
苦荞杂交后代株型性状和产量性状的遗传分析
2024-12-27 00:00:00李春花加央多拉吴晗梁秀雪李玉明王艳青李晗孙墨可田娟任长忠
南方农业学报 2024年9期




摘要:【目的】对苦荞杂交后代株型性状和产量性状进行遗传分析,为矮秆苦荞新品种培育提供良好的理论依据。【方法】以株高差异较大的2个苦荞品种杂交F2和F3代群体为研究对象,分析其株高、主茎节数、一级分枝数、二级分枝数、茎粗、单株粒数、单株粒重和千粒重8个农艺性状的遗传规律,并对性状间的相关性进行分析。【结果】8个农艺性状在F2和F3群体中均出现超亲分离;主茎节数和千粒重的变异系数小于20.00%,单株粒数和单株粒重的变异系数大于50.00%,其他性状的变异系数为20.00%~40.00%。株高、主茎节数、单株粒数和单株粒重的加性方差小于显性方差,平均显性度均大于1.00;一级分枝数、二级分枝数、茎粗和千粒重的显性方差小于加性方差,平均显性度小于1.00。千粒重的狭义遗传率、广义遗传率和固定遗传率均大于其他性状。F3代群体农艺性状间相关分析结果显示,株高和单株粒重均与单株粒数、千粒重、主茎节数、一级分枝数、二级分枝数和茎粗存在极显著正相关(Plt;0.01、Plt;0.001)。筛选出13个株高比母本矮、单株粒数和单株粒重比母本多的优良单株,其中有3个单株的株高比母本矮50 cm及以上。13个优良单株的农艺性状间相关分析结果显示,株高与主茎节数存在极显著正相关(Plt;0.01),单株粒重与茎粗、单株粒数存在极显著正相关(Plt;0.001),株高和单株粒重均与其他性状无显著相关(Pgt;0.05)。【结论】苦荞矮秆高产育种过程中,由于千粒重与其他性状相关系数较小,且具有较高遗传率,可在杂交初期世代进行单株选择;株高与其他主要农艺性状存在显著或极显著正相关,则需要综合考虑性状间的相关性进行单株选择,才能达到选育高产矮秆苦荞品种的目的。
关键词:苦荞;株型;产量;遗传规律;杂交育种
中图分类号:S517.035.1文献标志码:A文章编号:2095-1191(2024)09-2558-09
Genetic analysis on plant type traits and yield traits in tartary buckwheat hybrid progeny
LI Chun-hua1,Jiayangduola1,WU Han1,LIANG Xiu-xue1,LI Yu-ming2,WANG Yan-qing3,LI Han1,SUN Mo-ke1,TIAN Juan1,REN Chang-zhong1*
(1Baicheng Academy of Agricultural Sciences,Baicheng,Jilin 137000,China;2Integrated Service Center ofTianqiaoling Town of Wangqing County,Yanbian,Jilin 133204,China;3Biotechnology and GermplasmResources Institute,Yunnan Academy of Agricultural Sciences/Yunnan Provincial Key Lab of AgriculturalBiotechnology/Key Lab of Southwestern Crop Gene Resources and Germplasm Innovation,Ministry ofAgriculture and Rural Affairs,Kunming,Yunnan 650205,China)
Abstract:【Objective】The study aimed to conduct a genetic analysis of plant type traits and yield traits in tartary buckwheat hybrid progeny,providing theoretical basis for the cultivation of new dwarf tartary buckwheat varieties.【Method】The study focused on the F2 and F3 generation populations of hybrid progeny from 2 tartary buckwheat varietieswith great differences in plant height.The genetic patterns of 8 agronomic traits were analyzed,including plant height,node number of main stem,number of first branches,number of secondary branches,stem diameter,number of seeds per plant,seed weight per plant,and thousand-kernel weight,along with the analysis of the correlation among these traits.【Result】The 8 agronomictraits exhibited superparental segregation in both F2 and F3 populations.The coefficients of variation for node number of main stem and thousand-kernel weight were less than 20.00%,while the coefficients for the number of seeds per plant and seed weight per plant were greater than 50.00%.The coefficients for all other traits ranged from 20.00%to 40.00%.The additive variance for plant height,node number of main stem,number of seeds per plant and seed weight per plant was less than the dominant variance,with a mean degree of dominance greater than 1.00.Conversely,for the number of first branches,number of second branches,stem diameter and thousand-kernel weight,the dominant variance was less than the additive variance,resulting in a mean degree of dominance less than 1.00.In this case,the narrow sense heritability,broad sense heritability and fixed heritability of thousand-kernel weight were greater than those of the other traits.Correlation analysis among agronomic traits in the F3 generation population revealed that both plant height and seed weight per plant were extremely significantly and positively correlated with number of seeds per plant,thousand-kernel weight,node number of main stem,number of first branches,number of secondary branches and stem diameter(Plt;0.01,Plt;0.001).A total of 13 superior lines were identified which were shorter than the female pa-rent and had a greater number of seeds per plant and seed weight per plant than the female parent,with 3 lines being at least 50 cm shorter than the female parent.Correlation analysis among the agronomic traits of the 13 superior lines re-vealed extremely significant positive correlation between plant height and the node number of main stem(Plt;0.01),and extremely significant positive correlation between seed weight per plant and both stem diameter and number of seeds per plant(Plt;0.001).Both plant height and seed weight per plant were not significantly correlated with the other traits(Pgt;0.05).【Conclusion】In the breeding process for high-yield dwarf tartary buckwheat,thousand-kernel weight has a low cor-relation coefficient with other traits and exhibits high heritability,allowing for single-plant selection in the early genera-tions of hybridization.However,since plant height shows significant or extremely significant positive correlation with other main agronomic traits,it is necessary to comprehensively consider the correlations among traits during single-plant selection to achieve the optimal selection target for high-yield dwarf tartary buckwheat.
Key words:tartary buckwheat;plant type;yield;genetic patterns;hybrid breeding
Foundation items:National Natural Science Foundation of China(31860412);China Agriculture Research System—Oat and Buckwheat(CARS-07);Baicheng Guiding Scientific and Technological Development Plan Project(202210)
0引言
【研究意义】苦荞(Fagopyrum tataricum Gaertn.)具有耐寒、生育期短的特性,是我国经济作物和避灾救荒作物,也是中西部生态环境脆弱且欠发达地区的主要粮食作物(陆平,1994;林汝法,2013)。苦荞具有很高的营养价值和药用价值(王迪等,2023),作为保健食品深受人们的喜爱,其生产受到人们的重视(李旭霞等,2023;余莎等,2023),市场对苦荞的需求逐年增加,但其产量低,难以满足市场需求(陈庆富,2018)。株型是影响苦荞群体产量的重要性状,主要包括株高、茎粗、分枝数、叶片数、花序数及分枝部位高度和分枝区长度等(林汝法,2013)。多年来,较多学者致力于提高荞麦生产能力研究,培育出许多高产的荞麦新品种(马宁等,2016;杨媛等,2016;张清明等,2016;王安虎等,2017),但这些品种在生产过程中易倒伏,严重影响植株农艺性状及籽粒品质,甚至减产50%以上(向达兵等,2014;钟林等,2020)。倒伏是作物减产的主要原因(伍浩天等,2020),其中植株过高是易发生倒伏的重要因素之一(Wang et al.,2015a;高鑫等,2012)。株高作为农作物的重要农艺性状,是株型的重要组成部分,与产量、倒伏性等密切相关(罗炬等,2012)。因此,探究苦荞株高的遗传特性,并进行矮秆抗倒伏亲本的创制和育种,对提高苦荞产量具有重要的意义。【前人研究进展】近年来,关于荞麦倒伏性的问题越来越受到诸多学者的关注(向达兵等,2014;Wang et al.,2015b;佘恒志等,2018;高翔等,2019;陶建波等,2023)。据水稻和小麦相关研究报道,自1960年半矮秆基因sd1导入到禾本科植物以来,选育了无数的半矮秆品种,极大地提高了粮食产量(Hedden,2003),后续矮秆性状和矮秆品种对产量的贡献已被众多科研和生产实践所证实,矮化育种已成为重要的育种手段(薛建峰等,2011;Daoura et al.,2014;Wang et al.,2014;Yang et al.,2015;董晓杰等,2022)。研究证明,株高与倒伏呈显著正相关,降低株高是提高抗倒伏性的最有效措施(张志才,2006)。因此,培育矮秆品种是解决苦荞倒伏问题最经济、最有效的途径。苦荞的株高等株型性状及产量相关性状均属于数量性状,基因型和环境共同控制其表型(林汝法,2013),鉴定数量性状基因时直接利用传统的育种方法难度很大。要有效开展苦荞矮秆育种必须深入了解苦荞株高及相关性状的遗传特性。同时,要减少育种过程中选择的盲目性,提高育种效率,需要利用与矮秆基因连锁的遗传标记,对目标性状进行跟踪选择,缩小育种群体规模(Gupta et al.,1999;董娜等,2013)。【本研究切入点】当前,我国春夏季种植的大多数苦荞品种的植株较高,易倒伏。作物株高与产量、抗倒性等紧密相关,因此对苦荞株型性状和产量性状开展遗传分析及分子生物学研究十分必要。目前,未见有关苦荞杂交后代株型性状和产量性状的遗传规律解析及矮秆品种培育的研究报道。【拟解决的关键问题】利用株高差异较大的2个苦荞品种杂交后代开展株型性状和产量性状的遗传规律及相关分析,并从其后代中筛选出矮秆高产新品系,为苦荞矮秆性状的遗传基础研究提供良好的基础。
1材料与方法
1.1试验材料
试验母本材料为云南省农业科学院生物技术与种质资源研究所荞麦课题组由曲靖地方苦荞资源经60Co-γ辐射诱变后经过系统选育而成的高产优质品种云荞1号。该品种的适应范围较广、平均产量较高(3000 kg/ha以上),平均株高在130 cm以上(李春花等,2017),在全国各荞麦种植区均有种植,但种植在春夏季时植株较高,易倒伏,很难正常发挥该品种的高产效果。父本材料是在云南发现的一个矮秆的苦荞近缘野生种(F.tartaricum ssp.Potanini Bata-lin)YZ56,该品种植株较矮(60 cm以下),相对其他品种抗倒伏强,但产量低,其籽粒带刺不易加工,在生产上还未推广应用。
1.2试验方法
试验在吉林省白城市农业科学院内的大棚和试验地(45°37′N,122°47′E)中进行,该地海拔143.65 m。2019年9月对云荞1号和YZ56进行杂交,获得了F1代杂交种。2020年4月将其杂交种自交获得了236粒F1代的种子,同年9月将236粒F1代的种子与亲本各10粒,每盆1粒种植在花盆,并在10月初将花盆移到温室中,自交获得F2代的种子,收获时对F2代群体236个单株及亲本进行农艺性状测定。2021年6月将亲本各10粒和每株系各1粒组成的F3代群体236粒进行盆栽,每盆种1粒,自交获得F3代的种子,收获时对F3代群体236个单株及亲本进行农艺性状测定。2022年的3月,再利用单粒传法(SSD)在温室里自交获得F4代的种子,同年6月底从F4代群体236个单株中各选取50粒种植在大田中,待种子成熟时从中随机选取5个单株进行农艺性状测定。选目标材料的基准是株高比母本矮,单株粒数和单株粒重比母本大的单株。
1.3测定项目
在籽粒70%~80%成熟时,对供试材料进行株高、主茎节数、一级分枝数、二级分枝数和茎粗的测定后单独收获。单株粒数、单株粒重和千粒重的测定是在籽粒风干2周以后进行。
1.4统计分析
1.4.1 F2和F3代群体及亲本农艺性状的表现值分析
采用JMP 9.0.2和Excel 2010进行遗传多样性分析、相关分析等试验数据统计。
1.4.2农艺性状的遗传规律测定利用后代群体单株间的方差测定各农艺性状的遗传规律和遗传率(李春花等,2020)。F2和F3代群体的方差由加性方差(A)、显性方差(D)和环境方差(E1)表示。E1由公式(1)计算得出。
E1=(S1+S2)/(n1+n2-2)(1)
式中,S1和S2分别表示云荞1号和米荞的性状平方和,n1和n2分别表示亲本的个体数。
F2代群体单株间方差:Vp(F2)=1/2A+1/4D+E1(2)
F3代群体单株间方差:Vp(F3)=3/4A+3/16D+E1(3)
利用上述二元方程式(2)和(3)得出A和D,再根据李春花等(2020)的计算方式计算出广义遗传率、狭义遗传率、固定遗传率和平均显性度。
2结果与分析
2.1亲本及F2和F3代群体农艺性状测定结果
由表1可知,F2代群体的株高、主茎节数、一级分枝数、二级分枝数和千粒重平均值小于F3代群体,但F2代群体的茎粗、单株粒数和单株粒重平均值大于F3代群体。F3代群体的株高、茎粗、单株粒数、千粒重、单株粒重和F2代群体的8个性状的平均值均小于母本;F2代群体的株高、主茎节数、二级分枝数、茎粗、千粒重和F3代群体的株高、主茎节数、茎粗、单株粒数、千粒重、单株粒重的平均值均大于父本的平均值。
由表1还可知,除F2代群体株高和二级分枝数的变异系数大于F3代群体以外,其他性状的变异系数均小于F3代群体;F2和F3代群体主茎节数和千粒重的变异系数均小于20.00%,其中千粒重的变异系数最小,单株粒数和单株粒重的变异系数大于50.00%,其中单株粒重的变异系数最大,其他性状的变异系数为20.00%~40.00%。
2.2农艺性状的遗传规律分析结果
由表2可知,一级分枝数、二级分枝数、茎粗和千粒重的显性方差小于加性方差,平均显性度小于1.00;株高、主茎节数、单株粒数和单株粒重的加性方差小于显性方差,平均显性度大于1.00。8个农艺性状在F2代群体中的广义遗传率为0.62~0.86,在F3代群体中的广义遗传率为0.64~0.89,其中株高在F2和F3代群体中的广义遗传率较小,分别为0.62和0.64;千粒重在F2和F3代群体中的广义遗传率较大,分别为0.86和0.89。8个农艺性状在F2代群体中的狭义遗传率为0.41~0.65,在F3代群体中的狭义遗传率为0.51~0.77,其中主茎节数在F2和F3代群体中的狭义遗传率较小,分别为0.41和0.51;千粒重在F2和F3代群体中的狭义遗传率较大,分别为0.65和0.77。8个农艺性状的固定遗传率来为0.64~0.85,其中株高的固定遗传率最小(0.64),千粒重的固定遗传率最大(0.85)。
2.3 F3代群体农艺性状间的相关分析结果
对F3代群体农艺性状进行相关分析,结果如表3所示。除千粒重与一级分枝数和二级分枝数无显著(rgt;0.05,下同)相关性外,千粒重与其他性状间及其他性状之间均呈显著(rlt;0.05)或极显著(rlt;0.01、rlt;0.001)相关性。除千粒重与二级分枝数呈负相关外,其他性状之间均呈正相关。单株粒数与单株粒重的相关系数最大(0.983),一级分枝数与千粒重的相关系数最小(0.084)。千粒重与其他性状(除株高外)间的相关系数均小于其他性状间的相关系数,单株粒数和单株粒重与株高、主茎节数、二级分枝数、茎粗的相关系数均大于0.500以上。
2.4矮秆高产品种的筛选结果
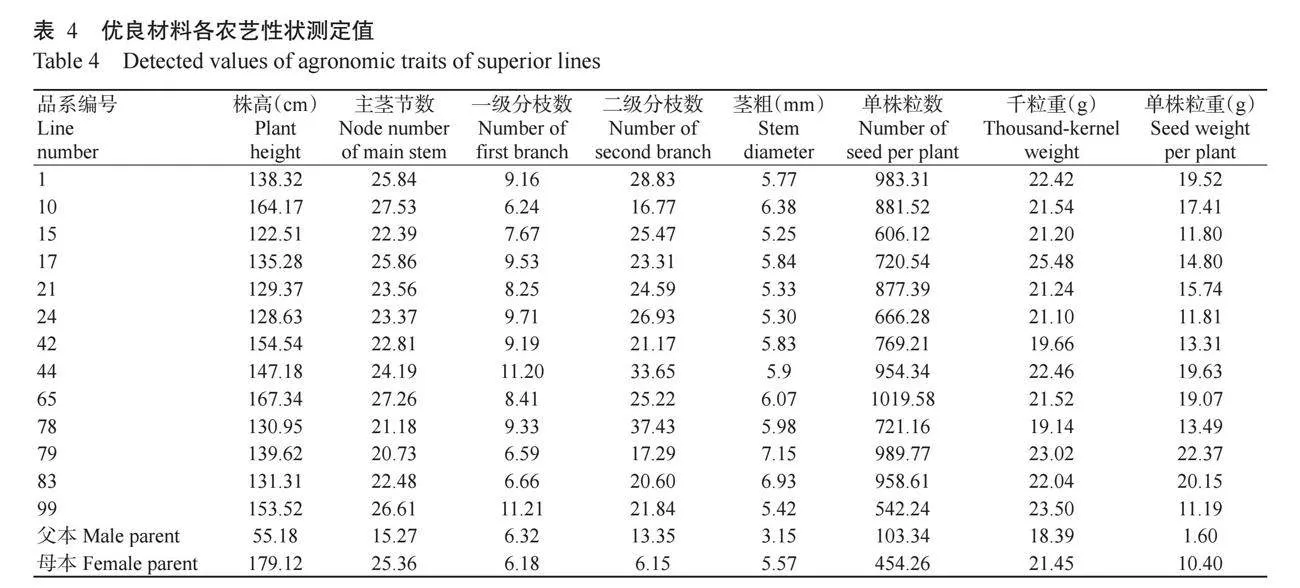
从236个单株中筛选出13个株高比母本矮,且单株粒数和单株粒重比母本多的单株(表4)。其中编号15、21、24的株高分别为122.51、129.37和128.63 cm,比母本(179.12 cm)矮49.00 cm及以上,主茎节数分别为22.39、23.56和23.37个,小于母本(25.36个),一级分枝数分别为7.67、8.25和9.71个,大于母本(6.18个),二级分枝数分别为25.47、24.59和26.93个,是母本(6.15个)的4倍以上,茎粗分别为5.25、5.33和5.30 mm,均小于母本(5.57 mm),单株粒数分别为606.12、877.39和666.28粒,比母本(454.26粒)多,单株粒重分别为11.80、15.74和11.81 g,比母本(10.40 g)重,千粒重分别为21.20、21.24和21.10 g,略小于母本(21.45 g),表明这3个单株有望可作为选育矮秆高产品种的株系。
2.5优良新材料农艺性状间的相关分析结果
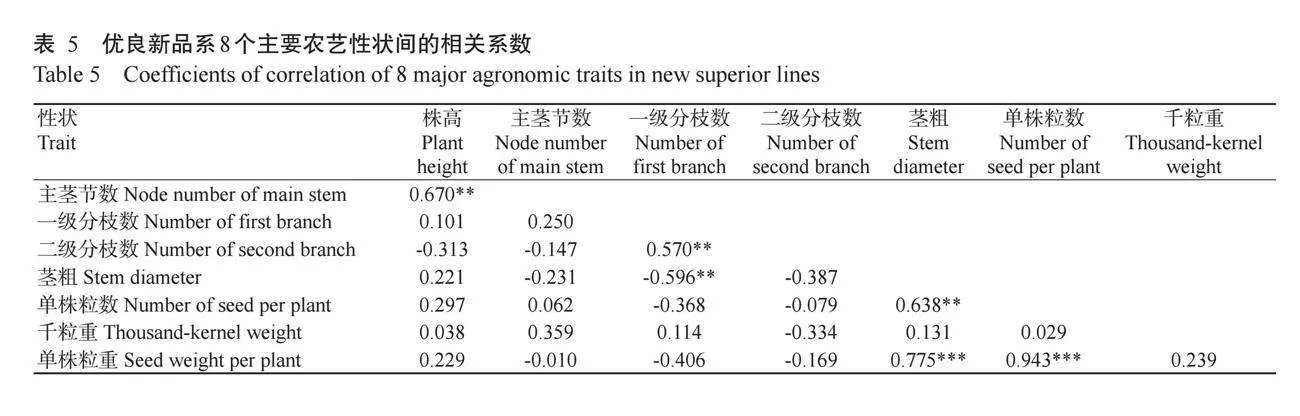
对13份优良新材料的8个农艺性状进行相关分析,结果如表5所示。株高与主茎节数、一级分枝数与二级分枝数、茎粗与单株粒数均呈极显著正相关(rlt;0.01)。单株粒重与茎粗和单株粒数呈极显著正相关(rlt;0.001)。一级分枝数和茎粗呈显著负相关(rlt;0.01)。其他大部分性状间无显著相关性。单株粒数和单株粒重的相关系数最高(0.943),主茎节数和单株粒重的相关系数最低(-0.010)。
3讨论
矮化育种中并非植株越矮越好,而是以降低株高为主要目标的株型综合改良育种(王坤波和刘正德,1996)。苦荞农艺性状的遗传规律及相关分析对矮秆育种非常关键。本研究通过对苦荞杂交后代的株型性状和产量性状进行遗传规律及相关分析,为今后矮秆苦荞育种提供理论依据。本研究结果显示,F2和F3代群体的8个农艺性状均出现超亲分离,表明杂交后代群体中出现了与父母本不同的基因型,暗示有望选择出超双亲性状的矮秆高产新品种。在不同作物的研究中,杂交后代的超亲分离是一个普通现象(梁慧珍等,2014;徐海等,2015;朱昊华等,2022)。本研究在苦荞上也验证了这一结论。作物的遗传多样性主要通过农艺性状的变异系数来体现,变异系数越大,遗传背景越丰富(Alizadeh etal.,2015)。贾瑞玲等(2015)对50份苦荞种质资源的6个主要农艺性状进行遗传多样性分析,结果发现变异系数最大的性状是单株粒重,最小的性状是千粒重。田爽琪等(2023)对3个苦荞杂交组合后代群体7个农艺性状进行遗传多样性分析,结果发现变异系数最大的性状是单株粒重(66.15%),最小的性状是千粒重(6.59%)。本研究结果也显示,苦荞主要农艺性状中,变异系数最大的性状是单株粒重,最小的性状是千粒重,与贾瑞玲等(2015)、田爽琪等(2023)的研究结果一致,说明8个农艺性状中单株粒重的个体差异较大,遗传多样性丰富,具有最大的选择和提高潜力,千粒重则选择范围窄,相对稳定。
本研究结果显示,株高、主茎节数、单株粒数和单株粒重的加性方差小于显性方差,平均显性度大于1.00,与Li等(2012)、李春花等(2020)的研究结果一致,表明株高、主茎节数和单株粒重易受外界环境因素的影响,不易在初期世代进行选拔。本研究还发现,一级分枝数的加性方差大于显性方差,并平均显性度小于1.00,与李春花等(2020)的研究结果一致,表明苦荞的一级分枝数以加性效应为主。千粒重的加性方差大于显性方差,平均显性度小于1.00,广义遗传率、狭义遗传率和固定遗传率均大于其他性状,与Li等(2019)的研究结果一致,表明千粒重可在初期世代进行单株选择。梁龙兵等(2016)对苦荞品种小米荞和晋荞2号杂交后代F2和F3代群体的7个农艺性状进行分析,结果显示单株粒数和株高的广义遗传率和狭义遗传率均最大。田爽琪等(2023)对3个苦荞杂交组合后代群体的7个农艺性状进行分析,结果发现单株粒重的广义遗传率最大,单株粒数的狭义遗传率最大。这与本研究发现苦荞株高的广义遗传率最小,千粒重的广义遗传率最大,主茎节数的狭义遗传率最小、千粒重的狭义遗传率最大存在一定差异,其原因可能是种植环境、杂交组配、后代群体大小、分析方法等不同所导致。
已有较多作物农艺性状与产量相关分析的研究报道(Zeng et al.,2019;李炜等,2022)。唐链等(2016)对苦荞品种小米荞和晋荞2号杂交F2和F3代群体的农艺性状进行相关分析,结果发现株高与主茎分枝数、单株粒重、单株粒数均存在极显著正相关。田爽琪等(2023)对3个苦荞杂交组合后代群体的7个农艺性状进行分析,结果发现株高、主茎分枝、茎基直径、单株粒数和单株粒重5个性状存在极显著正相关。本研究结果表明,株型性状(株高、主茎节数、茎粗等)与单株粒数和单株粒重间均存在极显著正相关,与唐链等(2016)、田爽琪等(2023)的研究结果一致,表明苦荞符合植株越高、茎秆越粗壮、分枝越多、单株粒数越多,产量就越高的一般规律。但由于株高越高,越易倒伏,故选择矮秆高产品种难度较大。但刘愈之等(2023)以平凉43号为母本、天水市农业科学研究所的优异抗锈材料41-42为父本进行杂交,选育了矮秆多抗高产冬小麦新品种陇麦671;周彦忠等(2017)以平凉43号为母本、天水市农业科学研究所的优异抗锈材料41-42为父本进行杂交,选育了矮秆高产花色新品种漯花4087。本研究筛选出13个株高比母本矮、单株粒数和单株粒重比母本多的优良单株,且株高除了与主茎节数存在显著正相关外,与其他性状无显著相关性,表明通过杂交可选育矮秆高产的理想型苦荞新品种,为实现苦荞株高的显著矮化,满足高产宜机化新品种选育提供了思路和借鉴。
4结论
苦荞矮秆高产育种过程中,由于千粒重与其他性状相关系数较小,且具有较高遗传率,可在杂交初期世代进行单株选择;株高与其他主要农艺性状存在显著或极显著正相关,则需要综合考虑性状间的相关性进行单株选择,才能达到选育高产矮秆苦荞品种的目的。
参考文献(References):
陈庆富.2018.荞麦生产状况及新类型栽培荞麦育种研究的最新进展[J].贵州师范大学学报(自然科学版),36(3):1-7.[Chen Q F.2018.The status of buckwheat production and recent progresses of breeding on new type of culti-vated buckwheat[J].Journal of Guizhou Normal Univer-sity(Natural Sciences),36(3):1-7.]doi:10.16614/j.gznuj.zrb.2018.03.001.
董娜,张新,王清连,李成奇,刘阳阳.2013.短季棉早熟及相关性状的QTL定位[J].核农学报,27(10):1431-1440.[Dong N,Zhang X,Wang Q L,Li C Q,Liu Y Y.2013.QTL location study on earliness and its related traits of short season cotton[J].Journal of Nuclear Agricultural Sciences,27(10):1431-1440.]
董晓杰,李志江,马金丰,李祥羽,孙广全,郑雅潞.2022.谷子矮秆育种现状及其与赤霉素敏感性关系[J].黑龙江农业科学,(3):80-86.[Dong X J,Li Z J,Ma J F,Li X Y,Sun G Q,Zheng Y L.2022.Current situation of millet dwarf breeding and its relationship with gibberellin sensitivity[J].Heilongjiang Agricultural Sciences,(3):80-86.]doi:10.11942/j.issn 1002-276.2022.03.008.
高翔,郝志萍,吕慧卿,刘璋,周忠宇,贺文文,阎昊,王官.2019.荞麦抗倒性研究进展[J].中国农学通报,35(13):6-11.[Gao X,Hao Z P,LüH Q,Liu Z,Zhou Z Y,He W W,Yan H,Wang G.2019.Research progress on lodging resistance of buckwheat[J].Chinese Agricultural Science Bulletin,35(13):6-11.]
高鑫,高聚林,于晓芳,王志刚,孙继颖,苏治军,胡树平,叶君,王海燕,崔超,李维敏.2012.高密植对不同类型玉米品种茎秆抗倒特性及产量的影响[J].玉米科学,20(4):69-73.[Gao X,Gao J L,Yu X F,Wang Z G,Sun J Y,Su Z J,Hu S P,Ye J,Wang H Y,Cui C,Li W M.2012.Stalks lodging-resistance characteristics and yield traits among different maize varieties under high close planting[J].Jour-nal of Maize Sciences,20(4):69-73.]doi:10.13597/j.cnki.maize.science.2012.04.014.
贾瑞玲,马宁,魏立平,刘彦明,南铭.2015.50份苦荞种质资源农艺性状的遗传多样性分析[J].干旱地区农业研究,33(5):11-16.[Jia R L,Ma N,Wei L P,Liu Y M,Nan M.2015.Genetic diversity analysis on the agronomic charac-teristics of 50 tartary buckwheat germplasms[J].Agricul-tural Research in the Arid Areas,33(5):11-16.]doi:10.7606/j.issn.1000-7601.2015.05.03.
李春花,孙道旺,何成兴,王艳青,卢文洁,尹桂芳,王莉花.2017.荞麦秸秆粉还田对杂草及苦荞产量的影响[J].杂草学报,35(2):61-66.[Li C H,Sun D W,He C X,Wang Y Q,Lu W J,Yin G F,Wang L H.2017.Effect of buckwheat-straw powder returning to field on weeds and tartary yield[J].Journal of Weed Science,35(2):61-66.]doi:10.19588/j.issn.1003-935X.2017.02.008.
李春花,尹桂芳,黄金亮,王艳青,卢文洁,孙道旺,王春龙,郭来春,魏黎明,加央多拉,任长忠,王莉花.2020.苦荞株型相关性状的遗传分析及其对产量的影响[J].江西农业大学学报,42(5):881-887.[Li C H,Yin G F,Huang J L,Wang Y Q,Lu W J,Sun D W,Wang C L,Guo L C,Wei L M,Jiayangduola,Ren C Z,Wang L H.2020.Genetic analysis of plant-type related traits and its effect on yield per plant in tartary buckwheat[J].Acta Agriculturae Uni-versitatis Jiangxiensis,42(5):881-887.]doi:10.13836/j.jjau.2020099.
李炜,毕影东,刘建新,王玲,刘淼,邸树峰,樊超,杨光,谢婷婷,来永才.2022.寒地野生大豆资源农艺性状的相关性和主成分分析[J].土壤与作物,11(1):10-17.[Li W,BiY D,Liu J X,Wang L,Liu M,Di S F,Fan C,Yang G,Xie T T,Lai Y C.2022.Correlation and principal component analysis for agronomic traits of wild soybean in cold region[J].Soils and Crops,11(1):10-17.]doi:10.11689/j.ieen.2095.2022.01.002.
李旭霞,师静,刘晶.2023.宁夏地区荞麦及制品加工现状调研分析[J].农产品加工,(9):84-86.[Li X X,Shi J,Liu J.2023.Investigation and analysis on the processing status of buckwheat and its products in Ningxia[J].Farm Pro-ducts Processing,(9):84-86.]doi:10.16693/j.cnki.1671-9646(X).2023.05.019.
梁慧珍,余永亮,杨红旗,张海洋,董薇,崔暐文,巩鹏涛,方宣钧.2014.幼苗期大豆根系性状的遗传分析与QTL检测[J].中国农业科学,47(9):1681-1691.[Liang H Z,Yu Y L,Yang H Q,Zhang H Y,Dong W,Cui W W,Gong P T,Fang X J.2014.Genetic and QTL analysis of root traits atseedling stage in soybean[Glycine max(L.)Merr.][J].Scientia Agricultura Sinica,47(9):1681-1691.]doi:10.3864/j.issn.0578-1752.2014.09.003.
梁龙兵,陈其皎,石桃雄,梁成刚,张启迪,唐链,饶庆琳,陈庆富.2016.苦荞杂交后代主花序特征遗传变异研究[J].河南农业科学,45(5):13-17.[Liang L B,Chen Q J,Shi T X,Liang C G,Zhang Q D,Tang L,Rao Q L,Chen Q F.2016.Genetic analysis of main inflorescence traits of hybridization progeny of tartary buckwheat[J].Journal of Henan Agricultural Sciences,45(5):13-17.]doi:10.15933/j.cnki.1004-3268.2016.05.003.
林如法.2013.苦荞举要[M].北京:中国农业科学技术出版社.[Lin R F.2013.Tartary buckwheat[M].Beijing:China Agricultural Science and Technology Press.]
刘愈之,郑琪,任根深,丁志远,刘众,王亚翠,李娜,郭怀亮.2023.矮秆多抗高产冬小麦新品种—陇麦671[J].麦类作物学报,43(6):808.[Liu Y Z,Zheng Q,Ren G S,Ding Z Y,Liu Z,Wang Y C,Li N,Guo H L.2023.Dwarf straw multi-resistant high yielding winter wheat variety-Longmai 671[J].Journal of Triticeae Crops,43(6):808.]doi:10.7606/j.issn.1009-1041.2023.06.16.
陆平.1994.荞麦低产的生理原因探析[J].种子,(1):46-47.[Lu P.1994.Exploration of physiological causes of low yield of buckwheat[J].Seed,(1):46-47.]doi:10.16590/j.cnki.1001-4705.1994.01.044.
罗炬,邵高能,魏祥进,陈明亮,唐绍清,焦桂爱,谢黎虹,胡培松.2012.一个控制水稻株高QTLqPH3的遗传分析[J].中国水稻科学,26(4):417-422.[Luo J,Shao G N,Wei X J,Chen M L,Tang S Q,Jiao G A,Xie L H,Hu P S.2012.Genetic analysis of a QTLqPH3 for plant height in rice[J].Chinese Journal of Rice Science,26(4):417-422.]doi:10.3969/j.issn.1001-7216.2012.04.005.
马宁,刘彦明,魏立平,赵小琴,贾瑞玲.2016.荞麦新品种定苦荞1号选育报告[J].甘肃农业科技,(9):1-4.[Ma N,Liu Y M,Wei L P,Zhao X Q,Jia R L.2016.Report on new-bred tartary buckwheat cultivar Dingkuqiao 1[J].Gansu Agricultural Science and Technology,(9):1-4.]doi:10.3969/j.issn.1001-1463.2016.09.001.
佘恒志,聂蛟,李英双,张玉珂,黄科慧,张园莉,方小梅,阮仁武,易泽林.2018.施硅量对甜荞倒伏及产量的影响[J].中国农业科学,51(14):2664-2674.[She H Z,Nie J,Li Y S,Zhang Y K,Huang K H,Zhang Y L,Fang X M,Ruan R W,Yi Z L.2018.Effects of silicon application rate on com-mon buckwheat lodging and yield[J].Scientia Agricultura Sinica,51(14):2664-2674.]doi:10.3864/j.issn.0578-1752.2018.14.004.
唐链,梁成刚,梁龙兵,张启迪,饶庆琳,陈庆富.2016.苦荞株高及主茎分枝数的遗传相关分析[J].江苏农业科学,44(9):129-132.[Tang L,Liang C G,Liang L B,Zhang Q D,Rao Q L,Chen Q F.2016.Genetic correlation analysis of plant height and branch number of main stem in tartary buckwheat[J].Jiangsu Agricultural Sciences,44(9):129-132.]doi:10.15889/j.issn.1002-1302.2016.09.037.
陶建波,伍浩天,王艺钢,张瑞丰,雷蕾,方小梅,易泽林.2023.硅肥和纳米土墒材料配施对苦荞倒伏的影响[J].西南大学学报(自然科学版),45(9):25-35.[Tao J B,Wu H T,Wang Y G,Zhang R F,Lei L,Fang X M,Yi Z L.2023.Effect of silica fertilizer and nano soil moisture mate-rial application on tartary buckwheat lodging[J].Journal of Southwest University(Natural Science Edition),45(9):25-35.]doi:10.13718/j.cnki.xdzk.2023.09.003.
田爽琪,唐链,王为旋,张帆,柯瑾,李洪有,石桃雄,陈庆富.2023.三个苦荞杂交组合后代群体主要农艺性状的遗传分析[J].分子植物育种.(2023-04-24)[2023-11-08].https://kns.cnki.net/kcms/detail/46.1068.S.20230424.1123.004.html.[Tian S Q,Tang L,Wang W X,Zhang F,Ke J,Li H Y,Shi T X,Chen Q F.2023.Genetic analysis on main agronomic traits of progeny populations of three tartary buckwheat(Fagopyrum tataricum)crosses[J].Melecular Plant Breeding.(2023-04-24)[2023-11-08].https://kns.cnki.net/kcms/detail/46.1068.S.20230424.1123.004.html.]
王安虎,夏明忠,蔡光泽,戴红燕.2017.苦荞新品种西荞4号的选育与栽培技术研究[J].种子,36(2):113-115.[Wang AH,Xia M Z,Cai G Z,Dai H Y.2017.Breeding and culti-vation techniques of buckwheat new varieties Xiqiao No.4[J].Seed,36(2):113-115.]doi:10.16590/j.cnki.1001-4705.2017.02.113.
王迪,杨汉梅,李阳倩,贾梦婷,邹亮,杨帆.2023.苦荞麦“品、质、效、用”的多维评价及其活性成分高值化利用的研究进展[J].浙江农业学报,35(8):1960-1974.[Wang D,Yang H M,Li Y Q,Jia M T,Zou L,Yang F.2023.Multidi-mensional evaluation of“variety,quality,efficiency and application”of tartary buckwheat and research progress of high-value utilization of active ingredients[J].Acta Agri-culturae Zhejiangensis,35(8):1960-1974.]doi:10.3969/j.issn.1004-1524.20230387.
王坤波,刘正德.1996.试论棉花矮化育种[J].中国棉花,23(9):2-3.[Wang K B,Liu Z D.1996.An experimentalstudy of cotton dwarf breeding[J].China Cotton,2(9):2-3.]
伍浩天,聂蛟,杨文娟,张智勇,吴康红,李晓瑜,方小梅,阮仁武,易泽林.2020.机播深度、播种量和施肥量对苦荞倒伏及产量的影响[J].草业学报,29(12):61-72.[Wu H T,Nie J,Yang W J,Zhang Z Y,Wu K H,Li X Y,Fang X M,Ruan R W,Yi Z L.2020.Effects of machine sowing depth and amounts of seeds and fertilizer on lodging and yield of tartary buckwheat[J].Acta Prataculturae Sinica,29(12):61-72.]doi:10.1168/cyxb2020048.
向达兵,李静,范昱,彭镰心,宋超,赵刚,赵江林.2014.种植密度对苦荞麦抗倒伏特性及产量的影响[J].中国农学通报,30(6):242-247.[Xiang D B,Li J,Fan Y,Peng L X,Song C,Zhao G,Zhao J L.2014.The effects of planting density on lodging resistance and yield of tartary buck-wheat[J].Chinese Agricultural Science Bulletin,30(6):242-247.]
徐海,宫彦龙,夏原野,闫志强,王华杰,唐亮,徐正进.2015.中日水稻品种杂交后代株型性状的变化及其相互关系[J].中国水稻科学,29(4):363-372.[Xu H,Gong Y L,Xia YY,Yan Z Q,Wang H J,Tang L,Xu Z J.2015.Varia-tions in plant type traits and their relationship of progeny derived from the cross between Chinese rice variety andJapanese rice variety[J].Chinese Journal Rice Science,29(4):363-372.]doi:10.3969/j.issn.1001-7216.2015.04.005.
薛建峰,汤永荣,查仁明,罗洪发.2011.2个水稻矮杆突变体的遗传分析[J].中国农学通报,27(27):60-64.[Xue J F,Tang Y R,Zha R M,Luo H F.2011.Genetic analysis of two dwarf mutants of rice[J].Chinese Agricultural Scien-ce Bulletin,27(27):60-64.]
杨媛,王慧,石金波,郭忠贤,李占成,李荫藩,杨芳.2016.国审苦荞新品种晋荞麦(苦)6号的选育[J].山西农业科学,44(12):1758-1760.[Yang Y,Wang H,Shi J B,Guo Z X,Li Z C,Li Y F,Yang F.2016.Breeding of a new state approved variety Jinqiaomai 6 of buckwheat[J].Journal of Shanxi Agricultural Sciences,44(12):1758-1760.]doi:10.3969/j.issn.1002-2481.2016.12.04.
余莎,甘国超,陈雪婷,蔡利,易华平,杨清,熊婷,唐雅琪.2023.凉山苦荞麦的营养价值及苦荞凉面的开发前景研究[J].现代食品,29(3):55-58.[Yu S,Gan G C,Chen X T,Cai L,Yi H P,Yang Q,Xiong T,Tang Y Q.2023.Study on the nutritional value of bitter buckwheat in Liangshan and the development prospect of bitter buckwheat cold noodles[J].Modern Food,29(3):55-58.]doi:10.16736/j.cnki.cn41-1434/ts.2023.03.012.
张清明,赵卫敏,马裕群,伍琼.2016.荞麦新品种六苦荞4号的选育及栽培管理技术[J].贵州农业科学,44(7):9-10.[Zhang Q M,Zhao W M,Ma Y Q,Wu Q.2016.Breeding of a new buckwheat cultivar Liukuqiao 4 and its cultiva-tion and management technology[J].Guizhou Agricultural Sciences,44(7):9-10.]doi:10.3969/j.issn.1001-3601.2016.07.004.
张志才.2006.作物倒伏成因分析及抗倒对策研究进展[J].耕作与栽培,(4):1-2.[Yang Z C.2006.Progress in analy-sing the causes of crop failures and countermeasuresagainst them[J].Tillage and Cultivation,(4):1-2.]doi:10.3969/j.issn.1008-2239.2006.04.001.
钟林,熊芳秋,罗晓玲,刘钢,杨馨,孙崇兰.2020.春苦荞抗倒伏栽培试验[J].农业科技通讯,(4):136-141.[Zhong L,Xiong F Q,Luo X L,Liu G,Yang X,Sun C L.2020.Experiment on lodging-resistant cultivation of spring tar-tary buckwheat[J].Bulletin of Agricultural Science and Technology,(4):136-141.]doi:10.3969/j.issn.1000-6400.2020.04.045.
周彦忠,李飞,姬小玲,卢邦林李斯佳,徐保华.2017.矮秆、高产花生新品种漯花4087的选育[J].山西农业科学,45(7):1069-1071.[Zhou Y Z,Li F,Ji X L,Lu B L,Li S J,Xu B H.2017.Breeding of new dwarf peanut variety Luo-hua 4087 with high and stable yield[J].Journal of Shanxi Agricultural Sciences,45(7):1069-1071.]doi:10.3969/j.issn.1002-2481.2017.07.06.
朱昊华,范超,王俊仁,许如根,郭宝健,朱娟,吕超.2022.不同生态类型大麦品种(系)杂交后代主要农艺性状的分析[J].大麦与谷类科学,39(1):1-7.[Zhu H H,Fan C,Wang J R,Xu R G,Guo B J,Zhu J,LüC.2022.Analysis of the main agronomic characters of hybrid progenies of barley varieties(lines)under different ecological types[J].Barley and Cereal Sciences,39(1):1-7.]doi:10.14069/j.cnki.32-1769/s.2022.01.001.
Alizadeh K,Fatholahi S,da Silva T J A.2015.Variation in the fruit characteristics of local pear(Pyrus spp.)in the northwest of Iran[J].Genetic Resources and Crop Evolu-tion,62(5):635-641.doi:10.1007/s 10722-015-0241-7.
Daoura B G,Chen L,Du YY,Hu Y G.2014.Genetic effects of dwarfing gene Rht-5 on agronomic traits in common wheat(Triticum aestivum L.)and QTL analysis on its linkedtraits[J].Field Crops Research,156:22-29.doi:10.1016/j.fcr.2013.10.007.
Gupta P K,Varsheny R K,Sharma P C,Ramesh B.1999.Molecular markers and their applications in wheat breeding[J].Plant Breeding,118(5):369-390.doi:10.1046/j.1439-0523.1999.00401.x.
Hedden P.2003.The genes of the green revolution[J].Trendsin Genetics,19(1):5-9.doi:10.1016/S0168-9525(02)00009-4.
Li C H,Kobayashi K,Yoshida Y,Ohsawa R.2012.Genetic analyses of agronomic traits in tartary buckwheat(Fagopy-rum tartaricum(L.)Gaertn.)[J].Breeding Science,62(4):303-309.doi:10.1270/jsbbs.62.303.
Li C H,Xie Z M,Wang Y Q,Lu W J,Yin G F,Sun D W,Ren C Z,Wang L H.2019.Correlation and genetic analysis ofseed shell thickness and yield factors in tartary buckwheat(Fagopyrum tataricum(L.)Gaertn.)[J].Breeding Sci-ence,69(3):464-470.doi:10.1270/jsbbs.18081.
Wang C,Hu D,Liu X B,She H Z,Ruan R W,Yang H,Yi Z L,Wu D Q.2015a.Effects of uniconazole on the lignin metabolism and lodging resistance of culm in common buckwheat(Fagopyrum esculentum M.)[J].Field Crops Research,180:46-53.doi:10.1016/j.fcr.2015.05.009.
Wang Y S,Chen L,Du Y Y,Yang Z Y,Condon A G,Hu Y G.2014.Genetic effect of dwarfing gene Rht13 compared with Rht-D1b on plant height and some agronomic traits in common wheat(Triticum aestivum L.)[J].Field Crops Research,162:39-47.doi:10.1016/j.fcr.2014.03.014.
Wang Y S,Du Y Y,Yang Z Y,Chen L,Condon A G,Hu Y G.2015b.Comparing the effects of GA-responsive dwarfing genes Rht13 and Rht8 on plant height and some agronomic traits in common wheat[J].Field Crops Research,179:35-43.doi:10.1016/j.fcr.2015.04.010.
Yang Z Y,Zheng J C,Liu C Y,Wang Y S,Condon A G,Chen Y F,Hu Y G.2015.Effects of the GA-responsive dwarfing gene Rht18 from tetraploid wheat on agronomic traits of common wheat[J].Field Crops Research,183:92-101.doi:10.1016/j.fcr.2015.07.028.
Zeng W J,Xu L,He Q L,Zhang L L,Xu H L,Liang Z S.2019.Correlation analysis of main agronomic traits and contents of diterpence lactones of Andrographis paniculata[J].China Journal of Chinese Materia Medica,44(15):3233-3238.doi:10.19540/j.cnki.cjcmm.20190527.109.
(责任编辑 陈燕)
猜你喜欢
作物学报(2022年7期)2022-05-11 12:58:02
河北果树(2022年1期)2022-02-16 00:41:00
青年歌声(2018年5期)2018-10-29 03:18:40
森林工程(2018年4期)2018-08-04 03:23:20
中国农业信息(2017年12期)2017-09-03 04:33:01
上海农业学报(2017年4期)2017-04-10 12:40:28
养生保健指南(2016年12期)2017-01-06 22:20:23
广东饲料(2016年3期)2016-12-01 03:43:12
长江蔬菜(2016年10期)2016-12-01 03:05:29
林业与生态(2016年2期)2016-02-27 14:24:01
