
滇山茶不同品种花表型性状的多样性分析
2024-12-31 00:00:00肖红星闵嘉晨吴学琴黄荻陈盛通陈龙清耿芳
中南林业科技大学学报 2024年10期
摘 要:【目的】通过分析38个滇山茶Camellia reticulata品种的12个花表型性状在数量、大小和形状三个维度上的分布和差异,探讨滇山茶不同品种花表型变异的(横向和纵向)规律,为滇山茶新种质的选育和高效利用提供参考。【方法】对滇山茶的12个花表型性状进行方差分析、相关性分析、主成分分析、综合分析和聚类分析,箱线图和应用频率分布函数等方法进行分析。【结果】滇山茶不同品种花的12个表型性状存在丰富的变异,变异系数为13.00%~67.00%,平均变异系数为27.24%;变异系数法的权重计算结果显示,雌蕊数(20.485%)的变异性影响最大,花瓣长宽比(3.99%)的变异性影响最小;相关性分析结果表明,12个性状间都存在极显著相关关系,花径与花瓣宽度相关性最大(r=0.822);主成分分析结果表明,前4个主成分提取到的特征值都均大于1.00,累计贡献率达到82.129%;通过聚类分析将其分为3大类。综合评价结果表明‘大理茶’的综合得分最高(F=3.230),在38个滇山茶品种中综合特性最好。对聚类分析中的3类滇山茶进行箱形图和频率分布函数分析,其中9个性状表现出显著差异,其余3个性状无显著差异,第Ⅰ类各性状分布较向上,第Ⅱ类分布较为居中,第Ⅲ类分布较向下。而在频率分布函数分析中,各类的性状均表现出明显的偏置,变化程度依次为数量相关特征>尺寸相关特征>形状相关特征。【结论】滇山茶的12个花表型性状存在丰富的变异现象,主要以雌雄蕊群逐渐瓣化为演化方向,研究结果确定了滇山茶花表型演化的趋势,为后期滇山茶定向育种提供一定的理论依据。
关键词:滇山茶;花表型性状;不同品种;多样性分析
中图分类号:S794.4 文献标志码:A 文章编号:1673-923X(2024)10-0181-15
基金项目:国家自然科学基金项目(32260416);云南省教育厅科学研究基金项目(2022J0520);云南省农业基础研究联合专项项目(202101BD070001-095);云南省“兴滇英才支持计划”青年人才专项项目(YNQR-QNRC-2019-104)。
Diversity analysis of flowers phenotypic traits of Camellia reticulata cultivars
XIAO Hongxinga,b,c, MIN Jiachena,b,c, WU Xueqina,b,c, HUANG Dia,b,c, CHEN Shengtonga,b,c, CHEN Longqinga,b,c, GENG Fanga,b,c
(a.College of Landscape Architecture and Horticulture; b. Southest Landscape Architecture Engineering Technology Research Center, National Forestry and Grassland Administration; c. Yunnan Functional Flower Resources and Industrialization Technology Engineering Research Center, Southwest Forestry University, Kunming 650224, Yunnan, China)
Abstract:【Objective】Phenotypic variation (including directions and degrees) of 12 floral traits from 38 cultivars of Camellia reticulata were explored by comparing their distributions and deviations in three different dimensions: floral organ number, size, and the shape.【Method】Analysis of variance, correlation analysis, principal component analysis, comprehensive analysis, cluster analysis, as well as box-plot and application of frequency distribution functions were conducted on the 12 floral phenotypic traits of C. reticulata.【Result】There were rich variations in 12 floral traits of different cultivars, with a coefficient of variation (CV) ranging from 13.00% to 67.00%, an average CV of 27.24%;The weight calculation using the CV showed that the variability of pistil number (20.485%) had the greatest impact, while the variability of petal length /petal width (3.99%) had the smallest impact; Correlation analysis showed that there was a highly significant correlation among all 12 traits, with the greatest correlation between flower diameter and petal width (r=0.822). Principal component analysis showed that the feature values extracted from the first four principal components were all greater than 1.00, with a cumulative contribution rate of 82.129%; Cluster analysis divided 38 cultivars into three groups. The comprehensive evaluation indicated that ‘Dali Cha’ received the highest comprehensive score (F=3.230), making it the best among the 38 cultivars of C. reticulata in terms of comprehensive characteristics. The three groups of 38 cultivars were also analyzed by using box plots and frequency distribution functions. There were only nine traits showed significant differences. Group I of floral traits all showed upward distributed box bodies in box plot analysis and Group III was downward distributed. However, all the 12 traits in three groups exhibited significant differences in frequency distribution functions. The variation degrees were quantitatively characterized by numbering traits>sizing traits>shaping traits. 【Conclusion】The 12 floral phenotypic traits of C. reticulata exhibit rich variability, primarily evolving towards gradual petalization of the stamen and pistil groups. The research findings establish the trend of floral phenotype evolution in C. reticulata, providing a theoretical basis for targeted breeding of C. reticulata in the future.
Keywords: Camellia reticulata; floral phenotypic traits; different cultivars; diversity analysis
滇山茶Camellia reticulata,属于山茶科Theaceae山茶属Camellia,常绿乔木或灌木,其观赏性极高,品种丰富多彩、形态变异多样、花型丰富多样、花色艳而不妖[1-2],位居云南八大名花之首,同时也是云南省的省花和昆明市、楚雄市和腾冲市的市花[3-4]。滇山茶是西部地区尤其是云南特有的木本观花植物,但其应用没有体现资源优势。首先,据世界山茶属品种注册中心记载(截至2023年11月),滇山茶的品种仅有841个品种,远远低于位居首位的华东山茶C. japonica(18 271个品种)和茶梅C. sasanqua(1 334个品种)。其次,花色是滇山茶重要的观赏性状[5],但滇山茶花色种类相比于其他木本观花植物,如月季[6-7]、杜鹃花[8]等的花颜色比较单一,主要以红色系为主,稀有浅色和白色花,更无黄色、蓝色及其他颜色的品种;花型的变异包括单瓣型、半重瓣型、重瓣型,以重瓣型和半重瓣型为主,稀有单瓣型[9-10]。如今,滇山茶虽经过长达1500多年的自然杂交和人工栽培选育,已育出形态变异多样,色彩丰富的品种[11-12],但这些品种的形态变异规律及演化趋势目前鲜有报道。本研究将对滇山茶不同品种的花表型性状特征进行分析,以探索滇山茶花表型演化的趋势,为后期滇山茶定向育种提供一定的理论依据。
表型性状分析是基于形态学对植物性状进行分析,为植物的育种研究及分子生物学等方面提供一定的研究基础[13]。其花表型性状是评价植物种质资源的重要指标,被认为是不同分类群复杂表型鉴定以及评价进化与发育偏差相互作用的传统性状[14],由于花形态特征所获得的信息是由大量的定性和定量性状数据集组成的,因此表型多样性分析被认为是对其进行评价的最合适的分析工具[15-16]。目前国内外山茶属植物表型研究的报道多集中在杜鹃叶红山茶C. azlea[17-18]、金花茶C. nitidissima[19]、香花油茶C. osmantha[20]、西南红山茶C. pitardii[21]等。而关于滇山茶的研究主要集中在对花型进行遗传多样性、遗传关系的AFLP分析[22]和腾冲红花油茶C. reticulata f. simplex花芽分化及雌配子体发育的形态学观察[23],对花表型性状研究较少。
本研究通过对选定38个健壮生长、性状稳定的滇山茶品种的花器官表型性状进行分析,探讨38个滇山茶品种12个花器官的表型变异:1)确定3个类群在花器官数量、大小和形状3个不同维度上花器官变化的方向和程度;2)比较不同分析方法对生成变异规律的影响。从而分析滇山茶品种的形态学多样性,以获得滇山茶品种花表型性状的现状及变异的规律,为后期滇山茶定向育种提供理论依据及分子生物学等方面提供一定的研究基础。
1 材料与方法
1.1 试验材料
本试验以昆明市植物园的山茶园(102°75′E,25°14′N,海拔1 992 m)和昆明市宜良县正扬茶花园林绿化有限公司(103°23′E,24°88′N,海拔1 950 m)的健壮滇山茶品种为研究对象,选取生长状况良好、健康无病害且处于盛花期(中期)的38个滇山茶品种的花(表1)作为试验材料。
1.2 研究方法
本研究综合参照中国茶花品种分类、测试指南及已知品种数据库[24],以及茶树种质资源描述规范和数据标准[25]。本研究对每个滇山茶品种随机选择4株植株,从每株茶花的中部以上分支采集4朵典型、标准一致的盛花期花,共计16朵,取其平均值进行统计分析。然后,立即观察并记录所采每朵茶花的12个花部形态学指标,包括3个数量特征(X)、7个尺寸特征(C)和2个形状特征(S)(图1、表2)。
研究方法包括三个组成部分:一是MS(测量调查),即对研究对象滇山茶花的各个器官的个体测量的收集,主要是数量性数据的收集;二是VG(视觉观察调查),即对研究对象滇山茶花的各个器官的整体视觉观察,主要是捕捉定性或视觉特征;三是结合了VS(具体观察调查),即对研究对象滇山茶花的各个器官的个体观察。

1.3 数据分析



箱线图的绘制采用以下方式:箱体覆盖了观察区域的50%,上下横线之间的区域覆盖了观察区域的90%,箱外的数值被视为异常值,本研究的重点在于箱体和均值。平均值用于邓肯多重比较检验确定3大类之间的差异显著性(P<0.05和P<0.01分别被视为显著和极显著),异常值则用于反映超越信息[31],这对于滇山茶种质创新非常重要。频率分布函数分析使用Excel 2019、Origin 2022和SPSS 22.0软件,拟合12个花器官表型性状的频率分布函数,探讨变异规律(包括方向和程度),12个花部表型性状均服从正态函数分布。
2 结果与分析
2.1 滇山茶不同品种花表型性状间的多样性分析
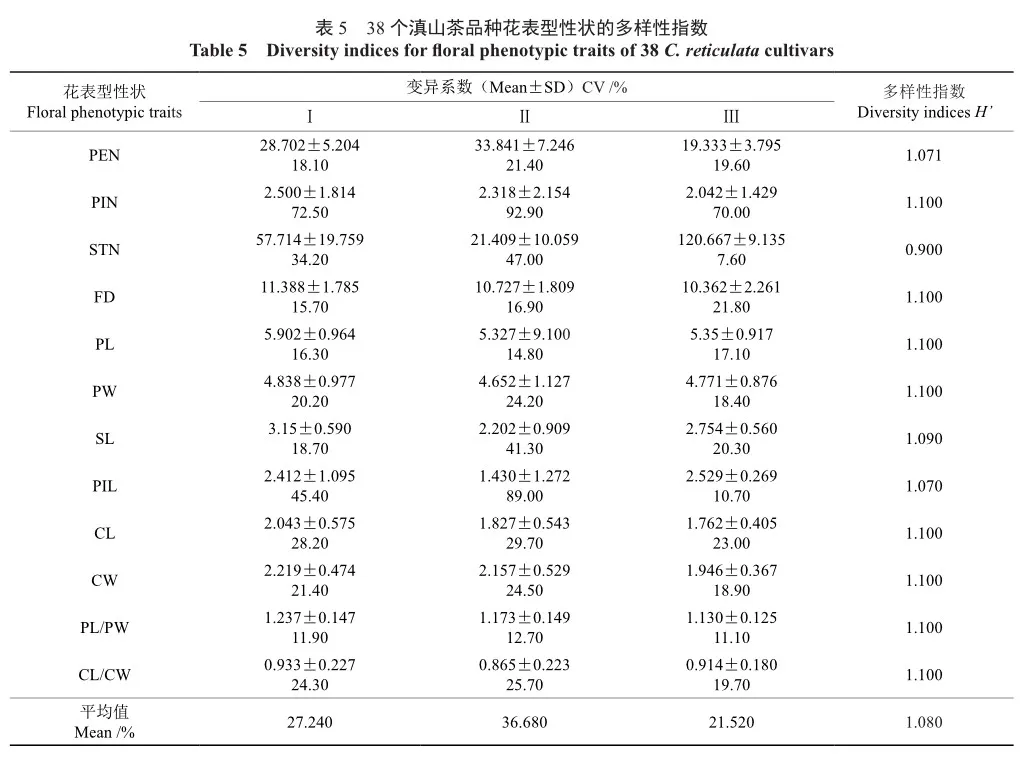
采用F检验分析滇山茶不同品种间的差异(表3),12个花表型性状在38个滇山茶品种间都存在极显著差异。12个花表型性状的多样性指数范围为0.40~2.20。其中,雄蕊数的多样性指数最高为2.20,其次为花瓣数和花径,多样性指数分别为2.18和2.02。而雌蕊长度的多样性指数最低为0.40;其次是萼片长度和花瓣长宽比,多样性指数分别为0.51和0.56。12个花表型性状的变异系数范围为13.00%~67.00%,平均变异系数为27.24%,说明38个滇山茶品种在花表型性状间存在较大程度的变异。涉及12个花表型性状变异系数高低依次为雌蕊数(67.00%)>雄蕊数(42.60%)>雌蕊长度(42.00%)>萼片长度(27.10%)>萼片长宽比(23.70%)>花瓣宽度(20.90%)>花瓣数(20.10%)>萼片宽度(19.90%)>雌蕊长度(19.80%)>花瓣长度(15.70%)>花径(15.10%)>花瓣长宽比(13.00%)。说明各性状中花瓣长宽比(13.00%)相对比较稳定,而雌蕊数(67.00%)变异程度最大。

变异系数法的权重计算结果显示(图2),涉及12个花表型性状变异系数权重高低依次为雌蕊数(20.485%)>雄蕊数(13.022%)>雌蕊长度(12.851%)>萼片长度(8.296%)>萼片长宽比(7.254%)>花瓣宽度(6.390%)>花瓣数(6.148%)>萼片宽度(6.100%)>雄蕊长度(6.056%)>花瓣长度(4.802%)>花径(4.606%)>花瓣长宽比(3.990%),其中指标权重最大值为雌蕊数(20.485%),最小值为花瓣长宽比(3.990%)。总体而言,雌蕊数、雄蕊数和雌蕊长度的变异性在涉及12个花表型性状影响较大,而其余性状的影响最小,但仍然对滇山茶的花表型和生殖特征有一定的影响。
2.2 滇山茶不同品种花表型性状间相关性分析
38个滇山茶品种的12个花表型性状相关性分析结果(图2)表明,12个花表型性状间都存在极显著相关关系(P<0.05),其中雌蕊数与雄蕊数、雄蕊长度、雌蕊长度存在极显著负相关,与花瓣长宽比存在显著正相关;花瓣数除了与雌蕊数和雄蕊数无显著性相关性外,与其他性状都存在显著性正相关或负相关;雄蕊数与雌蕊长度和雄蕊长度存在极显著正相关;花径和花瓣长度除了与雌蕊数、雄蕊数、花瓣长宽比和萼片长宽比无显著性相关性外,与其他性状都存在极显著正相关;花瓣宽度花径除了与雌蕊数、雄蕊数和萼片长宽比无显著性相关性外与其他性状都存在极显著正相关;雄蕊长度与雌蕊长度、萼片长度、萼片宽度存在极显著正相关;雌蕊长度与萼片长度、萼片宽度、萼片长宽比存在极显著正相关;萼片长度与萼片宽度、萼片长宽比存在极显著正相关;萼片长度与花瓣长宽比和萼片长宽比存在显著负相关。这些结果对于了解滇山茶的花部特征及其遗传基础具有一定的重要性。

2.3 滇山茶不同品种花表型性状主成分分析
对38个滇山茶品种的12个花表型性状进行主成分分析,结果如表4所示,前4个主成分提取到的特征值均大于1.00,累计贡献率达到82.129%,表明这4个主成分能够反映12个表型性状的大部分信息。其中,第1主成分的特征值最大,为4.992,对应的特征向量中花瓣长度的绝对值最大,为0.854,贡献率达到41.602%。第2主成分的特征值为1.871,对应的特征向量中雄蕊数的绝对值最大,为0.657,累计贡献率达15.595%。第3主成分的特征值为1.724,对应的特征向量中萼片长宽比的绝对值最大,为0.595,贡献率达14.365%。第4主成分的特征值最小,为1.268,对应的特征向量中花瓣长宽比的绝对值最大,为0.839,贡献率达10.567%。这些结果揭示了前4个主成分在总体上捕捉了大部分表型性状的变异情况。

2.4 滇山茶不同品种花表型性状聚类分析
对38个滇山茶品种的12个花表型性状进行聚类分析,在欧氏距离(Euclidean distance)为15.0处将38个滇山茶品种资源分为3大类(图3)。
本研究中3大类的表型性状多样性指数(H′)变化范围为0.90~1.10,平均变异系数分别为27.24%、36.68%和21.52%,说明这3大类的花表型具有丰富的遗传变异。
第Ⅰ类包含‘楚雄茶’、‘靖安茶’、‘大理茶’(图3)、‘西林’、‘雪娇’、‘节节高’等21个滇山茶品种,与第Ⅱ类和第Ⅲ类相比(表5),该类的雌蕊数、花径、花瓣长度、花瓣宽度、雄蕊长度、萼片长度、萼片宽度、花瓣长宽比和萼片长宽比的平均值最大,仅雄蕊数、雌蕊长度和花瓣数处于中等水平。此类群平均变异系数处于中等水平,为27.24%,其中,花瓣数的变异系数最大,为72.50%,花瓣长宽比的变异系数最小为11.90%。说明该类群花较大,且瓣化程度相对较低,多为半重瓣,在观赏上具有一定优势。
第Ⅱ类包含‘恨天高’、‘童子面’、‘朱砂紫袍’(图3)、‘菊瓣’、‘赛菊瓣’、‘紫袍’等11个滇山茶品种,与第Ⅰ类和第Ⅲ类相比,该类仅花瓣数均值最大,雌蕊数、花径、萼片长度和萼片宽度均值处于中等水平,而雄蕊数、花瓣长度、花瓣宽度、雌蕊长度、雄蕊长度和萼片长宽比的均值相对较小。此类平均变异系数最大,为36.68%,其中雌蕊数的变异系数最大,为 72.50%,其次是雌蕊长度为89.00%,花瓣长宽比的变异系数最小,为11.90%。说明该类瓣化程度相对较高,多为重瓣花,在观赏上极具优势。
第Ⅲ类包含‘粉蝴蝶’、‘张家茶’、‘银粉牡丹’、‘滇池粉妍’、‘东林’和‘早桃红’(图3)共6个滇山茶品种,与第Ⅰ类和第Ⅱ类相比,该类仅雄蕊数和雌蕊长度均值最大,花瓣长度、花瓣宽度和雄蕊长度的均值处于中等水平,花瓣数、雌蕊数、花径、萼片长度、萼片宽度和花瓣长宽比的均值最小。此类平均变异系数最小,为21.52%,说明该类相比于其他2个类群,稳定性较好,其中雌蕊数的变异系数最大,为70.00%,雄蕊数的变异系数最小,为11.90%。说明该类瓣化程度相对较低,雄蕊保留较多,在结实和杂交育种上更具优势。
2.5 滇山茶不同品种花表型性状的综合评价
滇山茶品种的12个花表型性状的得分系数如表6所示,构建其4个主成分为参数的线性组合模型,并分别计算38个滇山茶品种在4个主成分中的综合系数(表7)。基于各主成分的贡献率进一步建立滇山茶不同品种的综合评价模型,用于计算38个滇山茶品种综合得分,模型为F=(41.602F1+15.595F2+14.365F3+10.567F4)/ 82.129,根据此模型对38个滇山茶品种进行综合评价。在38个滇山茶品种的综合评价中,排在前6的品种分别为‘大理茶’(3.230)、‘靖安茶’(2.980)、‘咪依噜’(1.607)、‘凤山茶’(1.471)、‘柳叶银红’(1.350)和‘宝珠茶’(1.292),都为第Ⅰ类的滇山茶品种。整体评价结果显示,类群Ⅰ的品种综合评价较好,排名比较靠前,而第Ⅱ类和第Ⅲ类的排名波动较大,结合聚类分析结果,发现花径大、雄蕊数多等特征的滇山茶排名总体靠前,而花径小、花瓣少的滇山茶品种基本都排在末位,这与滇山茶以花大色艳而著名相吻合。
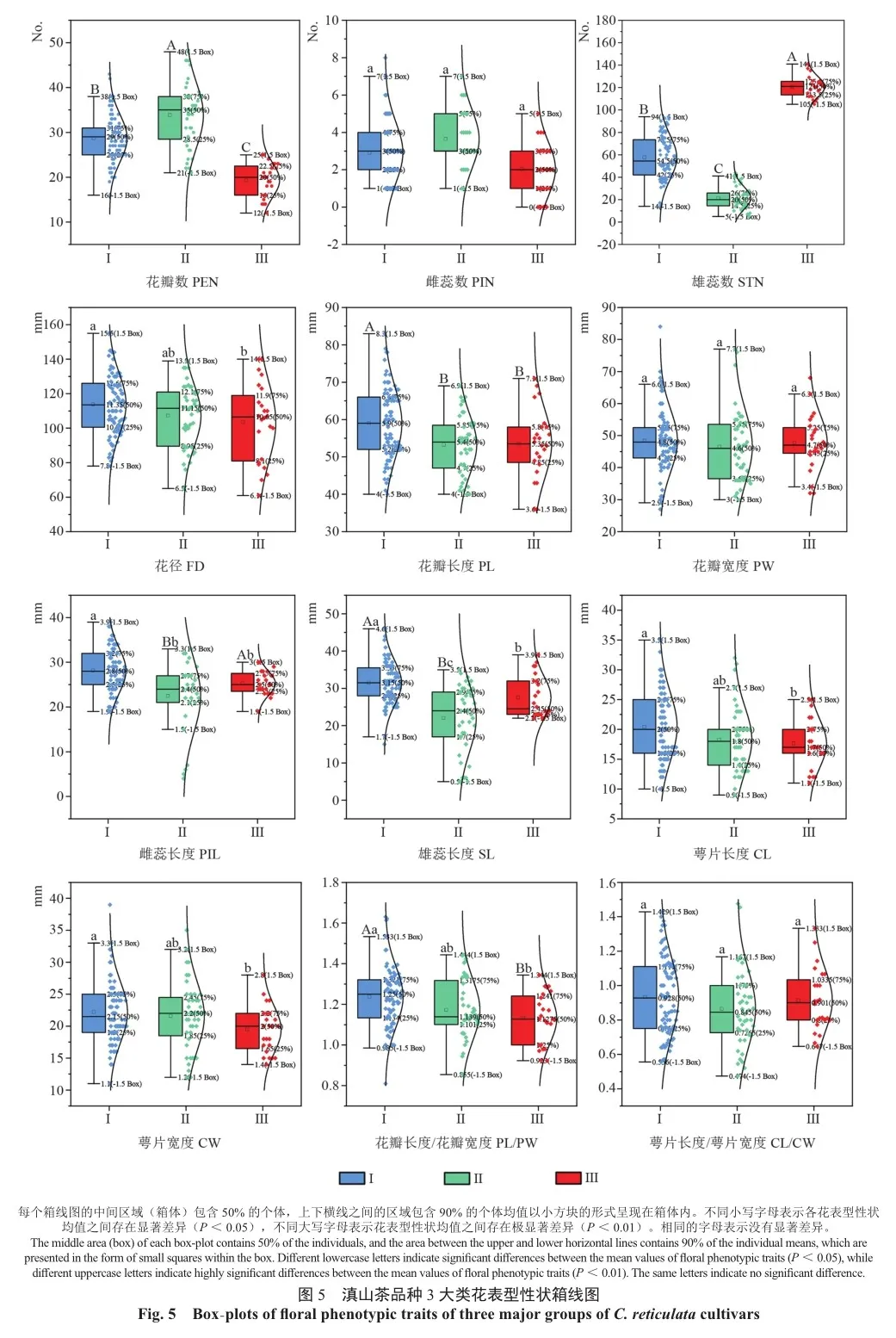
2.6 滇山茶不同品种花表型性状箱线图分析
38个滇山茶品种的12个花表型特征包括器官数量、大小和形状,通过箱线图呈现(图4)。对这些品种进行了聚类分析分成3大类,差异分析结果表明,有9个花表型性状具有显著差异,其余3个花表型性状则没有明显差异。这些发现可为滇山茶品种间花表型差异提供有力的统计支持,特别是在花瓣数、雄蕊数等方面的显著变异,有助于深入挖掘品种间的形态差异和遗传多样性。

对38个滇山茶品种聚类的3大类群进行分布比较,相比于第Ⅰ类,除花瓣数、雌蕊数、雄蕊数、花瓣宽度、萼片宽度外,第Ⅱ类和第Ⅲ类的其他花表型性状均呈向下分布。各表型性状的具体分布如下:在花器官数方面(图5),第Ⅱ类的花瓣数及第Ⅰ类的雌蕊数、雄蕊数分布范围较广,第Ⅰ类的花瓣数和雌蕊数存在一定的越界个体(比例分别为2.30%和1.20%),3大类的雄蕊数分布比较集中,但存在极显著差异,说明该性状变异性较大;雌蕊数分布范围较广,但无显著差异,说明该性状变异性较小。在花器官大小方面,第Ⅰ类的花瓣宽度、花瓣长度、雄蕊长度、雌蕊长度、萼片长度、萼片宽度和类群Ⅲ的花直径分布范围较大。其中,第Ⅰ类的花瓣宽度、雄蕊长度、萼片宽度和第Ⅱ类的雌蕊长度、萼片长度、萼片宽度存在一定的越界个体(比例分别为5.90%、1.20%、 1.20%、9.00%、6.80%和2.30%),3个类群的雌蕊长度分布较集中,且存在显著差异,说明该性状变异性较大;花瓣宽度分布范围较广,但无显著差异,说明该性状变异性较小。从花器官形状来看,第Ⅰ类的花瓣长宽比和第Ⅱ类的萼片长宽比的分布范围都较大,且存在一定的越界个体(比例分别为3.60%和4.50%)。

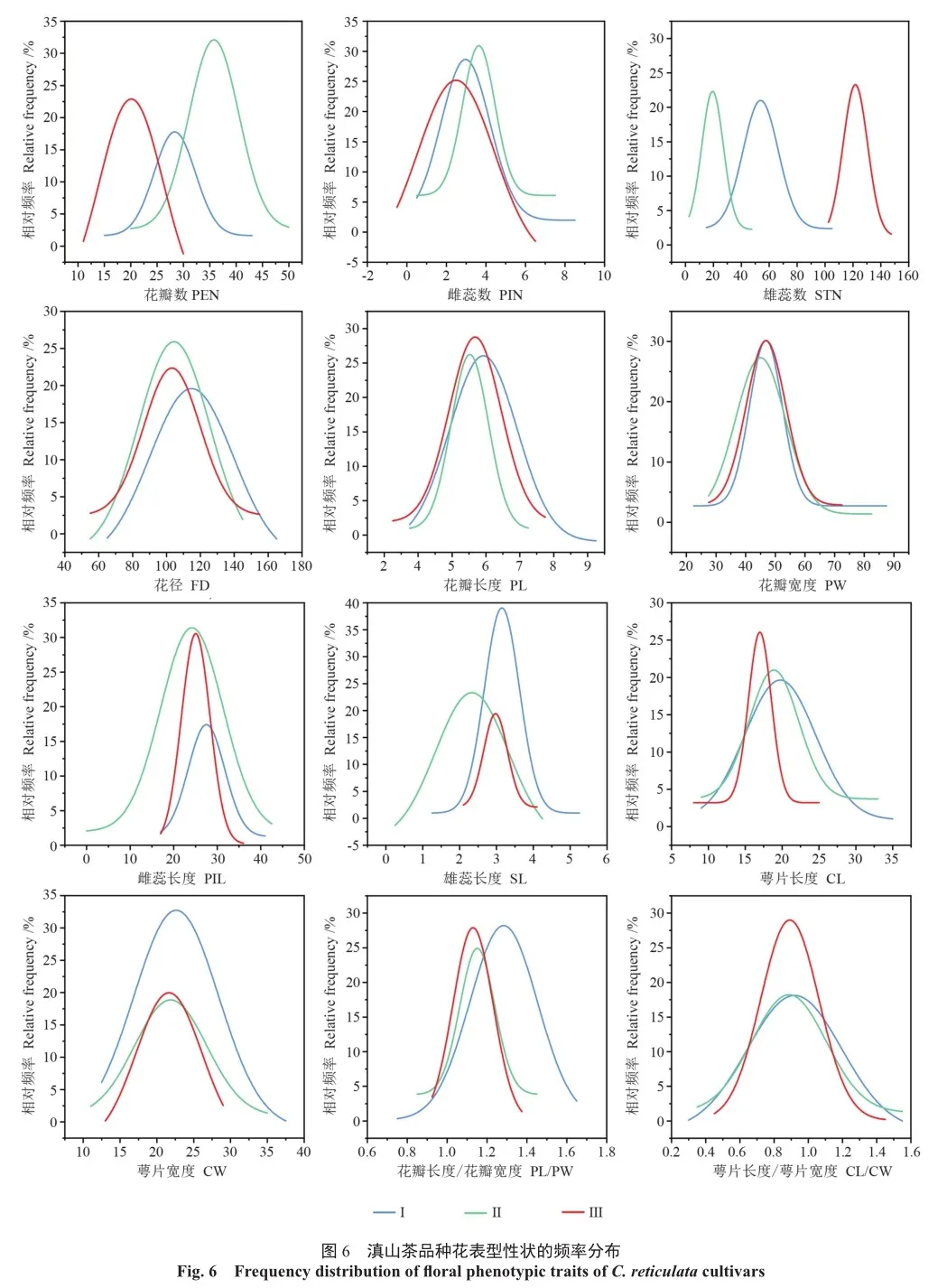
2.7 滇山茶不同品种花表型变异的频率分布函数分析
为了进一步分析滇山茶3大类花表型性状的变异情况,对12个花表型性状进行了频率分布函数拟合(图6),所有花器官表型性状均遵循正态分布(R2 =0.916 4~0.998 3)(表8)。对38个滇山茶品种的3大类进行频率分布函数比较,除雌蕊数、花瓣长度、花瓣宽度的幂分布函数比较集中之外,其余9个性状的幂函数分布较离散,进一步说明滇山茶不同品种花表型性状变异性较大。其中,在数量性状方面,3大类花瓣数和雌蕊数的正态分布函数呈右偏(增加趋势),而雄蕊数的正态分布函数呈左偏(减少趋势),这表明增加的花瓣数可能由雄蕊转化产生;在尺寸特征方面,花径、雄蕊长度、雌蕊长度和萼片长度显示出纵向和横向偏移的趋势,说明其性状变异性较大,而花瓣长度和花瓣宽度的幂分布函数没有出现明显的偏移,说明其性状变异性较小;在形状特征方面,第Ⅰ类的花瓣长宽比相比于第Ⅱ类和第Ⅲ类的正态分布函数呈右偏,说明该类群相比于其他2个类变异性较大,而萼片长宽比在横向方向没有出现明显的偏移,说明该性状变异性较小。
3 讨 论
表型性状对植物种类的鉴别、授粉生态学的研究以及植物进化和生态方面的理解具有重要意义[32]。通过对大规模表型数据的分析,可以更全面地了解基因型和表型之间的关系以及各种中间环节对性状表达的影响[33-34],有助于揭示物种内遗传多样性和遗传规律,为进化、遗传学和生物多样性研究提供重要的数据支持[35]。利用相关性、聚类以及主成分分析方法对种质资源性状遗传分析研究[36],广泛应用于育种、多样性研究及种质分类[37]。花是植物重要的组织器官,在生命周期中扮演着多重角色,既为观赏带来美感,又在植物繁殖中发挥着重要的生物学功能[17]。通过对滇山茶花表型特征的研究和分析,有助于筛选出更具优良性状的滇山茶品种,对花表型性状的改良和选育过程中的优化具有重要作用[12]。
本研究中,38个滇山茶品种的12个花表型性状的多样性指数在0.40~2.20,其中有9个花表型性状的多样性指数大于1.00,说明滇山茶品种具有较为丰富的花表型多样性,这与吴贵进等[12]的研究结果一致。变异系数是一种用于度量数据离散程度的统计指标,反映了性状变量的离散程度[38]。本研究中,12个花表型性状的变异系数范围为13.00%~67.00%,均大于10.00%,证实了滇山茶不同品种间花表型性状的变异程度较大,这与前人的研究结果相吻合[39],这也说明滇山茶品种资源多样性丰富,与辛桐[40]的研究保持一致,其中雌蕊数的变异系数(67.00%)最高,这可能与选取的材料中有些品种雌蕊数已退化有关。通过比较分析多样性指数和变异系数发现,变异系数与多样性指数的变化趋势相反,滇山茶变异系数大的性状其多样性指数相对较低,这与张海平等[41]对睡莲、丁释丰等[42]对黄心树及苏群等[43]对睡莲的研究结论相同。
相关性分析对滇山茶品种改良和生物技术研究都具有重要的指导意义[44]。本研究中,12个花表型性状间都存在极显著的相关关系,说明这12个花表性状可能在滇山茶品种的花朵的演化过程中发挥着密切的协同作用,如雌蕊和雄蕊存在瓣化,从而导致花瓣数的增加。聚类分析结果表明,第I类可作为观赏或育种优质亲本材料;第II类可用于培育新的品种,以改进观赏性和适应性等特征;第Ⅲ类滇山茶品种在某些特定用途或研究中更可靠和可预测。植物种质资源综合评价在育种研究中具有重要意义[45],综合评价结果表明,‘大理茶’的综合得分最高(F=3.230),说明该品种在38个滇山茶品种综合特性最好,结合12个花部性状,该品种相较于其他品种观赏价值高,但瓣化现象严重,不利于繁育后代。

在箱线分析中,在各类群中的花器官数量、大小和形状方面存在一定比例的越界个体。比如,在花器官数量和大小方面,某些滇山茶品种可能表现出与其所属类群不一致的特征,这可能是由于突变、特定生态条件、遗传重组或其他遗传机制引起的,可为选育新的滇山茶品种提供一定的参考。本研究在三大类进行应用频率分布函数分析中,清楚地揭示滇山茶的表型变异,其所有花器官表型性状均遵循正态分布,拟合度较高(R2为0.916 4~0.998 3),这也与箱线分析的结果保持一致,这些结果为滇山茶品种特性的理解和进一步的育种提供了重要信息,为后续的滇山茶相关形态以及机理研究提供了方向和思路。
本研究通过对38个滇山茶品种的12个花表型性状的全面分析,展现了滇山茶在花的形态特征上的丰富多样性和显著的变异性,进而为滇山茶品种的选育和生物多样性保护提供了有力的科学依据。研究揭示了38个滇山茶12个花表型性状之间的复杂关系和演化趋势,对于理解其遗传多样性和生物学特性具有重要意义。尽管如此,本研究存在局限性,主要体现在样本数量有限,未能完全捕捉到滇山茶的整体遗传和表型多样性;研究方法侧重于传统的统计分析,对于表型性状与基因型之间深层次联系的解析不足;同时,对于环境因素如生长地点、气候条件等外部影响因素考虑不够全面,这可能影响到对表型性状表达影响的全面理解。

鉴于这些局限性,未来的研究方向应当包括:1)通过扩大滇山茶品种的样本量和样本覆盖范围,增加不同生态环境下的样本,以期获得更加全面和具有代表性的数据集;2)结合现代生物技术如分子信息标记、基因组学和转录组学等方法,探讨表型性状与遗传信息之间的联系,以揭示性状变异的分子机理,特别是滇山茶花瓣化现象的研究机理,如在油茶[46-47]的应用;3)考虑到环境因素对植物表型的影响,在今后的研究中应系统评估环境变量对滇山茶表型性状表达的影响,以及可能的遗传与环境的互作效应;4)采用先进数据分析技术提升分析效率和精确度,从大规模数据中发掘更深层次的生物学信息;5)开展功能性研究,如研究特定表型性状(瓣化现象)对滇山茶观赏价值和生态适应性的影响,为滇山茶的应用和保护提供科学依据。通过这些综合性和多角度的研究,进一步深化对滇山茶遗传和表型多样性的认识,为其种质资源的保护、利用和持续育种提供更加坚实的科学基础。
4 结 论
本研究对38个滇山茶品种的12个花表型性状进行了多样性分析,结果表明38个滇山茶品种在12个花的表型性状上展现出丰富的多样性和较大的变异程度。证实了滇山茶作为一个种质资源的多样性和丰富性。综合评价结果表明,滇山茶品种‘大理茶’在综合特性上表现最佳,但其瓣化现象的存在也提示了对繁育后代可能的不利影响。箱线分析和应用频率分布函数分析结果进一步揭示了滇山茶的表型变异规律。特别值得注意的是,花表型性状间的显著相关性揭示了它们在滇山茶花朵演化中可能存在的密切协同作用,主要以雌雄蕊群逐渐瓣化为演化方向。研究结果确定了滇山茶花表型演化的趋势,为理解植物演化过程中的功能性状协同变化提供了新的视角,为深入了解滇山茶品种特性及其育种提供了宝贵信息。
参考文献:
[1] FU M, YANG X, ZHENG J, et al. Unraveling the regulatory mechanism of color diversity in Camellia japonica petals by integrative transcriptome and metabolome analysis[J]. Frontiers in Plant Science,2021,12:685136.
[2] GENG F, NIE R, YANG N, et al. Integrated transcriptome and metabolome profiling of Camellia reticulata reveal mechanisms of flower color differentiation[J]. Frontiers in Genetics,2022,13:1059717.
[3] 夏丽芳.山茶花[M].北京:中国建筑工业出版社,2002. XIA L F. Camellia flower[M]. Beijing: China Architecture and Building Press,2002.
[4] 陈世军,张明泽,姚玉仙.基于SSR标记的黔南茶树种质资源DNA指纹图谱构建[J].植物遗传资源学报,2017,18(1): 106-111. CHEN S J, ZHANG M Z, YAO Y X. Establishment of DNA fingerprinting for tea germplasm from Qiannan Prefecture by SSR markers[J]. Journal of Plant Genetic Resources,2017, 18(1):106-111.
[5] VELA P, SALINERO C, SAINZ M J. Phenological growth stages of Camellia japonica[J]. Annals of Applied Biology,2013, 162(2):182-190.
[6] 温佳辛,王超林,冯慧,等.月季花色研究进展[J].园艺学报, 2021,48(10):2044-2056. WEN J X, WANG C L, FENG H, et al. Research progress on flower color of rose[J]. Acta Horticulturae Sinica,2021,48(10):2044-2056.
[7] 尹世华,王康,黄晓霞,等.47份月季品种表型多样性分析及综合评价[J].江西农业大学学报,2021,43(1):94-105. YIN S H, WANG K, HUANG X X, et al. Phenotypic diversity analysis and comprehensive evaluation of 47 rose resources[J]. Acta Agriculturae Universitatis Jiangxiensis,2021,43(1):94-105.
[8] 王禹,张广辉,赫京生,等.杜鹃花色研究进展[J].世界林业研究,2020,33(5):19-24. WANG Y, ZHANG G H, HE J S, et al. Research progress of Rhododendron flower color[J]. World Forestry Research,2020, 33(5):19-24.
[9] 沈德周,纵丹,周安佩,等.AFLP引物组合数对准确分析马缨杜鹃遗传多样性的影响[J].西南林业大学学报,2016,36(6): 22-28. SHEN D Z, ZONG D, ZHOU A P, et al. Effect of the quantity of AFLP primer combinations on accurately analyzing Rhododendron delavayi genetic diversity[J]. Journal of Southwest Forestry University,2016,36(6):22-28.
[10] 司马永康,郭玉红,徐德兵,等.云南红花油茶基于花瓣数量的分类[J].西部林业科学,2018,47(5):80-89. SIMA Y K, GUO Y H, XU D B, et al. Classification of Camellia reticulata Lindley based on its petal numbers[J]. Journal of West China Forestry Science,2018,47(5):80-89.
[11] 沈立新,梁洛辉,王庆华,等.腾冲红花油茶自然类型及其品种类群划分[J].林业资源管理,2009(6):75-80. SHEN L X, LIANG L H, WANG Q H, et al. The natural types and variety groups of Camellia reticulata. Lindley[J]. Forest Resources Management,2009(6):75-80.
[12] 吴贵进,陈龙清,耿芳,等.滇山茶登录品种统计及表型性状分析[J].园艺学报,2023,50(10):2157-2170. WU G J, CHEN L Q, GENG F, et al. Statistics and phenotypic traits analysis of Camellia reticulata registered cultivars[J]. Acta Horticulturae Sinica,2023,50(10):2157-2170.
[13] 林存学,杨晓华,刘海荣.东北寒地96份李种质资源表型性状遗传多样性分析[J].园艺学报,2020,47(10):1917-1929. LIN C X, YANG X H, LIU H R. Genetic diversity analysis of 96 plum germplasm resources by phenotypic traits in northeast cold area[J]. Acta Horticulturae Sinica,2020,47(10):1917-1929.
[14] WESSINGE C A, HILEMAN L C. Accessibility, constraint, and repetition in adaptive floral evolution[J]. Developmental Biology,2016,419(1):175-183.
[15] KHADIVI-KHUBAND A, ANJAM K. Morphological characterization of Prunus scoparia using multivariate analysis[J]. Plant Systematics and Evolution,2014,300:1361-1372.
[16] OLIVEIRA E J D, DIAS N L P, DANTAS J L L. Selection of morpho-agronomic descriptors for characterization of papaya cultivars[J]. Euphytica,2012,185(2):253-265.
[17] 唐毓玮,彭绿春,段福军,等.15个高山杜鹃品种的花表型性状多样性分析[J/OL].分子植物育种,https://link.cnki.net/urlid/ 46.1068.S.20231025.1128.004. TANG Y W, PENG L C, DUAN F J, et al. Analysis of floral phenotypic trait diversity of 15 Rhododendrons cultivars[J/OL]. Molecular Plant Breeding,https://link.cnki.net/urlid/46.1068. S.20231025.1128.004.
[18] 徐斌,彭莉霞,张方秋,等.杜鹃红山茶叶片主要性状的遗传多样性分析[J].植物研究,2015,35(5):730-734. XU B, PENG L X, ZHANG F Q, et al. Genetic diversity analysis for leaf main traits of Camellia azalea[J]. Bulletin of Botanical Research,2015,35(5):730-734.
[19] 伍思宇,梁晓静,廖健明,等.中国金花茶与越南金花茶叶表型差异性分析[J].广西林业科学,2019,48(3):336-341. WU S Y, LIANG X J, LIAO J M, et al. Analysis on leaf phenotypic differences of Chinese and Vietnamese Camellia section Chrysantha Chang[J]. Guangxi Forestry Science,2019, 48(3):336-341.
[20] 戴艳花,杜佩莲,郝丙青,等.香花油茶不同家系群体叶片和果实表型性状的比较[J].经济林研究,2023,41(4):109-117. DAI Y H, DU P L, HAO B Q, et al. Comparison of phenotypic characters of leave and fruit in different family groups of Camellia osmantha[J]. Non-wood Forest Research,2023,41(4):109-117.
[21] 李芳,霍达,罗扬,等.西南红山茶天然居群种实表型性状变异分析[J].种子,2021,40(4):109-114. LI F, HUO D, LUO Y, et al. Variation analysis of phenotypic traits of seed of natural populations of Camellia pitardii in southwest China[J]. Seed,2021,40(4):109-114.
[22] 甘沛华,沈德周,何承忠,等.不同花型滇山茶遗传多样性与遗传关系的AFLP分析[J].植物生理学报,2019,55(8):1222-1230. GAN P H, SHEN D Z, HE C Z, et al. AFLP analysis on genetic diversity and relationship of Camellia reticulata of different floral patterns[J]. Plant Physiology Journal,2019,55(8):1222-1230.
[23] 韦开萍,杨茜,和润喜,等.腾冲红花油茶花芽分化及雌配子体发育的形态学观察[J].经济林研究,2023,41(3):244-251. WEI K P, YANG X, HE R X, et al. Morphological study on floral bud differentiation and the development of female gametophytes in Camellia reticulate Lindl f. simplex Sealy[J]. Non-wood Forest Research,2023,41(3):244-251.
[ 24] 张晓庆.中国茶花品种分类、测试指南及已知品种数据库构建[D].北京:中国林业科学研究院,2008. ZHANG X Q. The classification of Chinese Camellia cultivars, test guideline and construction of database for the known cultivars[D]. Beijing: Chinese Academy of Forestry,2008.
[25] 中华人民共和国农业部.农业行业标准NY/T 2943—2016 茶树种质资源描述规范[S].北京:中国农业出版社,2016. Ministry of Agriculture of the PRC. Chinese Agricultural Standard NY/T 2943-2016 Specification for the description of tea plant germplasm resources[S]. Beijing: China Agriculture Press,2016.
[26] SUMARTINI, T. Evaluation of mungbean lines for resistance to Cercospora leaves spot and powdery mildew (Erysiphe polygoni) diseases in the green house[J]. Earth and Environmental Science,2021,743(1):1-7.
[27] 张悦,索玉静,李芳东,等.柿种质资源果实形态多样性分析[J].园艺学报,2022,49(7):1473-1490. ZHANG Y, SUO Y J, LI F D, et al. Analysis on fruit morphological diversity of persimmon germplasm resources[J]. Acta Horticulturae Sinica,2022,49(7):1473-1490.
[28] 喻华平,赵志常,高爱平,等.基于主成分分析和聚类分析的23份黄皮种质资源的品质评价[J].热带作物学报,2022,43(7): 1357-1364. YU H P, ZHAO Z C, GAO A P, et al. Quality evaluation of 23 species of Clausena lansium (Lour.) Skeels germplasm resources based on principal component analysis and cluster analysis[J]. Chinese Journal of Tropical Crops,2022,43(7):1357-1364.
[29] 郭松,李在留,陈晶晶,等.不同种源掌叶木果实和种子表型性状多样性分析及综合评价[J].植物资源与环境学报,2018, 27(4):11-20. GUO S, LI Z L, CHEN J J, et al. Diversity analysis and comprehensive evaluation on phenotypic traits of fruit and seed of Handeliodendron bodinieri from different provenances[J]. Journal of Plant Resources and Environment,2018,27(4):11-20.
[30] SHAIJU P N, OMANAKUMARI N. Floral morphology and systematics of the genus Thottea Rottb. (Aristolochiaceae) from the western Ghats, India[J]. Plant Systematics and Evolution,2012,288:213-225.
[31] ZHOU T, FAN J J, ZHAO M M, et al. Phenotypic variation of floral organs in Malus using frequency distribution functions[J]. BMC Plant Biology,2019,19(1):1-11.
[32] 向贵生,王其刚,邱显钦,等.云南川滇蔷薇天然居群表型多样性分析[J].云南大学报,2018,40(4):786-794. XIANG G S, WANG Q G, QIU X Q, et al. Phenotypic diversity of natural population of Rosa soulieana in Yunnan[J]. Journal of Yunnan University,2018,40(4):786-794.
[33] 张莹,曹玉芬,张小双,等.基于花表型性状的梨种质资源多样性研究[J].园艺学报,2016,43(7):1245-1256. ZHANG Y, CAO Y F, ZHANG X S, et al. Research on diversity of pear germplasm resources based on flowers phenotype traits[J]. Acta Horticulturae Sinica,2016,43(7):1245-1256.
[34] 颉刚刚,欧阳丽婷,谢军,等.新疆地区欧洲李叶片表型性状多样性及亲缘关系分析[J].植物资源与环境学报,2018,27(3): 72-78. XIE G G, OUYANG L T, XIE J, et al. Analyses on diversity of leaf phenotypic traits and genetic relationships of Prunus domestica in Xinjiang region[J]. Journal of Plant Resources and Environment,2018,27(3):72-78.
[35] 吴根松,孙丽丹,郝瑞杰,等.梅花种质资源表型多样性研究[J].安徽农业科学,2011,39(20):12008-12009. WU G S, SUN L D, HAO R J, et al. Study on the phenotypic diversity of P. mume Sieb. et Zucc. germplasm resources[J]. Journal of Anhui Agricultural Sciences,2011,39(20):12008-12009.
[36] 陈文星,杨琳懿,唐军荣,等. 10个新樟属植物叶片表型性状多样性分析[J].中南林业科技大学学报,2023,43(12):83-93. CHEN W X, YANG L Y, TANG J R, et al. Analysis of the diversity on leaf phenotypic traits of 10 Neocinnamomum genus[J]. Journal of Central South University of Forestry Technology,2023,43(12):83-93.
[37] 孙泽硕,李永华,柳新红,等.四十一份樱花品种资源表型性状遗传多样性分析[J].北方园艺,2022(22):57-66. SUN Z S, LI Y H, LIU X H, et al. Genetic diversity analysis of phenotypic traits in 41 cultivars of oriental cherry[J]. Northern Horticulture,2022(22):57-66.
[38] 柳江群,尹明宇,左丝雨,等.长柄扁桃天然种群表型变异[J].植物生态学报,2017,41(10):1091-1102. LIU J Q, YIN M Y, ZUO S Y, et al. Phenotypic variations in natural populations of Amygdalus pedunculata[J]. Chinese Journal of Plant Ecology,2017,41(10):1091-1102.
[39] 王辉,谢利娟,罗丹,等.深圳外来月季品种的数量分类研究[J].西南林业大学学报,2013,33(4):81-87. WANG H, XIE L J, LUO D, et al. Mathematic classification of the introduced rose cultivars in Shenzhen[J]. Journal of Southwest Forestry University,2013,33(4):81-87.
[40] 辛桐.云南山茶花Camellia reticulata多样性的人文因素及遗传背景[D].北京:中央民族大学,2015. XIN T. The cultural factors and genetic background of the diversity of Camellia reticulata[D]. Beijing: Central University for Nationalities,2015.
[41] 张海平,房伟民,陈发棣,等.部分睡莲属植物形态性状的多样性分析[J].南京农业大学学报,2009,32(4):47-52. ZHANG H P, FANG W M, CHEN F D, et al. Investigation on the morphological diversity of taxa in genus Nymphaea[J]. Journal of Nanjing Agricultural University,2009,32(4):47-52.
[42] 丁释丰,高筱钰,吴林源,等.广东黄心树种质表型与遗传多样性分析[J].南方农业学报,2021,52(5):1291-1299. DING S F, GAO X Y, WU L Y, et al. Phenotype and genetic diversity of Machilus gamblei King ex Hook. f. germplasm in Guangdong[J]. Journal of Southern Agriculture,2021,52(5):1291-1299.
[43] 苏群,杨亚涵,田敏,等.49份睡莲资源表型多样性分析及综合评价[J].西南农业学报,2019,32(11):2670-2681. SU Q, YANG Y H, TIAN M, et al. Phenotypic diversity analysis and comprehensive evaluation of 49 waterlily resources[J]. Southwest China Journal of Agricultural Sciences,2019,32(11):2670-2681.
[44] 陶萍,宋燚,张晴晖,等.云南普洱茶种古茶树资源表型性状数据分析研究[J].西部林业科学,2020,49(6):144-151. TAO P, SONG Y, ZHANG Q H, et al. Data analysis of phenotypic characteristics of ancient tea resources of Yunnan Pu’er tea species[J]. Journal of West China Forestry Science,2020, 49(6):144-151.
[45] KIM E J, SA K J, PARK K C. Study of genetic diversity and relationships among accessions of foxtail millet Setaria italica (L.) P. Beauv. in Korea, China, and Pakistan using SSR marker[J]. Genes Genomics,2012,34(5):529-539.
[46] 童海浪,张华健,李健,等.四倍体油茶全基因组SSR位点开发与特征分析[J].中南林业科技大学学报,2024,44(3):126-135. TONG H L, ZHANG H J, LI J, et al. Development and characterization of whole genome SSR in tetraploid Camellia oleifera[J]. Journal of Central South University of Forestry Technology,2024,44(3):126-135.
[47] 付超,刘凯,张幸,等.广西凤山中籽茶群体种质SNP标记开发及遗传进化分析[J].经济林研究,2023,41(3):169-175,186. FU C, LIU K, ZHANG X, et al. SNP markers development and genetic structure analysis of Camellia oleifera genetic populations germplasm in Fengshan of Guangxi[J]. Non-wood Forest Research,2023,41(3):169-175,186.
[本文编校:吴 彬]
