
散生木荷对4a生闽楠生长的影响
2025-02-11 00:00:00欧文胜
防护林科技 2025年1期





摘要:以南平市建阳区国有林场保留木荷散生木的造林山场为研究对象,分析散生木荷对4a生闽楠生长的影响。结果表明,散生木荷对以其为中心的2倍树冠面积圆形样地内闽楠的生长具有抑制作用,散生木荷对闽楠的树高、树冠长度和当年树高生长量具有极显著的抑制作用,对其地径和侧枝数具有显著的抑制作用,对冠幅、圆满度、树冠率、尖削度、枝下高、枝间距有抑制作用,但不显著;同时造成闽楠生长分化,长势不均匀。
关键词:散生木;木荷;闽楠;生长;变异系数
中图分类号:S725.7"""""""""""""""""" 文献标识:A"""""""" 文章编号:
Effects of scattered Schima superba on growth
of 4-year old Phoebe bournei
OU Wensheng
(Forestry bureau of Nanping City, Nanping354200, Fujian,China)
Abstract:The afforestation mountain with scattered Schima superba in Jianyang District of Nanping City taked as the research object, the effect of scattered Schima superba on the growth of 4-year old Phoebe bournei was analyzed.The results showed that the scattered Schima superba had an inhibitory effect on the growth of Phoebe bournei in the circular plot with twice the crown area as the center. The scattered Schima superba had a very significant inhibitory effect on the tree height, crown length and current-year tree height growth of Phoebe bournei, and had a significant inhibitory effect on its ground diameter and number of lateral branches. It had an inhibitory effect on crown width, fullness, crown rate, tapering degree, branch height and branch spacing, but it was not significant, which also causes growth differentiation and uneven growth in Phoebe bournei.
Key words: scattered trees; Schima superba; Phoebe bournei; growth; coefficient of variation
松材线虫病又称松枯萎病,是一种毁灭性虫害,在福建发生并流行成灾[1],为切实降低因松材线虫病造成松林大面积死亡的风险,构建健康、稳定、高效的森林生态系统,2020年福建省林业局开展松林改造提升行动,预测将采伐松林面积16.00万hm2[2],松林改造提升技术要点要求保留阔叶树[3],木荷作为乡土阔叶树[4]以散生木的形式得到广泛保留。保留木对退化森林生态系统结构与功能的维持和恢复具有多方面的生态效应,可有效减少采伐对生态系统结构和功能所造成的损失[5、6],同时也与周围林木存在竞争关系[7-9],目前国内关于对散生木或保留木对人工更新幼林生长影响方面的研究鲜有报道,本文通过分析采伐迹地上保留的木荷散生木对周边闽楠幼树生长的影响,可为保留散生木的采伐迹地造林更新提供理论依据。
1 研究区概况
试验地设在南平市建阳区国有林场,位于建阳区潭城街道严墩村054林班内,海拔150~325m。试验地位于武夷山南麓,属中亚热带季风性气候,光热资源丰富,年平均日照时数1802h,年平均气温18℃,无霜期282d,雨季集中,年平均降雨量1700~2400mm。土壤类型以山地红壤。
2022年建阳区国有林场选择连续留圃培育的3年生闽楠、木荷、枫香、赤皮青冈、银杏等容器苗,在建阳区潭城街道严墩村054林班皆伐改造提升且保留阔叶树的采伐迹地上造林,按照株行距2.0m×2.5m,初植密度130株/亩,挖穴规格50×40×30cm,施总养分45%的复合肥0.2kg/株,每年抚育2次。造林山场立地肥沃,土壤为山地红壤。
2 研究方法
2.1 样地设置与数据收集
2023年11月,根据造林山场地形、散生木荷和闽楠分布情况,选择立地条件基本一致的地段,设置9块标样地,其中:以散生木荷为中心的试验样地6块,无散生木遮挡的对照样地3块。试验样地为以木荷为中心,设置树冠面积2倍的圆形样地,对照样地为试验样地平均面积的圆形样地。样地基本情况见表1。
2.2.生长性状调查
对每块样地进行调查,共调查闽楠58株。调查内容主要包括样地内每株闽楠的地径、树高、冠幅、枝下高、侧枝数、当年树高生长量。根据树冠长度=树高-枝下高,枝间距=侧枝数量/树冠长度,树冠圆满度=冠幅/树冠长度,树冠率=树冠长度/树高,尖削度=地径/树高,计算枝间距、树冠圆满度、树冠率、尖削度。
2.3 数据处理
用Excel2007和DPS9.1软件对数据进行统计分析,采用单因素方差分析法分析,表中数据为平均值±标准差,变异系数CV=(标准差/平均值)×100%。
3" 结果与分析
3.1散生木荷对闽楠生长的影响
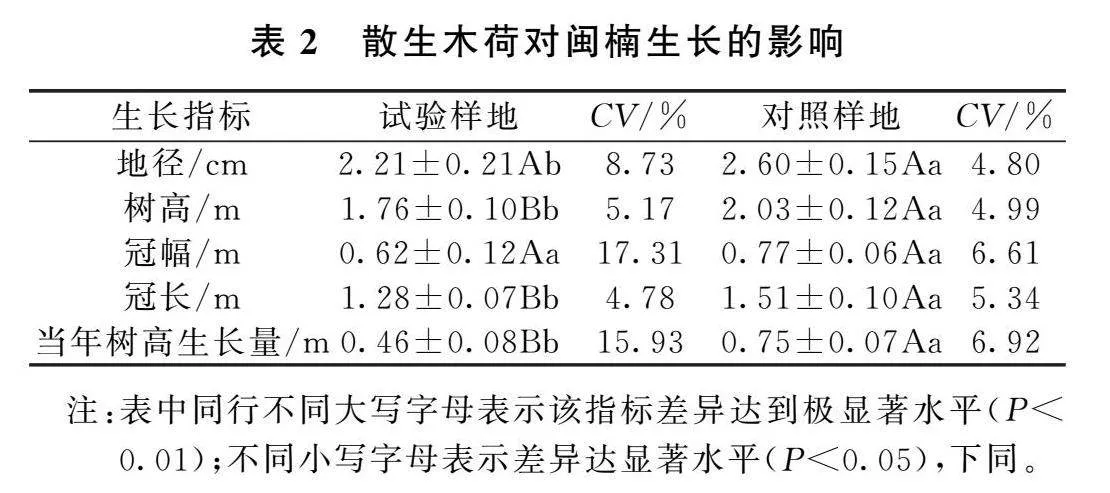
散生木荷对闽楠生长的影响见表2、表3,由表可知试验样地闽楠的平均地径显著低于对照样地闽楠平均地径,为对照样地的85.0%,试验样地闽楠的平均地径变异系数为8.37%,明显高于对照样地。试验样地闽楠的平均树高、树冠长度、当年树高生长量分别为对照样地的86.7%、84.8%、61.3%,差异均达到极显著水平,试验样地闽楠的平均树高和冠长的变异系数差异不大,试验样地闽楠的平均树高变异系数略大于对照样地,对照样地闽楠的平均冠长变异系数略大于试验样地,试验样地闽楠当年树高生长量变异系数为15.93%,明显大于对照样地,为对照样地的2.30倍。试验样地闽楠的平均冠幅比对照样地的少0.15m,两者差异不显著,试验样地闽楠的平均冠幅变异系数与对照样地存在明显差异,为对照样地的2.62倍。散生木荷对实验样地内的闽楠地径、树高、树冠长度和当年树高生长有的抑制作用显著,但对冠幅的生长影响不大,同时明显影响闽楠地径、冠幅、当年树高生长的整齐度,造成闽楠生长分化。
3.2散生木荷对闽楠干形的影响
散生木荷对闽楠干形的影响见表4、表5,试验样地内闽楠的平均圆满度比对照样地的低0.03,二者差异不显著。试验样地内闽楠的平均树冠率略小于照样地的,试验样地内闽楠的平均尖削度比照样地的低0.02,差异均不显著。试验样地闽楠干形指标的变异系数均大于对照样地,其中圆满度变异系数差异明显,试验样地闽楠圆满度变异系数为13.09,是对照样地的5.45倍。散生木荷对实验样地内的闽楠地径、树高、树冠长度和当年树高生长有的抑制作用,但不显著,同时明显影响闽楠的圆满度。
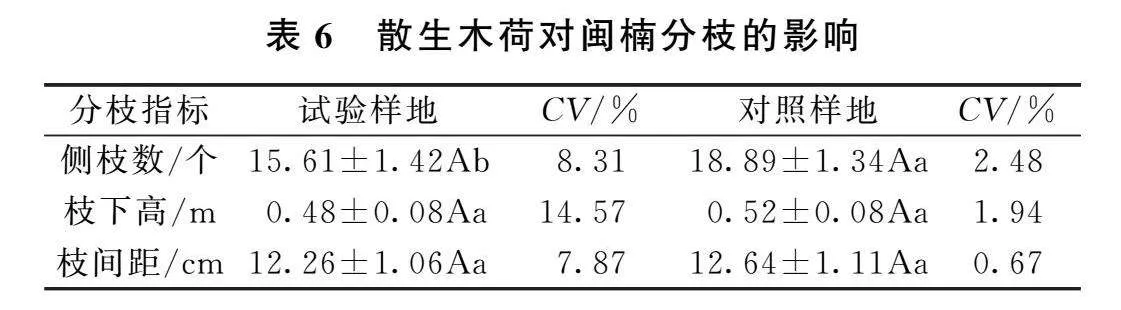
3.3散生木荷对闽楠分枝的影响
散生木荷对闽楠分枝的影响见表6、表7,试验样地内闽楠的平均侧枝数比对照样地的少3.28,差异显著;试验样地内闽楠的平均枝下高和枝间距均低于对照样地,但差异不显著。试验样地闽楠分枝指标的变异系数均大于对照样地,且差异明显,试验样地闽楠侧枝数、枝下高、枝间距的变异系数分别为8.31%、14.57%、7.87%,分别是对照样地的3.34、7.50、11.67倍。散生木荷对实验样地内的闽楠侧枝数有显著的抑制作用,对枝下高和枝间距有抑制,但不显著;同时造成闽楠侧枝数、枝下高和枝间距分布不均匀。
4讨论与结论
本研究结果表明,散生木荷对试验地范围内闽楠的树高、树冠长度和当年树高生长量具有极显著的抑制作用,对其地径和侧枝数具有显著的抑制作用,对冠幅、圆满度、树冠率、尖削度、枝下高、枝间距有抑制作用,但不显著。闽楠在前3年生长较缓慢,到第5年后生长逐渐加快[10],光照是影响楠木生长的重要因子[11],对闽楠幼林的生长具有显著影响[12],散生木荷的树冠遮挡了闽楠幼树阳光,光照强度的降低限制了生物量的积累[13],因此全光照条件下对照样地楠木人工林生长快、长势齐,而在散生木荷树冠附近楠木的生长较差[14]。闽楠与木荷各个器官对养分的利用均存在着不同程度的竞争,且木荷对养分的竞争力比闽楠的强,整体养分含量高于闽楠[15],加之散生木荷树大根深,而楠木尚处于幼树阶段,楠木对于养分的竞争必然处于弱势一方。在分散方式的保留木林冠下,更新幼树距离保留木越近,则其更新质量就越差[16],因此散生木荷同时造成闽楠生长分化,长势不均匀。本研究中,未来随着闽楠逐生长,林分的光照环境因素、种间关系、空间结构等因素的变化,都将对闽楠的生长产生影响。因此,需要对此进行长期的定位跟踪。
参考文献
参考文献审校尚未完成,请过10分钟后重试。
[1]杨宝君等编著.松材线虫病[M].北京:中国林业出版社,2003.
[2]潘俊忠,周奇,方晓敏等.松林改造提升对福建森林资源影响的推演分析[J].林业勘察设计,2022,(01):1-4.
[3]翁友恒.福建省松林改造与森林经营技术模式探究[J].林业勘察设计,2022,42(04):28-30.
[4]李莹,陈永滨,范辉华等.福建19种主要造林乡土阔叶树种光响应曲线特性分析[J].西部林业科学,2020,49(01):59-64.
[5]张平.主伐山场异龄阔叶树资源保护的探讨[J].防护林科技,2016,(06):103-104.
[6]缪宁,刘世荣,史作民等.强度干扰后退化森林生态系统中保留木的生态效应研究综述[J].生态学报,2013,33(13):3889-3897.
[7]龚道生.不同马尾松散生木密度对毛竹生长的影响[J].林业勘察设计,2010,(01):162-164.
[8]郝炜,姜华年,金爱武等.散生杉木对毛竹空间分布和胸径的影响[J].福建林业科技,2015,42(04):93-97+111.
[9]李茂萍,缪宁,喻泓等.青藏高原东缘红桦-岷江冷杉次生林中大径级保留木对其他林木的影响[J].植物生态学报,2014,38(12):1263-1272.
[10]符泽华.八年生闽楠人工林光合特性研究[D].中南林业科技大学,2014.
[11]王俊男,于洋洋,林柳兴等.杉木林林内光环境对楠木生长的影响[J].森林与环境学报,2018,38(04):425-430.
[12]李肇锋,潘军,王金盾等.光环境对闽楠幼树生长及其表型可塑性的影响[J].西南林业大学学报,2014,34(06):65-69.
[13]王振兴,朱锦懋,王健等.闽楠幼树光合特性及生物量分配对光环境的响应[J].生态学报,2012,32(12):3841-3848.
[14]吴载璋,陈绍栓.光照条件对楠木人工林生长的影响[J].福建林学院学报,2004,(04):371-373.
[15]林立彬.闽楠木荷混交林生长特性及养分竞争关系研究[D].中南林业科技大学,2019.
[16]Pastur G J M, Cellini J M, Lencinas M V, Barrera M, Peri P L. Environmental variables influencing regeneration of Nothofagus pumilio in a system with combined aggregated and dispersed retention. Forest Ecology and Management, 2011, 261(1): 178-186.
猜你喜欢
中国林副特产(2022年4期)2022-08-08 05:50:30
散文诗世界(2022年5期)2022-05-23 02:02:55
小读者(2021年2期)2021-03-29 05:03:48
华人时刊(2019年13期)2019-11-17 14:59:54
防护林科技(2019年12期)2019-04-14 04:01:18
文苑(2018年22期)2018-11-19 02:54:14
东方教育(2016年3期)2016-12-14 20:53:20
红领巾·萌芽(2016年1期)2016-09-10 07:22:44
花卉(2016年14期)2016-07-31 18:46:24
现代经济信息(2016年13期)2016-06-17 21:40:50
