
氮素营养对白檀幼苗生长及光合特性的影响
2024-12-31 00:00:00曹剑锋李凤汪芝玲韦启娇冉全艳丁波
天津农业科学 2024年8期





摘" " 要:为确定适合白檀幼苗快速生长的最佳施氮量和提高白檀幼苗培育效率,以1年生白檀幼苗为试验材料,设置CK(0 g·株-1)、 N1(4.5 g·株-1)、N2(7.5 g·株-1)、N3(10.5 g·株-1)和N4(13.5 g·株-1)5个氮素水平的氮肥施用试验,研究氮素营养对白檀幼苗生长、叶片叶绿素含量、光合参数与叶绿素荧光参数的影响。结果表明:适量施用氮肥显著促进了白檀幼苗株高、地径和叶片面积的生长,上述指标均在N2处理下达到最大值,分别较CK提高67.49%、50.00%、49.95%;与CK相比,4个氮肥处理不同程度地增加了叶片叶绿素含量,提升了净光合速率(Pn),促进了水分利用率(Wue),以N2处理的叶绿素含量、净光合速率(Pn)和水分利用率(Wue)最高,分别比CK提高63.6%、89.8%、79.9%;在暗适应条件下,白檀叶片光系统Ⅱ(PSⅡ);最大光化学效率(Fv/Fm)、实际光合效率( ΦPS Ⅱ)、光化学猝灭系数(qP)和表观光合电子传递速率(ETR)均随着施氮量的增加呈先上升后下降的趋势,并且均在N2处理下达到最大值,非光化学淬灭系数(NPQ)降低。综上所述,适量施用氮肥能显著提升白檀适应强光的能力和光合能力,促进白檀幼苗的生长。本试验条件下,白檀幼苗适宜的施氮量为7.10~7.86 g·株-1。
关键词:白檀幼苗;氮营养;生物量;光合特性;叶绿素荧光
中图分类号:S793.9" " " " " " "文献标识码:A" " " " " " "DOI 编码:10.3969/j.issn.1006-6500.2024.08.001
Effects of Nitrogen Nutrition on Seedling Growth and Photosynthetic Characteristics of Symplocos tanakana
CAO Jianfeng1,2,3, LI Feng1, WANG Zhilin1, WEI Qijiao1, RAN Quanyan1, DING Bo1,3
(1.School of Biological Sciences,Guizhou Education University, Guiyang, Guizhou 550018, China; 2.Institute of Medicinal Plants Biotechnology, Guizhou Education University, Guiyang, Guizhou 550018, China; 3.Guizhou Bioresource Development and Utilization Key Laboratory, Guiyang, Guizhou 550018, China)
Abstract:To determine the optimal nitrogen fertilizer dosage for the rapid growth of Symplocos tanakana seedlings and enhance seedling cultivation efficiency, 1-year-old Symplocos tanakana seedlings were used as experimental materials to set up 5 levels of nitrogen fertilizer application experiment, CK ( 0 g·plant-1), N1 (4.5 g·plant-1), N2 (7.5 g·plant-1), N3 (10.5 g·plant-1) and N4 (13.5 g·plant-1). The effects of nitrogen nutrition on seedling growth, chlorophyll content, photosynthetic parameters and chlorophyll fluorescence parameters of S.tanakana seedlings were studied. The results showed that appropriate nitrogen fertilizer application significantly promoted plant height, ground diameter, and leaf area of S. tanakana seedlings, with the highest values observed under N2 treatment, increasing by 67.49%, 50.00%, and 49.95% compared to the control group.Chlorophyll content, net photosynthetic rate (Pn), and water utilization efficiency(Wue) also increased under 4 fertilization treatments, with peak values under N2 treatment showing increases of 63.6%, 89.8%, and 79.9% compared to the control group. Maximum photochemical efficiency(FV/FM) under PSII dark adaptation, actual photosynthetic efficiency (ΦPS II), photochemical quenching coefficient (qP), and apparent photosynthetic electron transfer rate (ETR) of S.tanakana leaves initially increased and then decreased with nitrogen application, peaking under N2 treatment. Meanwhile, non-photochemical quenching coefficient(NPQ) decreased. In conclusion, proper nitrogen fertilizer application can enhance adaptability of S.tanakana to strong light,increase photosynthetic capacity, and effectively promote seedling growth. In this test, the optimal nitrogen supply was found to be 7.10-7.86 g·plant-1.
Key words: Symplocos tanakana; nitrogen nutrient; biomass; photosynthetic; chlorophyll fluorescence
白檀( Symplocos tanakana) 为山矾科山矾属落叶灌木或小乔木,枝叶茂盛,可闻香、观花、观果,是优良的绿化资源。白檀果实(果肉和种子均含油)含油率高(36.6%)[1],白檀油中不饱和脂肪酸占比达85%以上,我国部分山区已将其作为食用油[2],也可作为性能优良的生物柴油和油脂基材料[3]。白檀木坚硬致密、纹理美观,树皮及根可药用[4],根系发达,防风固土能力强,可在喀斯特地等劣质地貌生长[5]。目前,白檀仍处于半野生状态或野生状态,尚未规模化开发利用,也无可推广的生产良种,对白檀树的人工栽培、优良资源评价和良种选育等还缺乏深入研究[6],特别是白檀肥效利用方面未见研究报道,从而限制了白檀资源的开发和推广应用。
氮是植物生长、发育和繁殖的必需元素[7-8]。在农业和园艺生产中,含氮化肥经常被大量使用,以实现粮食和果蔬的高产量[9-11]。然而,一部分氮肥并没有被植物利用,而是释放到大气、土壤和水圈中[12-13],造成土壤酸化、土壤营养失衡和下游水富营养化等严重危害[14]。因此,在林木培育实践中,合理施肥是促进苗木生长、改善苗木质量、提高成活率的关键技术之一,也是防治农业面源污染的重要措施。研究施肥对苗木光合生理代谢的影响,对调节苗木生长、提高苗木质量和抗逆性、减少生态环境破坏具有重要意义。近年来,国内外有关施肥对植物生长影响的研究较多[15-16]。合理适量施用氮素能显著提升幼苗光合能力,增加生物量,但施用过多的氮素不利于幼苗生长。因此,本试验主要研究不同氮素水平对白檀幼苗生长、光合生理和叶绿素荧光特性的影响,确定白檀生长所需的最佳氮素施用量及条件,以便进一步为白檀高效栽培的氮肥管理与调控提供理论依据。
1 材料与方法
1.1 试验材料
本试验于2023年春季在贵州黔南瓮安县白檀育苗试验苗圃(107°32′54″E,26°54′28″N)进行。试验地土壤为黄壤,pH值为5.34,有机质含量为34.8 g·kg-1,全氮、全磷、全钾含量分别为2.26、0.90、1.47 g·kg-1。白檀种植面积为0.17 hm2。选择生长健康且长势一致的一年生实生幼苗,带土移栽到苗圃试验田,株距为50 cm,行距为20 cm。缓苗8周后,进行不同的施肥处理。
1.2 试验设计
根据土壤肥力现状,采用随机区组试验设计,共设置5个处理。每个处理30株,每个处理3次重复。试验施用的氮肥为尿素CO(NH2)2(46% N),均衡肥使用“优乐卉”(以色列海法·凯尔凯特平衡型钾冠大量元素水溶肥),总氮、五氧化二磷、氧化钾含量均为20%。施肥方式采用单株施肥,施肥3次。每次施肥量(纯氮)如下:CK处理为0 g·株-1;N1、N2、N3、N4处理分别为1.5、2.5、3.5、4.5 g·株-1。所有处理均施用平衡肥五氧化二磷1.5 g·株-1、氧化钾1.5 g·株-1。5月28日、6月28日、7月28日,在距离根茎8 cm处,绕苗木挖深8 cm、宽10 cm的环状沟,将肥料与土壤混合后撒入沟内并覆土。施肥结束后,N1、N2、N3、N4处理的单株施肥总氮量分别为4.5、7.5、10.5、13.5 g。6月25日、7月25日,分别除草1次,育苗期间进行常规的水分管理。
1.3 测试指标
1.3.1 苗木生长量与生物量" 2023年5月28日,每个处理选择15株幼苗,测定地径、苗高和冠幅,之后每月测定1次。2023年9月中旬,测定生物量,每个施肥处理选取5株幼苗,测定根、茎、叶。
1.3.2 叶片叶绿素含量 叶绿素含量测定参照Pires等[17]的方法。在施肥结束后(9月中旬),每个处理随机摘取5株幼苗的新鲜叶片,提取制备叶绿素测试液,测定每克叶片的叶绿素a含量、叶绿素b含量和叶绿素总量。
1.3.3 光合参数 每个处理随机选取5株植株,每株选取2片健康叶片,采用LI-6800型便携式光合测定仪测定白檀幼苗的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci),并计算瞬时水分利用率(Wue)。测定时间为2023年9月12日—13日,每日9:00—11:30进行。
1.3.4 叶绿素荧光参数 本研究采用MINI-PAM-II型便携式荧光测定仪对叶片荧光动力学参数进行测定。选取不同处理的白檀幼苗顶端完全展开的第2片叶片,测定各叶绿素荧光参数,计算PSII最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSⅡ)、表观量子传递速率(ETR)、光化学淬灭系数(qP)和非光化学淬灭(NPQ)等。
1.4 数据分析与统计方法
本研究利用Microsoft Excel 软件整理原始数据,利用Prism软件绘图,利用SPSS 16.0 统计软件进行方差分析和相关性分析。
2 结果与分析
2.1 施氮量对白檀幼苗生长的影响
2.1.1 施氮量对白檀幼苗主要形态变化的影响 白檀幼苗的株高、地径和冠幅的增长量变化如表1所示。6月白檀幼苗生长缓慢,7月白檀幼苗生长加速,8月白檀幼苗生长进入快速生长期。施肥90 d后,株高和地径随着氮肥施用量的增加呈先增后降的趋势。N2处理的净增长量达到最大值,与CK相比,株高和地径分别增加67.49%、50.00%;施肥90 d后,各施肥处理的白檀幼苗叶片冠幅增长量均高于CK,随着施氮量的增加,各施肥处理的叶片冠幅增长差异不显著。
由图1可知,施肥90 d后,各施肥处理的叶长、叶宽和叶面积呈先增加后减小的趋势。N2处理的叶长、叶宽和叶面积最大,分别比CK增加23.37%、26.89%、49.95%。N4处理的叶片各指标显著小于N2处理。
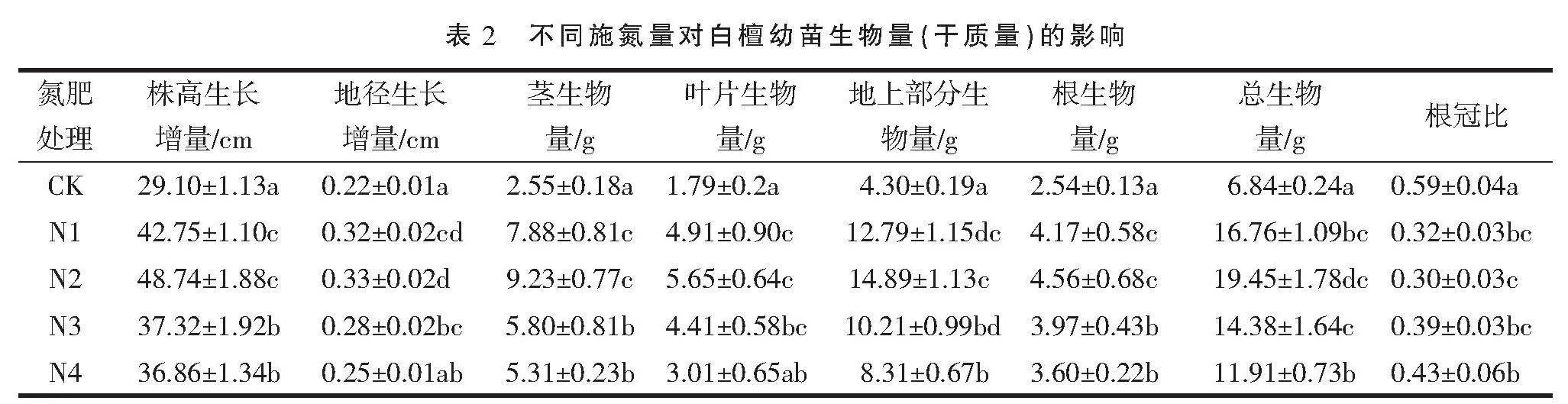
2.1.2 不同施氮量对白檀幼苗生物量增长的影响 由表2可知,不同氮肥处理的根、茎、叶生物量、总生物量均显著高于CK,并且N2处理的根生物量、茎生物量、叶生物量、总生物量均为最大值,分别为4.56、9.23、5.65、19.45 g,分别比CK增长1.80倍、3.62倍、3.16倍、2.84倍。N2处理的根、茎、叶生物量与N1处理无显著差异,与N4处理有显著性差异;但N1、N2、N3处理的总生物量之间无显著性差异。此外,不同氮肥处理的白檀幼树根冠比范围为0.30~0.43,并且各处理的根冠比均小于CK,N1、N2、N3处理之间差异未达到显著水平。
2.2 不同施氮量对白檀幼苗叶片叶绿素含量的影响
由图2可知,随着施肥量的增加,白檀幼苗叶片叶绿素a与叶绿素b含量、叶绿素总量呈先升高后降低的趋势,不同氮肥处理的上述指标含量均高于CK。其中,N1、N2、N3处理与CK有显著性差异。N2处理的叶绿素a与叶绿素b含量、叶绿素总量均达到最大值,与CK相比,增长量分别为56.97%、63.64%、58.86%。由叶绿素a/b比值可知,施肥降低了该比值,不同施肥处理的叶绿素a/b值均小于CK,不同施肥处理间差异不显著。
2.3 不同施氮量对白檀幼苗光合特性的影响
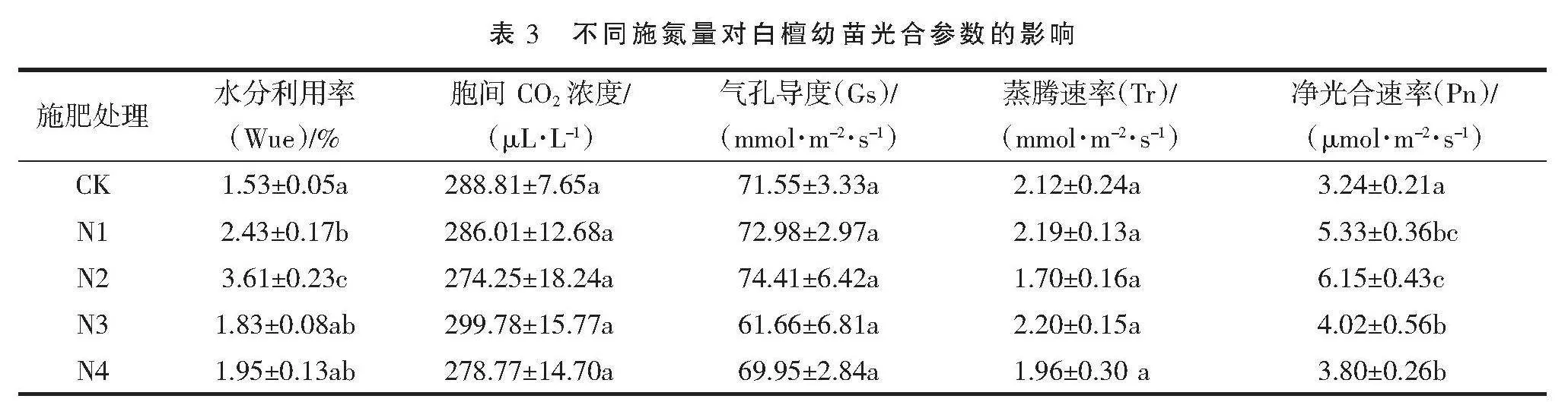
由表3可知,与CK相比,不同氮肥处理的白檀幼苗叶片水分利用率Wue均高于CK,分别增加72.8%、79.9%、8.9%、8.3%,其中N2处理的水分利用率(Wue)最大。细胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)和气孔导度(Gs)差异不明显。不同氮肥处理的净光合速率(Pn)均高于CK,分别增加64.5%、89.5%、24.1%、17.3%,其中N2处理的净光合速率(Pn)最大,N4处理的净光合速率(Pn)显著小于N2处理。上述结果表明,氮肥的施用显著提升了白檀幼苗的水分利用率(Wue)和净光合速率(Pn),并且N2处理的氮素促进效果最好。
2.4 不同施氮水平对白檀幼苗叶绿素荧光参数的影响
由图3可知,施用氮肥后,与CK相比,N1、N2、N3、N4处理的Fv/Fm分别增加3.3%、4.8%、1.3%、0.4%,其中N2处理的Fv/Fm最高。白檀幼苗叶片光合系统的ΦPSⅡ、ETR 和qP等参数均随着氮素施用量的增加与光合作用参数Pn表现出一致的变化趋势,其中N2处理的上述指标均达到最大值后下降。施肥90 d后,与CK相比,N1、N2、N3、N4处理的ΦPSⅡ分别增加22.4%、29.4%、18.2%、17.1%,ETR分别增加18.5%、52.6%、14.2%、10.2%,qP分别增加13.2%、15.1%、13.2%、9.8%。随着施氮量的增加,白檀幼苗叶片的ΦPSⅡ、qP呈明显降低趋势。NPQ值在各处理组间差异不显著(Pgt;0.05)。
2.5 施氮量与白檀幼苗生长及其光合特征参数相关性分析
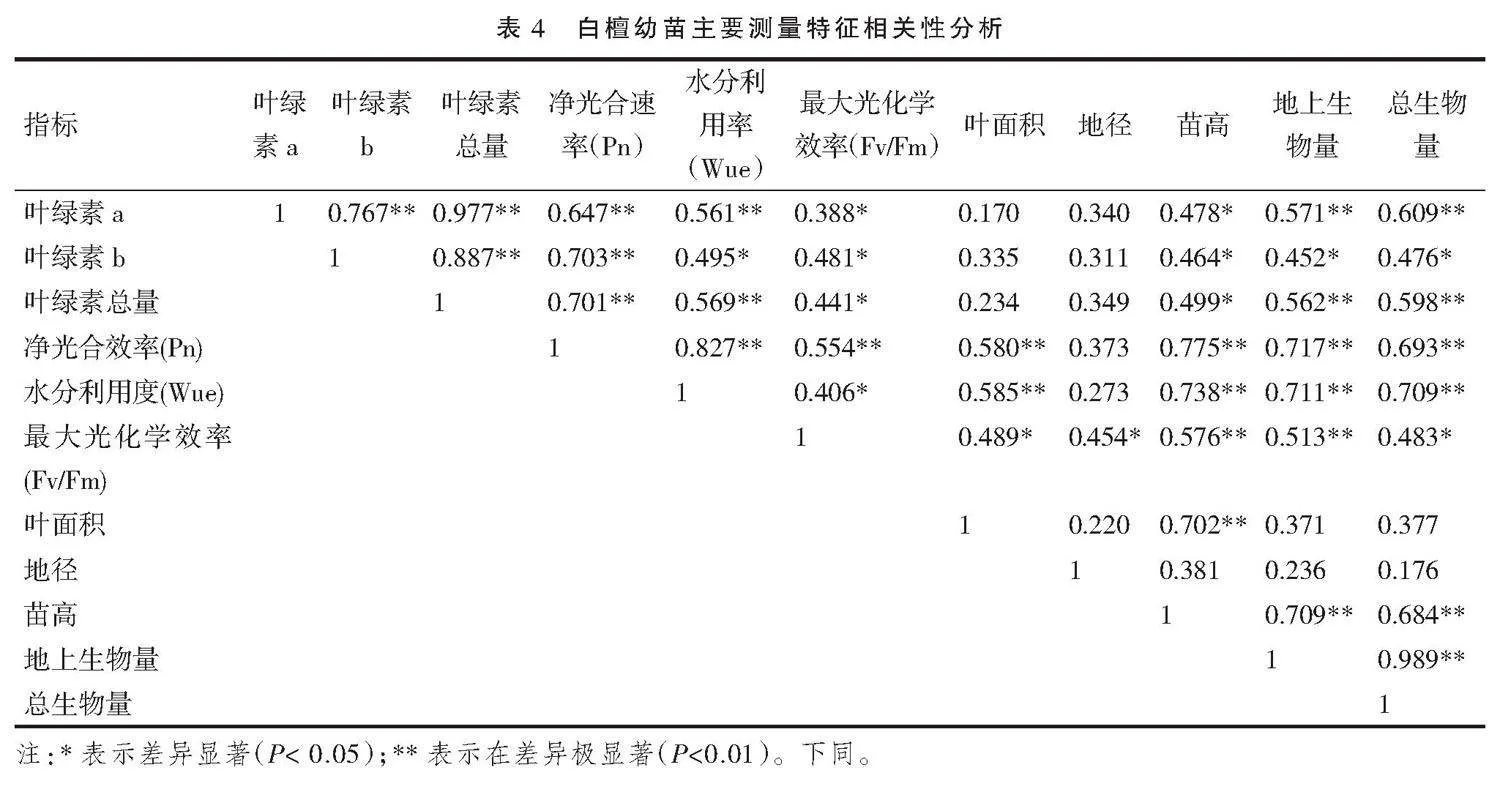
对白檀幼苗苗高、叶绿素含量、叶面积、净光合速率(Pn)等11个生理生态指标进行相关性分析,结果如表4所示。幼苗生长总生物量、地上生物量、叶片面积和苗高均与净光合速率(Pn)、水分利用率(Wue)、最大光化学效率(Fv/Fm)呈显著正相关。这说明氮肥的施用使植株的最大光化学效率(Fv/Fm)提高、净光合速率(Pn)明显提升,从而显著提升白檀适应强光的能力和光合能力,促进了光合产物的积累。从试验结果可以看出,适当施用氮肥后,幼苗的净光合速率(Pn)、水分利用率(Wue)、最大光化学效率(Fv/Fm)显著高于CK,幼苗光合叶片面积显著增大,植株苗高和地径显著增加,植株生物量显著积累。随着叶绿素a含量的增加,叶绿素b含量也正向增加,呈极显著正相关。叶绿素a和b与光合参数净光合速率(Pn)、水分利用率(Wue)、最大光化学效率(Fv/Fm)也呈显著正相关,叶绿素a和叶绿素b与幼苗地径间无显著相关性。
2.6 施氮量与白檀幼苗生长指标间的回归分析
将白檀幼苗生长指标与施氮量的关系拟合成一元二次回归方程[18],分析推算得到各指标的理论最大值和对应的适宜施氮量(表5)。白檀苗高和地径的增长量与叶面积的最大值分别为46.61、0.32、14.67 cm2,相应的适宜施氮量分别为7.22、7.10、7.67 g·株-1;根生物量、茎生物量、叶生物量、地上部分生物量、总生物量的最大值分别为4.41、8.40、5.43、13.84、18.53 g·株-1,相应的适宜施氮量分别为7.86、7.42、7.33、7.39、7.49 g·株-1;叶绿素总量的最大值为2.04 mg·g-1,适宜的施氮量为7.18 g·株-1。比较分析可知,白檀幼苗适宜的施氮量为7.10~7.86 g·株-1。
3 讨论与结论
3.1 讨论
氮素在植物生理生化活动中起着至关重要的作用,尤其在光合生理中表现尤为突出,并且植物幼苗时期对氮素的需求量最大[19]。氮素占植物叶片叶绿体组成的75%[20],不但是叶绿素的关键构成元素,也是多种蛋白质的必需组成成分[21]。氮素对作物最终产量的贡献率范围为40%~50%,是植物生长不可或缺的关键元素[22-23]。然而,植物主要通过土壤吸收氮素,而自然界土壤中氮、磷、钾含量普遍较低,这成为制约植物生长的主要因素。已有研究结果表明,适量施用氮肥可显著提高林木光合速率,从而促进林木幼苗地径和株高的增加[24-25],但过量施肥会导致林木光合能力下降,光合产物减少[26],进而抑制其生长发育[27]。
本研究深入探讨了施用氮肥对白檀幼苗生长的影响。本研究发现,氮肥处理增加了白檀幼苗叶片面积、株高、地径、地上部分生物量。这表明施用氮肥可促进白檀幼苗光合作用,进而促进幼苗地上部分生长。氮肥不仅影响植物地上部分的生长,也影响植物地下根系营养的分配,合理施用氮肥能促进植物将额外产生的碳水化合物分配给根系,以促进地下根系生长[28]。白檀幼苗根冠比的差异表明,施用氮肥后,白檀幼苗植株地上部分生物量分配多于地下部分,说明施氮更有利于地上部分生物量的累积,这与欧洲鹅耳枥[29]、桤木[30]和北美红杉[31]等林木施用氮肥的效应一致。然而,研究结果也表明,过量施用氮肥对幼苗生长会产生不利影响。值得注意的是,植物营养供给过剩造成生长抑制的生物学过程和机制仍需进一步探索。
光合生理特性的研究结果显示,氮肥处理显著提高了白檀幼苗叶片叶绿素含量,为光合作用提供了重要的光合色素基础。由净光合速率(Pn)的变化可知,不同处理的净光合速率(Pn)存在显著差异,各处理的净光合速率(Pn)平均值均高于CK,其中N2处理的净光合速率(Pn)最高。N2处理的白檀幼苗能够更加高效地利用光能,促进光合产物积累,从而促进植株生长[32-34]。这种效应可能与氮素促进叶绿素合成以及核酮糖1,5-二磷酸羧化酶(Rubisco)活性和含量的提高有关[35]。然而,当超过一定的氮肥施用量时,白檀幼苗光合速率有所降低,表现出过量施用氮肥的副作用,这与白檀幼苗生物量的变化一致。这表明过量施用氮肥限制了白檀幼苗光合能力,减少了光合产物积累,从而抑制了幼苗生长。原因可能是氮肥施用过多,导致光合含氮产物大量合成,限制了幼苗对P、Mg等矿质元素的吸收和代谢,进而影响植物光合效率[36];另外,过量施用氮肥也可能导致幼苗叶片参与碳同化RuBP羧化酶的数量和活性降低[24-25,37],使得光合作用减弱,影响幼苗的生长。还有一种观点认为,过量供应氮素会破坏叶绿体,降低光合作用效率[38]。
白檀幼苗叶片叶绿素荧光效应的测定结果显示,氮肥处理显著提高了白檀幼苗叶片的Fm、ΦPSⅡ和Fv/Fm值,并且N2处理的Fm和Fv/Fm均达到最大值,这说明适宜施用氮肥可提高白檀幼苗光能捕获、转移和转化效率。然而,N4处理的Fm和Fv/Fm反而降低,这说明过量供应氮素使PSII的活性中心受损,导致电子传递降低,抑制其PSII光化学效率,这与各氮肥处理的净光合速率(Pn)变化一致[39-40]。光化学淬灭系数(qP)在一定程度上反应了光系统II反应中心的开放程度[41]。本研究中,施用氮素后,白檀幼苗叶片qP升高,这说明氮素促进了白檀幼苗叶片PSII光反应中心开放,但较高的氮浓度使qP下降,说明过量施用氮肥会导致白檀幼苗发生光抑制,降低电子传递速率[42]。由相关性分析结果可知,白檀幼苗不同生长特征与光合因子之间存在着紧密的相关性。适宜施用氮肥能够显著提高白檀幼苗叶片的光化学效率和电子传递速率,发挥最大的光合潜力[43],促进白檀幼苗生长和生物量的增加。
3.2 结论
综上所述,适量施用氮肥能显著提升白檀幼苗光合能力,促进其生长及生物量累积。基于不同施肥处理的白檀幼苗生长指标与光合特性的回归分析可知,氮素最佳施用量为7.10~7.86 g·株-1。因此,为确保白檀幼苗健康成长,应保持合理的氮肥施用量,避免施肥不当导致养分缺乏、生长抑制、植株死亡。未来,需进一步深化白檀对不同营养的吸收利用效率及机制研究,选育出适应多样土壤类型且养分利用高效的白檀品种,这既是当前研究的重要方向,也是农业面源污染治理的关键目标。
参考文献:
[1] 管正学, 朱太平, 仇田青. 白檀种子的油脂和氨基酸的分析与利用评价[J]. 中国野生植物, 1991(2): 11-14.
[2] 刘光斌, 刘苑秋, 黄长干, 等. 白檀油的理化性质及其制备生物柴油的研究[J]. 中国粮油学报, 2011, 26(3): 64-67.
[3] 刘强, 刘倩倩, 蒋丽娟, 等. 油料植物白檀果实中油脂的积累规律[J]. 经济林研究, 2015, 33(1): 73-78.
[4] SEMWAL R B, SEMWAL D K, SEMWAL R, et al. Chemical constituents from the stem bark of Symplocos paniculata Thunb. with antimicrobial, analgesic and anti-inflammatory activities[J]. Journal of Ethnopharmacology, 2011, 135(1): 78-87.
[5] 刘倩倩, 刘强, 孙友平, 等. 白檀果实内含物与矿物质变化规律及相关性[J]. 经济林研究, 2017, 35(2): 145-150.
[6] 安佰义, 郭才南, 包文慧, 等. 白檀离体快繁技术[J]. 植物学报, 2018, 53(5): 693-699.
[7] KRAPP A. Plant nitrogen assimilation and its regulation: a complex puzzle with missing pieces[J]. Current Opinion in Plant Biology, 2015, 25: 115-122.
[8] TEGEDER M, MASCLAUX-DAUBRESSE C. Source and sink mechanisms of nitrogen transport and use[J]. New Phytologist, 2018, 217(1): 35-53.
[9] ZÖRB C, LUDEWIG U, HAWKESFORD M J. Perspective on wheat yield and quality with reduced nitrogen supply[J]. Trends in Plant Science, 2018, 23(11): 1029-1037.
[10] GALLOWAY J N, ABER J D, ERISMAN J W, et al. The nitrogen cascade[J]. BioScience, 2003, 53(4): 341-356.
[11] LIU J G, YOU L Z, AMINI M, et al. A high-resolution assessment on global nitrogen flows in cropland[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(17): 8035-8040.
[12] LUO J, ZHOU J J. Growth performance, photosynthesis, and root characteristics are associated with nitrogen use efficiency in six poplar species[J]. Environmental and Experimental Botany, 2019, 164: 40-51.
[13] PITRE F E, POLLET B, LAFARGUETTE F, et al. Effects of increased nitrogen supply on the lignification of poplar wood[J]. Journal of Agricultural and Food Chemistry, 2007, 55(25): 10306-10314.
[14] DISE N B, ROTHWELL J J, GAUCI V, et al. Predicting dissolved inorganic nitrogen leaching in European forests using two independent databases[J]. Science of the Total Environment, 2009, 407(5): 1798-1808.
[15] 耿东梅, 单立山, 李毅, 等. 土壤水分胁迫对红砂幼苗叶绿素荧光和抗氧化酶活性的影响[J]. 植物学报, 2014, 49(3): 282-291.
[16] 张卫强, 黄芳芳, 甘先华, 等. 施肥对银叶树幼苗生长及光合特性的影响[J]. 广西植物, 2021, 41(6): 862-871.
[17] PIRES M V, ALMEIDA A A F, FIGUEIREDO A L, et al. Photosynthetic characteristics of ornamental passion flowers grown under different light intensities[J]. Photosynthetica, 2011, 49(4): 593-602.
[18] 王玉杰, 张大克. 多元肥料效应函数模型的优化方法[J]. 生物数学学报, 2002, 17(1): 74-77.
[19] SCOULLAR K, SEELY B, WELHAM C, et al. Forecasting forest futures: A hybrid modelling approach to the assessment of sustainability of forest ecosystems and their values[M]. London: Routledge, 2010.
[20] CECHIN I, DE FÁTIMA FUMIS T. Effect of nitrogen supply on growth and photosynthesis of sunflower plants grown in the greenhouse[J]. Plant Science, 2004, 166(5): 1379-1385.
[21] GRASSI G, MEIR P, CROMER R, et al. Photosynthetic parameters in seedlings of Eucalyptus grandis as affected by rate of nitrogen supply[J]. Plant, Cell amp; Environment, 2002, 25(12): 1677-1688.
[22] KINDRED D R, VERHOEVEN T M O, WEIGHTMAN R M, et al. Effects of variety and fertiliser nitrogen on alcohol yield, grain yield, starch and protein content, and protein composition of winter wheat[J]. Journal of Cereal Science, 2008, 48(1): 46-57.
[23] SYLVESTER-BRADLEY R, KINDRED D R. Analysing nitrogen responses of cereals to prioritize routes to the improvement of nitrogen use efficiency[J]. Journal of Experimental Botany, 2009, 60(7): 1939-1951.
[24] MANTER D K, KAVANAGH K L, ROSE C L. Growth response of Douglas-fir seedlings to nitrogen fertilization: importance of Rubisco activation state and respiration rates[J]. Tree Physiology, 2005, 25(8): 1015-1021.
[25] NAKAJI T, FUKAMI M, DOKIYA Y, et al. Effects of high nitrogen load on growth, photosynthesis and nutrient status of Cryptomeria japonica and Pinus densiflora seedlings[J]. Trees, 2001, 15(8): 453-461.
[26] SU B Q, WANG L F, SHANGGUAN Z P. Morphological and physiological responses and plasticity in Robinia pseudoacacia to the coupling of water, nitrogen and phosphorus[J]. Journal of Plant Nutrition and Soil Science, 2021, 184(2): 271-281.
[27] 何金金, 邓甜甜, 刘倩钰, 等. 不同施肥处理对紫金牛(Ardisia japonica)生长和生理特性的影响[J]. 生态科学, 2023, 42(4): 92-97.
[28] POORTER H, NAGEL O W. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review[J]. Australian Journal of Plant Physiology, 2000, 27(6): 595-607.
[29] 钱燕萍, 祝遵凌. 氮素营养对欧洲鹅耳枥幼苗生长及光合特性的影响[J]. 西北农林科技大学学报(自然科学版), 2016, 44(6): 71-78.
[30] 饶龙兵, 李霞, 段红平, 等. 不同供氮水平对4种桤木幼苗生长的影响[J]. 林业科学研究, 2012, 25(2): 246-253.
[31] 白尚斌, 王懿祥, 左显东, 等. 北美红杉幼苗对不同供N水平的生长反应[J]. 林业科学研究, 2005, 18(5): 561-566.
[32] 尹丽, 胡庭兴, 刘永安, 等. 施氮量对麻疯树幼苗生长及叶片光合特性的影响[J]. 生态学报, 2011, 31(17): 4977-4984.
[33] 邢鸿林, 刘天义, 扎史都吉, 等. 光照与氮添加对红皮云杉幼树生长与叶形态功能的影响[J]. 森林工程, 2022, 38(4): 1-9.
[34] 王力朋, 李吉跃, 王军辉, 等. 指数施肥对楸树无性系幼苗生长和氮素吸收利用效率的影响[J]. 北京林业大学学报, 2012, 34(6): 55-62.
[35] 向芬, 李维, 刘红艳, 等. 氮素水平对不同品种茶树光合及叶绿素荧光特性的影响[J]. 西北植物学报, 2018, 38(6): 1138-1145.
[36] YANG J Y, ZHENG W, TIAN Y, et al. Effects of various mixed salt-alkaline stresses on growth, photosynthesis, and photosynthetic pigment concentrations of Medicago ruthenica seedlings[J]. Photosynthetica, 2011, 49(2): 275-284.
[37] 靳甜甜, 傅伯杰, 刘国华, 等. 不同坡位沙棘光合日变化及其主要环境因子[J]. 生态学报, 2011, 31(7): 1783-1793.
[38] 李佳秋, 田佳赫, 于丽娜, 等. 不同施氮量对皂荚幼苗生长及光合特性的影响[J]. 林业与生态科学, 2024, 39(1): 28-33.
[39] 甘龙, 罗玉红, 李晓玲, 等. Cd胁迫下一年蓬的生长、Cd积累及叶绿素荧光特性[J]. 武汉大学学报(理学版), 2018, 64(1): 70-78.
[40] ZHUANG J, WANG Y L, CHI Y G, et al. Drought stress strengthens the link between chlorophyll fluorescence parameters and photosynthetic traits[J]. PeerJ, 2020, 8: e10046.
[41] 何勇, 符庆功, 朱祝军. 低温弱光对辣椒叶片光合作用、叶绿素荧光猝灭及光能分配的影响[J]. 核农学报, 2013, 27(4): 479-486.
[42] 孔芬, 刘小勇, 王港振, 等. 施肥量对山地核桃光合与快速叶绿素荧光特性的影响[J]. 林业科学研究, 2016, 29(5): 764-769.
[43] 陈琳, 曾杰, 徐大平, 等. 氮素营养对西南桦幼苗生长及叶片养分状况的影响[J]. 林业科学, 2010, 46(5): 35-40.
猜你喜欢
今日农业(2020年19期)2020-12-14 14:16:52
山东农业科学(2017年2期)2017-03-15 18:55:09
山东农业科学(2016年12期)2017-01-21 14:41:54
山东农业科学(2016年12期)2017-01-21 14:39:43
山东农业科学(2016年11期)2016-12-17 21:16:16
吉林农业·下半月(2016年10期)2016-12-16 23:45:53
现代园艺(2016年2期)2016-03-15 16:05:02
江苏农业科学(2015年11期)2016-01-27 14:59:30
中国科技信息(2015年2期)2015-11-16 08:18:32
江苏农业科学(2015年8期)2015-09-10 16:07:34
