biology, 2019, 25(1):12-24.
[4] COTRUFO M F, RANALLI M G, HADDIX M L, et al. Soil carbon
storage informed by particulate and mineral-associated organic matter
[J]. Nature Geoscience, 2019, 12:989-994.
[5] ANGST G, MUELLER K E, NIEROP K G J, et al. Plant- or microbialderived?
A review on the molecular composition of stabilized soil
organic matter[J]. Soil Biology and Biochemistry, 2021, 156:108189.
[6] KÖGEL-KNABNER I. The macromolecular organic composition of
plant and microbial residues as inputs to soil organic matter:fourteen
years on[J]. Soil Biology and Biochemistry, 2017, 105:A3-A8.
[7] COTRUFO M F, WALLENSTEIN M D, BOOT C M, et al. The
Microbial Efficiency-Matrix Stabilization (MEMS) framework
integrates plant litter decomposition with soil organic matter
stabilization:do labile plant inputs form stable soil organic matter?[J].
Global Change Biology, 2013, 19(4):988-995.
[8] JIA J, LIU Z G, HAGHIPOUR N, et al. Molecular 14C evidence for
contrasting turnover and temperature sensitivity of soil organic matter
components[J]. Ecology Letters, 2023, 26(5):778-788.
[9] LIANG C, SCHIMEL J P, JASTROW J D. The importance of anabolism
in microbial control over soil carbon storage[J]. Nature Microbiology,
2017, 2:17105.
[10] PAN G X, LI L Q, WU L S, et al. Storage and sequestration potential
of topsoil organic carbon in China’s paddy soils[J]. Global Change
Biology, 2004, 10(1):79-92.
[11] CAO Y, WU Y H, ZHANG Y J, et al. Landscape pattern and
sustainability of a 1300-year-old agricultural landscape in
subtropical mountain areas, southwestern China[J]. International
Journal of Sustainable Development amp; World Ecology, 2013, 20(4):
349-357.
[12] CAO B W, YU L, NAIPAL V, et al. A 30 m terrace mapping in China
using Landsat 8 imagery and digital elevation model based on the
Google Earth Engine[J]. Earth System Science Data, 2021, 13(5):
2437-2456.
[13] LIU X H, HE B L, LI Z X, et al. Influence of land terracing on
agricultural and ecological environment in the Loess Plateau regions
of China[J]. Environmental Earth Sciences, 2011, 62(4):797-807.
[14] ZHANG J H, WANG Y, LI F C. Soil organic carbon and nitrogen
losses due to soil erosion and cropping in a sloping terrace landscape
[J]. Soil Research, 2015, 53(1):87.
[15] CHEN D, WEI W, DARYANTO S, et al. Does terracing enhance soil
organic carbon sequestration? A national-scale data analysis in China
[J]. Science of the Total Environment, 2020, 721:137751.
[16] DENG C X, ZHANG G Y, LIU Y J, et al. Advantages and
disadvantages of terracing:a comprehensive review[J]. International
Soil and Water Conservation Research, 2021, 9(3):344-359.
[17] 闵庆文, 骆世明, 曹幸穗, 等. 农业文化遗产:连接过去与未来的桥
梁[J]. 农业资源与环境学报, 2022, 39(5):855-868. MIN Q W,
LUO S M, CAO X S, et al. Agri-cultural heritage:a bridge between
past and the future[J]. Journal of Agricultural Resources and
Environment, 2022, 39(5):855-868.
[18] 李惠, 宋维峰, 马建刚. 中国南方古梯田的分布与景观结构特征
[J]. 中国水土保持科学, 2023, 21(2):93-106. LI H, SONG W F,
MA J G. Distribution and landscape structures of ancient terraces in
southern China[J]. Science of Soil and Water Conservation, 2023, 21
(2):93-106.
[19] 鲁如坤. 土壤农业化学分析方法[M]. 北京:中国农业科学技术出
版社, 2000. LU R K. Analytical methods for soil and agrochemistry
analysis[M]. Beijing:China Agricultural Science and
Technology Press, 2000.
[20] OTTO A, SHUNTHIRASINGHAM C, SIMPSON M J. A comparison
of plant and microbial biomarkers in grassland soils from the prairie
ecozone of Canada[J]. Organic Geochemistry, 2005, 36(3):425-448.
[21] CHEN S T, FENG X, LIN Q M, et al. Pool complexity and molecular
diversity shaped topsoil organic matter accumulation following
decadal forest restoration in a Karst terrain[J]. Soil Biology and
Biochemistry, 2022, 166:108553.
[22] CHEN X B, HU Y J, XIA Y H, et al. Contrasting pathways of carbon
sequestration in paddy and upland soils[J]. Global Change Biology,
2021, 27(11):2478-2490.
[23] HAUTALA K, PEURAVUORI J, PIHLAJA K. Estimation of origin of
lignin in humic DOM by CuO oxidation[J]. Chemosphere, 1997, 35
(4):809-817.
[24] BURGESS M S, MEHUYS G R, MADRAMOOTOO C A.
Decomposition of grain-corn residues(Zea mays L.):a litterbag study
under three tillage systems[J]. Canadian Journal of Soil Science,
2002, 82(2):127-138.
[25] ZHANG X D, AMELUNG W. Gas chromatographic determination of
muramic acid, glucosamine, mannosamine, and galactosamine in soils
[J]. Soil Biology and Biochemistry, 1996, 28(9):1201-1206.
[26] JOERGENSEN R G. Amino sugars as specific indices for fungal and
bacterial residues in soil[J]. Biology and Fertility of Soils, 2018, 54
(5):559-568.
[27] IUSS Working Group WRB. World reference base for soil resources.
International soil classifcation system for naming soils and creating
legends for soil maps[M]. 4th Edition. Vienna:IUSS,2022.
[28] LIU Y L, WANG P, DING Y J, et al. Microbial activity promoted with
organic carbon accumulation in macroaggregates of paddy soils under
long-term rice cultivation[J]. Biogeosciences, 2016, 13(24):6565-
6586.
[29] 常琳溪, 梁新然, 王磊, 等. 中国稻田土壤有机碳汇特征与影响因
素的研究进展[J]. 土壤, 2023, 55(3):487-493. CHANG L X,
LIANG X R, WANG L, et al. Characteristics and influencing factors
of soil organic carbon sink in paddy fields in China:a review[J]. Soils,
2023, 55(3):487-493.
[30] HASSINK J. The capacity of soils to preserve organic C and N by
their association with clay and silt particles[J]. Plant and Soil, 1997,
191(1):77-87.
[31] 周萍, 宋国菡, 潘根兴, 等. 三种南方典型水稻土长期试验下有机
碳积累机制研究Ⅱ. 团聚体内有机碳的化学结合机制.[J]. 土壤学
报, 2009, 46(2):263-273. ZHOU P, SONG G H, PAN G X, et al.
SOC enhancement in three major types of paddy soils in a long-term
agro-ecosystem experiment in south China Ⅱ. Chemical binding and
protection in micro-aggregate size fractions[J]. Acta Pedologica
Sinica, 2009, 46(2):263-273.
[32] 刘纯, 赵正, 刘晓雨, 等. 乡村小流域不同土壤景观表土有机质团
聚体分布与分子组成变化[J]. 植物营养与肥料学报, 2022, 28(5):
798-811. LIU C, ZHAO Z, LIU X Y, et al. Changes in aggregate
distribution and molecular composition of organic matter of topsoil
across soil landscapes within a small watershed in a rural area[J].
Journal of Plant Nutrition and Fertilizers, 2022, 28(5):798-811.
[33] 赵正, 刘纯, 尚明月, 等. 宁镇丘陵区村域小流域不同土壤景观下
表土质量变化及评价[J]. 水土保持学报, 2022, 36(4):265-276.
ZHAO Z, LIU C, SHANG M Y, et al. Changes and evaluation of
topsoil quality across different soil - landscapes in a village level
watershed of Nanjing-Zhenjiang hilly region[J]. Journal of Soil and
Water Conservation, 2022, 36(4):265-276.
[34] 陈硕桐, 夏鑫, 丁元君, 等. 不同形态秸秆还田下乌栅土耕层土壤
有机质含量与组成变化[J]. 中国农业科学, 2023, 56(13):2518-
2529. CHEN S T, XIA X, DING Y J, et al. Changes in topsoil
organic matter content and composition of a gleyic stagnic anthrosol
amended with maize residue in different forms from the Tai Lake
Plain, China[J]. Scientia Agricultura Sinica, 2023, 56(13):2518-
2529.
[35] FENG X, XIA X, CHEN S T, et al. Amendment of crop residue in
different forms shifted micro-pore system structure and potential
functionality of macroaggregates while changed their mass proportion
and carbon storage of paddy topsoil[J]. Geoderma, 2022, 409:115643.
[36] LI H C, VAN DEN BULCKE J, MENDOZA O, et al. Soil texture
controls added organic matter mineralization by regulating soil
moisture:evidence from a field experiment in a maritime climate[J].
Geoderma, 2022, 410:115690.
[37] SPARLING G P. Ratio of microbial biomass carbon to soil organic
carbon as a sensitive indicator of changes in soil organic matter[J].
Soil Research, 1992, 30(2):195.
[38] HE Y, LEHNDORFF E, AMELUNG W, et al. Drainage and leaching
losses of nitrogen and dissolved organic carbon after introducing
maize into a continuous paddy-rice crop rotation[J]. Agriculture,
Ecosystems amp; Environment, 2017, 249:91-100.
[39] ANGST G, MUELLER K E, CASTELLANO M J, et al. Unlocking
complex soil systems as carbon sinks:multi-pool management as the
key[J]. Nature Communications, 2023, 14(1):2967.
[40] YU W J, HUANG W J, WEINTRAUB-LEFF S R, et al. Where and
why do particulate organic matter(POM) and mineral-associated
organic matter(MAOM)differ among diverse soils?[J]. Soil Biology
and Biochemistry, 2022, 172:108756.
[41] 熊毅, 李庆逵. 中国土壤[M]. 第2 版. 北京:科学出版社, 1987.
XIONG Y, LI Q K. China soils[M]. 2nd Edition. Beijing:Science
Press, 1987.
[42] CHAOPRICHA N T, MARÍN-SPIOTTA E. Soil burial contributes to
deep soil organic carbon storage[J]. Soil Biology and Biochemistry,
2014, 69:251-264.
[43] WANG S Y, GAO Y, JIA J J, et al. Vertically stratified water source
characteristics and associated driving mechanisms of particulate
organic carbon in a large floodplain lake system[J]. Water Research,
2022, 209:117963.
[44] 祝贞科, 肖谋良, 魏亮, 等. 稻田土壤固碳关键过程的生物地球化
学机制及其碳中和对策[J]. 中国生态农业学报(中英文), 2022, 30
(4):592 -602. ZHU Z K, XIAO M L, WEI L, et al. Key
biogeochemical processes of carbon sequestration in paddy soil and
its countermeasures for carbon neutrality[J]. Chinese Journal of Eco-
Agriculture, 2022, 30(4):592−602.
[45] THEVENOT M, DIGNAC M F, RUMPEL C. Fate of lignins in soils:a
review[J]. Soil Biology and Biochemistry, 2010, 42(8):1200-1211.
[46] GHERARDI L A, SALA O E. Global patterns and climatic controls of
belowground net carbon fixation[J]. Proceedings of the National
Academy of Sciences of the United States of America, 2020, 117(33):
20038-20043.
[47] DAI G H, ZHU S S, CAI Y, et al. Plant-derived lipids play a crucial
role in forest soil carbon accumulation[J]. Soil Biology and
Biochemistry, 2022, 168:108645.
猜你喜欢
造纸信息(2022年8期)2022-11-10 08:19:20
上海包装(2019年8期)2019-11-11 12:16:14
科技资讯(2017年1期)2017-03-27 11:19:38
江苏农业科学(2016年8期)2017-02-15 18:40:19
江苏农业科学(2016年8期)2017-02-15 18:40:04
江苏农业科学(2016年10期)2017-02-05 01:33:41
天津造纸(2016年1期)2017-01-15 14:03:29
湖南大学学报·自然科学版(2015年12期)2016-03-05 12:00:47
中国造纸学报(2015年1期)2015-12-16 19:35:25
安徽农学通报(2015年19期)2015-10-23 01:11:53
