
不同氮素形态配比对花榈木幼苗根系生长及氮代谢关键酶活性的影响
2024-12-31 00:00:00张明睿吴琴香曹基武吴小丽肖亚琴孙敏红
西北农业学报 2024年8期



摘" 要" 为探究最适于花榈木幼苗生长的氮素形态配比,采用氮素形态配比为NH+4∶NO-3=10∶0、5∶5、" 0∶10及以改良霍格兰标准营养液为对照的营养液,共4个处理,对花榈木1 a生幼苗进行处理。结果表明,处理期间氮素配比为NH+4∶NO-3=5∶5时,花榈木幼苗根系形态指标及硝酸还原酶和谷氨酰胺合成酶活性均显著大于其他处理,可知花榈木幼苗根系喜欢混合氮源,且铵硝1∶1均衡配比更利于氮素吸收和代谢,而单一氮源营养液对于花榈木幼苗生长的促进效果不佳,其中全铵营养液会抑制幼苗生长,降低硝酸还原酶活性。
关键词" 花榈木;氮素形态;硝酸还原酶;谷氨酰胺合成酶;根系形态;根系活力
doi:10.7606/j.issn.1004-1389.2024.08.012
https://doi.org/10.7606/j.issn.1004-1389.2024.08.012
收稿日期:2023-07-11" 修回日期:2023-10-20
基金项目:2023年中央国家重点野生动物保护补助资金(BH2023A001);湖南省教育厅科学研究项目(21C0156)。
第一作者:张明睿,男,硕士研究生,研究方向为观赏园艺。E-mail:15042216669 @163.com
通信作者:肖亚琴,女,本科,工程师,研究方向为森林培育。E-mail:809632466@qq.com
孙敏红,女,博士,讲师,研究方向为园艺植物栽培与营养品质研究。E-mail:sunminhongcaddie@126.com
花榈木(Ormosia henryi" Prain)为红豆属(Ormosia)常绿乔木,树姿挺拔优美,枝叶繁茂浓荫覆地,常作为优质的园林绿化树种和防火树种[1];花榈木还具有宝贵的药用价值,根、茎、叶等器官均可入药,其内富含丁香酚、苯乙醇、榄香素等活性化合物,具有祛风、消肿、活血化淤之功[2];此外,花榈木质地细密、纹理清晰的木材亦使其成为优质的用材树种[3]。然而,花榈木广泛的利用价值使其野生种群遭到了严重砍伐,受人为干扰严重;其自身繁殖能力又不高,种子休眠期长且不易发芽、苗期生长缓慢,现野生种群数量锐减,已被列为国家二级重点保护植物[4]。
氮素是影响植物生长的主要元素之一,在植物的生长过程中,尤其是幼苗时期的生长发育中具有不可替代的作用[5],如何提高作物对氮素养分的吸收利用效率一直是中国农业科学研究中的焦点和热点[6]。铵态氮(NH+4-N) 和硝态氮(NO-3-N)是两种植物主要吸收利用的氮素形态,不同作物对氮素的偏好有所差异[7],探究植物的氮素类型偏好能够更有效地提升种植效果和产量,提升产品品质。因此,科学地利用氮素配比施肥对促进花榈木苗木营养生长和提高苗木质量至关重要[8]。
植物根系的生长情况通常会影响植物吸收利用氮素营养的效率,并直接影响地上部分的生长情况[9]。乔哓燕等[10]研究表明:铵态氮和硝态氮按一定比例施用更有利于根系的吸收并促进作物生长。薛泽政等[11]在核桃幼苗的研究中发现,混合施肥时幼苗根系内氮素含量积累更快更多,对植株各器官生长的促进作用更加显著。这表明混合施用两种形态的氮素可以增强植物根系对氮素的吸收能力,对植株生长发育的促进作用更加显著[12]。
植物体内的氮素代谢过程是由一系列酶进行调控的,其中硝酸还原酶(Nitrate reductase,NR)和谷氨酰胺合成酶(Glutamine synthetase,GS)被认为是氮代谢过程的两个关键酶,耿玉辉等[13]研究表明:植物体内NR的活性通常随着氮营养水平一起增高或降低,可见硝酸还原酶活性的高低代表着植物氮代谢的强弱。金正勋等[14]对水稻GS活性的研究认为:GS是处于氮代谢中心的多功能酶,其在水稻营养生长和产量的形成过程中起到关键性作用。因此,通过研究两种酶活性的高低与变化能够显示出植物氮素代谢的强度[15]。
关于花榈木的研究多集中于根瘤菌等根际促生菌[16]和基质栽培[17]方面,而关于氮素形态的研究鲜有报道。本试验主要采取不同氮素形态配比营养液处理1 a生花榈木幼苗,通过根系形态、根系活力及氮素代谢过程中的两个关键酶(NR和GS),来探寻氮素形态对花榈木幼苗生长的影响,以此为花榈木的苗期栽培及肥水管理提供理论依据。
1" 材料与方法
1.1" 试验材料
试验所用花榈木种源为‘株洲1号’,于2021年4月筛选大小均匀、颗粒饱满,无病虫害的健康花榈木种子,进行适当催芽处理后播种在湖南省林业科学院试验林场。2022年1月筛选株高一致的无病虫害幼苗作为试验材料移栽到花盆中,花盆规格为高16 cm,上口直径22 cm,基质采用蛭石∶珍珠岩=1∶1的无营养基质,移栽后缓苗一段时间转移至湖南省森林植物园杜鹃花温室。
1.2" 试验设计
采用浇灌液体营养液的方式施肥,试验营养液设置4个处理,氮素形态配比分别为NH+4∶NO-3=10∶0(A1)、5∶5(A2)、0∶10 (A3),以及采用改良霍格兰标准营养液的对照组(A0)。营养液中总氮含量:15 mmol/L,其中NO-3由" Ca(NO3)2、KNO3和NH4NO3提供;NH+4由NH4NO3、(NH4)2SO4和NH4H2PO4提供;" 1 mmol/L 磷由K2HPO4提供;6 mmol/L K+由KNO3提供;4 mmol/L Ca2+由Ca(NO3)2提供;4 mmol/L Mg2+由MgSO4提供,以保证在氮素总量不变的基础上通过调整营养液配方来实现硝态氮和铵态氮的不同配比。各组营养液中均加入7 μmol/L硝化抑制剂双氰胺(C2H4N4)抑制硝化作用,pH 保持在6.0左右。
试验采用完全随机区组设计,共计4个处理,每个处理3次重复,共12株。试验于2022年8月中旬开始,每株花榈木幼苗通过量筒准确称取100 mL营养液进行浇灌,浇透,每隔15 d再次进行浇灌,共3次。
1.3" 测定指标及方法
1.3.1" 根系形态测定" 于浇灌营养液 0 d和" 45 d取样进行调查,因试验选用的基质为蛭石+珍珠岩,这种混合基质保水透气且质地松散,可确保取出全部根系且不造成损伤。测量前仔细清洗待测根系,去除沙砾和植物残体,使用EPSON 12000XL扫描仪对根系进行扫描,获取根系形态结构图像,并利用WinRHIZO Pro植物根系分析系统进行数据分析,测得0 d、45 d植株根系长度(cm)、根表面积(cm2)、根系体积(cm3)和根平均直径(mm)。
1.3.2" 根系活力测定" 使用甲烯蓝吸附法测定 0 d和45 d花榈木幼苗的根系活力,根据王云礼等[18]的研究,通过计算甲烯蓝溶液被根系吸附的量可以得出根系的总吸收面积,而根系的活跃部分能够将其吸附的物质进一步转移到细胞中去,从而在根系吸附达到饱和后再额外吸附一定量甲烯蓝。因此,根据再吸附的甲烯蓝溶液量可计算得出根系活跃吸收面积,并作为表示根系吸收活力的指标。
1.3.3" 生物量测定" 取不同铵硝配比营养液处理45 d的花榈木幼苗作为样品,将样品地上、地下部分离并分别放入烘干箱于105 ℃烘干,最后用天平测得植株地上、地下部生物量;再将二者对比即可得出根冠比。
1.3.4" 氮代谢关键酶活性测定" 分别取不同处理下培养0 d和45 d的花榈木幼苗,用清水迅速清洗干净,整根放于液氮中冷冻,之后放入" -80 ℃超低温冰箱保存。采用磺胺显色法测定硝酸还原酶(NR)活性;氯化铁比色法测定谷氨酰胺合成酶(GS)活性。
1.4" 数据分析
所有数据利用Microsoft office Excel 2021软件进行整理,并利用IBM SPSS statistics 24软件进行方差分析和多重比较(Duncan),显著性水平为" P<0.05。
2" 结果与分析
2.1" 不同氮素形态配比对花榈木幼苗根系生长的影响
从根系扫描结果图1中可以发现,经过" 45 d的不同营养液处理,各组花榈木幼苗根系较0 d时有明显变化。主要表现为根系长度均有一定伸长,须根明显增多。由表1可知,不同氮素形态配比处理下花榈木幼苗根系长度的增长存在显著差异(P<0.05),铵硝配比(NH+4∶NO-3=" 5∶5)的混合氮源处理A2的根系长度为313.41 cm,显著大于其他处理。硝态氮比例高于50%的处理(A2、A3、A0)的幼苗根系长度均显著大于A1处理(P<0.05),这表明一定量的硝态氮有助于侧根的伸长。
45 d时,A2处理根系表面积显著高于A1、A3处理,同时与A0对照组无显著差异,这表明混合氮源处理下花榈木幼苗的根系表面积较单一氮源处理下有明显增大,说明混合氮源有利于花榈木幼苗根系表面积的扩大。
(NH+4∶NO-3=5∶5)的根平均直径与全铵(NH+4∶NO-3=10∶0)存在显著差异(P<" 0.05),且与A3、A0处理无显著差异,这表明一定量的硝态氮有助于花榈木幼苗根平均直径的" 增长。
2.2" 不同氮素形态配比对花榈木幼苗根系活力的影响
由表2可知,不同氮素形态配比营养液处理下,花榈木幼苗根系总吸收面积和活跃吸收面积间均存在显著差异(P<0.05),且呈现出相同的规律。在不同氮素形态配比营养液处理下,花榈木幼苗的根系活力间存在显著差异。45 d时A2处理植株的根系总吸收面积和活跃吸收面积均显著高于其他处理,分别为17.57 cm2和1.513 cm2。这表明铵硝均衡配比(NH+4∶NO-3=" 5∶5)有助于提高花榈木幼苗的根系活力。加入硝态氮的A2、A3、A0处理无论根系总吸收面积还是活跃吸收面积均显著高于全铵处理A1,这表明硝态氮与根系侧根的伸长有关,也有利于提高根系活力。
2.3" 不同氮素形态配比对花榈木幼苗生物量及根冠比的影响
由表3可知,各处理的地上部生物量均性差异不显著,而地下部生物量间存在显著性差异" (P<0.05),说明不同氮素形态配比对植物根系生长发育影响较大,两者按一定比例施用更有利于根系吸收和作物生长。氮素形态配比为" (NH+4∶NO-3=5∶5)的A2处理的地上部生物量为4.90 g,地下部生物量为2.71 g,总生物量为7.60 g,分别为该组数据的最大值,这表明铵硝均衡配比的营养液可能更有利于花榈木幼苗的营养生长,可以显著提高花榈木幼苗生物量。
全铵配比(NH+4∶NO-3=10∶0)的A1处理地上、地下部和总生物量均为最小值,分别为" 3.83、1.83、5.66 g。这表明全铵配比的营养液促进花榈木幼苗生长的效果最差,并可能抑制花榈木幼苗地上、地下部的生长。
2.4" 不同氮素形态配比对花榈木幼苗根系氮代谢关键酶的影响
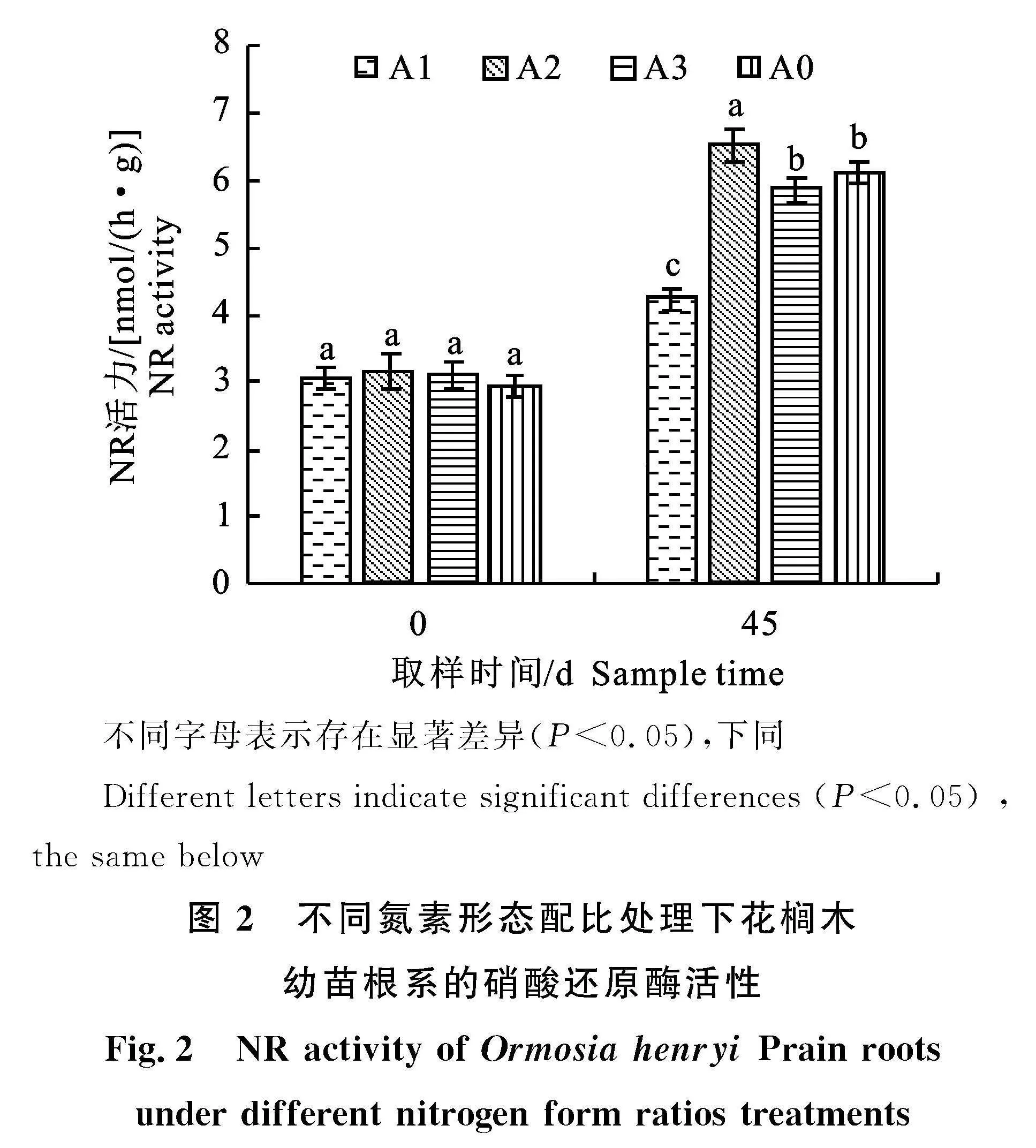
2.4.1" 硝酸还原酶(NR)" 由图2可知,45 d时A2处理,即氮素形态配比为混合氮源(NH+4∶NO-3=5∶5)时,NR活力达到最大,为6.51 nmol/(h·g),而氮素配比为全铵(NH+4∶NO-3=" 10∶0)的A1处理NR活性最低,为4.24"" nmol/(h·g)。
45 d时,4个处理的NR活性表现为A2处理显著大于其他处理(P<0.05),这表明铵硝均衡配比的氮素营养液更有利于花榈木幼苗根系NR活性的提高;全硝处理A3与对照A0处理的NR活性间不存在显著差异;全铵处理A1的NR活性显著小于其他处理,为4.24 nmol/(h·g);这表明硝态氮源对硝酸还原酶的活性表达具有一定的促进作用,而全铵处理可能会抑制硝酸还原酶活性的增长。由此可知,硝酸还原酶的活性主要受硝态氮含量调控。
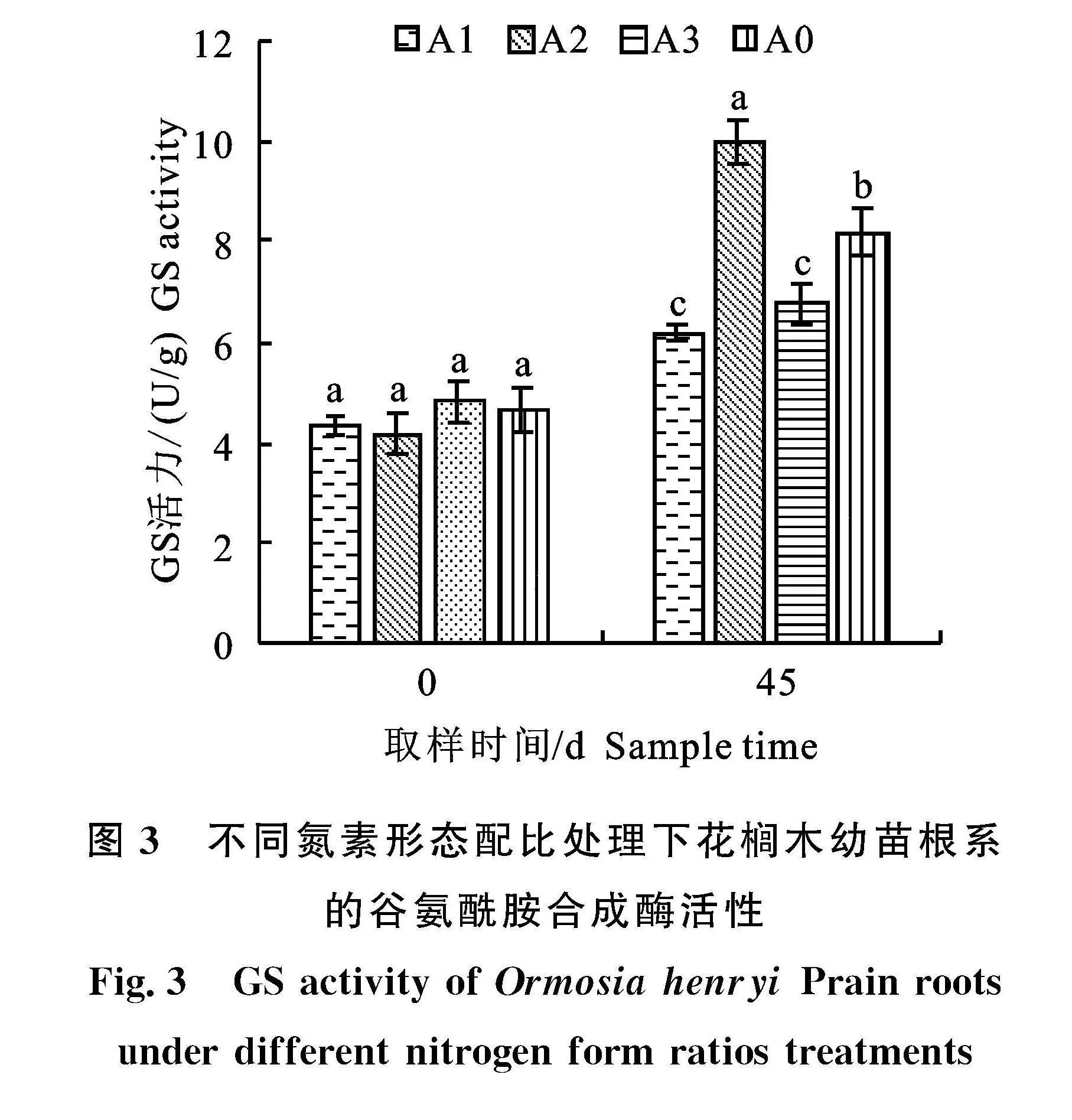
2.4.2" 谷氨酰胺合成酶(GS)" 由图3可知,花榈木幼苗叶片GS活力随氮素营养液处理时间的增长呈上升趋势,且不同氮素形态配比营养液处理下,花榈木幼苗GS活力存在显著性差异;在经过45 d的培养后,铵硝均衡配比(NH+4∶NO-3=5∶5)A2处理的GS活性达到最大,为10.02 U/g,是比" 0 d的2.4倍,并显著高于其余3个处理;对照组A0处理的GS活性虽显著小于A2处理,但其GS活性同样显著高于A1、A3处理;全铵处理(NH+4∶NO-3=10∶0)A1和全硝处理(NH+4∶NO-3=0∶10)A3之间差异不显著。
这表明了铵态氮、硝态氮混合的氮素营养液更有利于花榈木幼苗根系谷氨酰胺合成酶活性的增长,且铵硝氮源1∶1配比的A2处理的促进作用更加显著,而单一氮源的氮素营养则不利于花榈木幼苗GS活性的增长。
3" 讨" 论
3.1" 不同氮素形态配比对花榈木幼苗根系生长的影响
根系形态数据(根系长度、根表面积、根系体积、根平均直径)是衡量根系吸收营养能力的一系列综合指标。根系活力则表示植株根系对环境水分和矿质影响的吸收水平,直接影响着植株形态的发育和营养物质积累。本试验中花榈木幼苗根系形态指标和根系活力的分析结果均显示出,A2处理,即铵硝态氮配比为NH+4∶NO-3=5∶5时,花榈木幼苗的根长、根表面积、根系活力等指标大都显著高于其他处理,与前人的研究一致,与单一氮源相比,多数植物在铵态氮和硝态氮两种氮素营养共存的条件下生长会更好,且氮素利用率提高[19]。
不同氮素营养液处理下,植株生理变化是一个渐进的过程,最终结果都会表现在生物量的变化上。班甜甜等[20]的研究表明,单一铵态氮处理会导致植株根际环境酸化,阻碍根系对土壤中营养的吸收,这可能是本研究中全铵处理组的花榈木幼苗各项根系指标与其他处理组相比表现较差、生物量较低的原因之一。
3.2" 不同氮素形态配比对花榈木幼苗硝酸还原酶活性的影响
硝酸还原酶是NO-3同化步骤中的第一个酶,也是植物硝态氮代谢的一个关键酶、限速酶,在植物氮素代谢过程中起着关键的调控作用[21]。植物体吸收的NO-3首先在叶肉细胞和根系中由NR还原成NO-2,进而在各自细胞的质体中被亚硝酸还原酶还原成NH3,形成的NH3则在GS和谷氨酸合成酶(GOGAT)的催化下合成植物体所需的各种氨基酸。本研究结果表明:全铵处理下花榈木幼苗根系NR活性显著低于其他处理,说明高浓度的铵态氮可能对根系NR活性存在一定抑制作用。在铵硝氮源配比为1∶1时,花榈木幼苗根系NR活性显著高于其他处理,与张朋等[22]在杭白菊上的研究一致:杭白菊根系的NR活性在NH+4-N∶NO-3-N=1∶1时达到最大值,这表明混合氮源提高了植株对NO-3的同化能力。试验结果还显示,随着营养液中NO-3比例的增高,花榈木幼苗根系的NR活性表现出先增高后降低的趋势,这可能是高NO-3环境抑制了根系中NO-3通过木质部向叶片的运输所导致的[23]。
本研究发现花榈木幼苗根系的NR活性受到环境中硝态氮含量的调控,且会因高浓度的铵态氮产生抑制。铵硝均衡的氮素配比更有利于提高花榈木幼苗的NR活性。近年来有关硝酸还原酶活性的研究多集中于基因层面,植物体内存在着多种硝酸还原酶基因,不同的基因对NR活性表达的贡献率也有所不同,在分子水平上寻找花榈木NR活性的变化规律,有待进一步的研究与论证。
3.3" 不同氮素形态配比对花榈木幼苗谷氨酰胺合成酶活性的影响
谷氨酰胺合成酶是NH+4同化过程中的主要酶之一。戴廷波等[24]对小麦的研究中发现,与单一NO-3营养相比,增加NH+4营养的植株净光合速率、可溶性糖含量明显增加,且可维持较高的GS活性。李彩凤等[25]在甜菜上的发现证明:甜菜块根的GS活性在NH+4比例小于NO-3时,随NH+4比例增大而增加,当NH+4∶NO-3=1∶1时活性最大,当NH+4比例大于NO-3时,活性下降。本试验中,混合氮源配比的A2、A0处理的GS活性显著高于单一氮源的A1、A3处理,且铵硝氮源配比为1∶1的A2处理的GS活性显著高于对照A0处理。可以发现,NH+4比例的提高可以增加GS活性,这可能与较高的净光合速率密切相关,虽然根系同化NH+4需要消耗大量的碳水化合物,但叶片光合产物的增加保证了根系对营养物质的需求,这就使得在NH+4同化过程中起主要作用的GS的活性得到了提高,这可能是植株对NH+4营养适应机制的一部分;同时,全铵处理反而不利于植株生长和酶活性的提高,持续单一的高NH+4环境会导致植株中大量形成NO-2和NH+4等中间产物,对植物细胞进行毒害。因此,铵硝混施比单施的效果更好,尤其是铵硝均衡的氮素配比更有利于提高花榈木幼苗的GS活性。
4" 结" 论
探究植物对氮素形态配比的偏好能够更有效地提升植物营养吸收效率,利于植物生长。本研究发现,铵硝1∶1的均衡氮素形态配比促进花榈木幼苗根系生长的效果最好,而单一铵态氮或硝态氮处理对于花榈木幼苗根系生长的效果均不佳,其中全铵处理会抑制花榈木幼苗根系生长。综上所述,在实际生产种植花榈木幼苗中应注意采用混合氮源类型的氮素肥料。
参考文献" Reference:
[1]" 桂" 平,龙" 鹏.珍稀树种花榈木研究进展[J].贵州农业科学,2021,49(7):98-106.
GUI P,LONG P.Research progress on rare tree species of Ormosia henryi[J].Guizhou Agricultural Sciences,2021, 49(7):98-106.
[2]YADAV V K,MISHRA A.In vitro amp; in silico study of hypoglycemic potential of Pterocarpus marsupium heartwood extract[J].Natural Product Research,2018,33(22):1-5.
[3]刘" 鹏,何万存,黄小春,等.花榈木研究现状及保护对策[J].南方林业科学,2017,45(3):45-48.
LIU P,HE W C,HUANG X CH,et al.Research status and conservation strategy of Ormosia henryi[J].South China Forestry Science,2017,45(3):45-48.
[4]孟宪帅,韦小丽.濒危植物花榈木野生种群生命表及生存分析[J].种子,2011,30(7):66-68.
MENG X SH,WEI X L.Life table and surival analysis on ormosia wild endangered populations[J].Seed,2011, 30(7):66-68.
[5]戴良香,张智猛,张冠初,等.氮肥用量对花生氮素吸收与分配的影响[J].核农学报,2020,34(2):370-375.
DAI L X,ZHANG ZH M,ZHANG G CH,et al.Effects of nitrogen application on nitrogen uptake and distribution in peanut[J].Journal of Nuclear Agricultural Sciences, 2020,34(2):370-375.
[6]刘" 磊,宋娜娜,齐晓丽,等.水稻根系特征与氮吸收利用效率关系的研究进展[J].作物杂志,2022(1):11-19.
LIU L,SONG N N,QI X L,et al.Research advances on the relationship between root characteristics and nitrogen uptake and utilization efficiency in rice[J].Crops,2022(1):11-19.
[7]ONDOUA R N,WALSH O.Varietal differences in nitrogen use efficiency among spring wheat varieties in Montana[J].Crops Soils,2017,50(5):40-42.
[8]LIU Z,LI W,XU Z,et al.Effects of different nitrogen forms and concentrations on seedling growth traits and physiological characteristics of" Populus simonii×P.nigra[J].Journal of Forestry Research,2022,33(5):1593-1606.
[9]XU G,JIANG M,LU D,et al.Nitrogen forms affect the root characteristic,photosynthesis,grain yield,and nitrogen use efficiency of rice under different irrigation regimes[J].Crop Science,2020,60(5):2594-2610.
[10]" 乔晓燕,吴" 燕,高青海.氮素形态及配比对栝楼幼苗生长和氮代谢的影响[J].井冈山大学学报(自然科学版),2021,42(6):46-51.
QIAO X Y,WU Y,GAO Q H.Effect of nitrogen forms and ratios on the growth and nitrogen metabolism of" Trichosanthes kirilowii seedlings[J].Journal of Jinggangshan University(Natural Science),2021,42(6):46-51.
[11]薛泽政,王世伟,丁俊杰,等.不同氮素形态配比对核桃幼苗根系形态及氮素吸收的影响[J].山东农业大学学报(自然科学版),2021,52(5):759-763.
XUE Z ZH,WANG SH W,DING J J,et al.Effects of different nitrogen forms on root morphology and nitrogen uptake of walnut seedlings[J].Journal of Shandong Agricultural University(Natural Science Edition), 2021, 52(5):759-763.
[12]屈时银.不同形态氮素对红豆杉幼苗生长和生理特性的影响[J].绿色科技,2023,25(1):120-122,127.
QU SH Y.Effects of different nitrogen forms on growth and physiological characteristics of" Taxus Linn.seedlings[J].Journal of Green Science and Technology, 2023, 25(1):120-122,127.
[13]耿玉辉,李" 刚,曹秀艳,等.氮、钾不同营养水平对春玉米氮代谢的影响[J].玉米科学,2009,17(6):101-104.
GE Y H,LI G,CAO X Y,et al.Effects of different N and K levels on nitrogen metabolism of spring maize[J].Journal of Maize Sciences,2009,17(6):101-104.
[14]金正勋,钱春荣,杨" 静,等.水稻灌浆成熟期籽粒谷氨酰胺合成酶活性变化及其与稻米品质关系的初步研究[J].中国水稻科学,2007,21(1):103-106.
JIN ZH X,QIAN CH R,YANG J,et al.Changes of activity of glutamine synthetase during grain filling and its influence on rice grain quality[J].Chinese Journal of Rice Science,2007,21(1):103-106.
[15]张" 露,陈书融,吴龙龙,等.减施氮肥和增氧灌溉对水稻氮代谢关键酶活性及氮素利用的影响[J].农业工程学报,2022,38(9):81-90.
ZHANG L,CHEN SH R,WU L L,et al.Effects of nitrogen fertilizer reduction and oxygen-enhancing irigation on the key enzyme activities of nitrogen metabolism and nitrogen utilization in rice[J].Transactions of the Chinese Society of Agricultural Engineering,2022,38(9):81-90.
[16]朱天才,周洁尘,段" 翔,等.花榈木根际促生菌的筛选鉴定及促生特性[J].中南林业科技大学学报,2023,43(1):43-49.
ZHU T C,ZHOU J CH,DUAN X,et al.Screening and identication of plant growth-promoting rhizobacteria (PGPR) in the rhizosphere of Ormosia henryi and their growth-promoting characteristics[J].Journal of Central South University of Forestry amp; Technology,2023, 43(1):43-49.
[17]田" 鸿,韦小丽,罗" 旋.基质配比对花榈木容器苗生长及生理生化的影响[J].东北林业大学学报,2022,50(2):" 1-5.
TIAN H,WEI X L,LUO X.Effects of substrate ratios on growth and photosynthetic physiology of Ormosia henryi Prain containerized seedlings[J].Journal of Northeast Forestry University,2022,50(2):1-5.
[18]王云礼,陈香艳,唐洪杰.不同玉米品种的根系活力与产量性状关系的初步研究[J].农业科技通讯,2015,520(4):68-72.
WANG Y L,CHEN X Y,TANG H J.Preliminary study on the relationship between root activity and yield traits of different corn varieties[J].Bulletin of Agricultural Science and Technology,2015,520(4):68-72.
[19]OCHIENG I O,GITARI H I,M,OCHOGE B,et al.Optimizing maize yield,nitrogen efficacy and grain protein content under different N forms and rates[J].Journal of Soil Science and Plant Nutrition,2021,21(3):1867-1880.
[20]班甜甜,李晓慧,马" 超.不同形态氮肥对黄瓜生长和营养元素吸收、利用的影响[J].分子植物育种,2021,22(2):526-634.
BAN T T,LI X H,MA CH.Effects of different nitrogen forms on Cucumis sativus L.growth and nutrient elements absorption and utilization[J].Molecular Plant Breeding,2021,22(2):526-634.
[21]巩金壮,焉学倩,陈艳阳,等.氮素形态对人参氮代谢相关酶活性的影响[J].吉林农业大学学报,2023,45(6):709-715.
GONG J ZH,YAN X Q,CHEN Y Y,et al.Effects of nitrogen forms on enzyme activities related to nitrogen metabolism in Panax qinseng[J].Journal of Jilin Agricultural University,2023,45(6):709-715.
[22]张" 朋,王康才,成明超,等.氮素形态对杭白菊生长及品质的影响[J].中国中药杂志,2014,39(17):3263-3268.
ZAHNG P,WANG K C,CHENG M CH,et al.Effects of nitrogen form on growth and quality of Chrysanthemums morifolium[J].China Journal of Chinese Materia Medica,2014,39(17):3263-3268.
[23]隋" 利,王康才,易家宁.不同氮素形态对紫苏生长及品质的影响[J].土壤通报,2018,49(3):667-672.
SUI L,WANG K C,YI J N.Effect of different nitrogen forms on growth and quality of" Perilla frutescens (L.) Britt[J].Chinese Journal of Soil Science,2018,49(3):667-672.
[24]戴廷波,曹卫星,孙传范,等.增铵营养对小麦光合作用及硝酸还原酶和谷氨酰胺合成酶的影响[J].应用生态学报,2003,14(9):1529-1532.
DAI T B,CAO W X,SUN CH F,et al.Effect of enhanced ammonium nutrition on photosynthesis and nitrate reductase and glutamine synthetase activities of winter wheat[J].Chinese Journal of Applied Ecology,2003,14(9):1529-1532.
[25]李彩凤,马凤鸣,赵" 越,等.氮素形态对甜菜氮糖代谢关键酶活性及相关产物的影响[J].作物学报,2003,29(1):128-132.
LI C F,MA F M,ZHAO Y,et al.Effects of nitrogen forms on key enzyme activities and related products in sugar and nitrogen metabolism of sugar beet( Beta vulgaris L.) [J].Acta Agronomica Sinica,2003,29(1):128-132.
Effects of Different Nitrogen Forms Ratios on Root Growth and
Activities of Key Enzymes in Nitrogen Metabolism in Ormosia henryi" Prain Seedlings
ZHANG Mingrui1,2,WU Qinxiang1,2,CAO Jiwu1,2,WU Xiaoli2,XIAO Yaqin2 and" SUN Minhong1,2
(1.Research Center for Conservation Engincering of Ancient and Famous Trees,Central South University of
Forestry amp; Technology,Changsha" 410004, China; 2.Hunan Academy of Forestry,Changsha" 410004, China)
Abstract" To determine the optimum nitrogen ratio for the growth of Ormosia henryi" Prain seedlings,a modified Hoagland standard nutrient solution was used as the control,four treatments were set in this study using annual seedlings of Ormosia henryi Prain,treated with the nutrient solution with NH4+∶ NO-3=10∶0,5∶5,0∶10.The results showed that during the treatment period,when the nitrogen ratio was (NH+4∶NO-3=5∶5)" the root morphological indexes and the activities of NR and GS of Ormosia henryi" Prain seedlings were significantly higher than those in the other treatments.This suggests that Ormosia henryi" Prain seedlings exhibit a preference for mixed nitrogen sources,and a balanced ratio of ammonium nitrate 1∶1 is more conducive to nitrogen absorption and metabolism.In contrast,the single nitrogen source nutrient solution has a limited effect on the growth of Ormosia henryi" Prain seedlings.The total ammonium nutrient solutions can inhibit seedling growth and reduce nitrate reductase activity.
Key words" Ormosia henryi Prain; Nitrogen forms; Nitrate reductase; Glutamine synthetase; Root morphology; Root activity
Received"" 2023-07-11""" Returned" 2023-10-20
Foundation item" 2023 Central State Key Wildlife Protection Subsidy Fund(No.BH2023A001); Science Research Projectof Education Department of Hunan Province(No.21C0156).
First author" ZHANG Mingrui,male,master student.Research area:ornamental horticulture." E-mail:15042216669 @163.com
Corresponding"" author" XIAO Yaqin,female,undergraduate,engineer.Research area:forest cultivation.E-mail:809632466@qq.com
SUN Minhong,female,Ph.D,lecturer.Research area:horticultural plant cultivation and nutritional quality.E-mail:sunminhongcaddie@126.com(责任编辑:史亚歌" Responsible editor:SHI Yage)
