
基于酵母双杂交技术对与纳尔逊海湾正呼肠孤病毒σNS 相互作用宿主蛋白的筛选
2024-12-23 00:00:00孙绿茵马竹萍李润林李永刚陶晓莉
吉林大学学报(医学版) 2024年5期
[摘 要] 目的:探讨小鼠成纤维L929细胞中与纳尔逊海湾正呼肠孤病毒 (NBV) σNS蛋白相互作用 (互作) 的宿主蛋白,阐明宿主蛋白对病毒复制的影响。方法:构建表达 σNS蛋白的诱饵质粒pGBKT7-S3, 采用测序技术验证诱饵质粒在Y2H 酵母细胞中的准确表达。将pGBKT7-S3 和pGADT7 质粒分别和共同转化至Y2HGold 酵母细胞中,涂布于固体培养基进行培养,观察菌落生长情况,确认σNS 蛋白对酵母细胞无毒性,不能自行激活报告基因。诱饵质粒pGBKT7-S3 与小鼠成纤维L929 细胞的cDNA 文库进行杂交,对编码互作蛋白的阳性克隆进行质粒抽提,阳性测序结果通过Uniprot 数据库检索筛选得到与NBV σNS 互作的蛋白。对互作蛋白进行基因本体论(GO) 功能富集分析、京都基因和基因组百科全书(KEGG)信号通路富集分析和STRING生物信息学分析。结果:成功筛选出61 个阳性克隆。DNA 测序分析和BLAST 比对分析去除23 个未匹配到数据库和测序基本相同的阳性克隆。阳性测序结果通过Uniprot 数据库检索筛选得到38 个与NBV σNS 互作的蛋白, 31 个蛋白参与细胞生物过程,36 个蛋白是细胞解剖成分,31 个蛋白具有结合功能,5 个蛋白是线粒体呼吸链组成部分,7 个蛋白是核糖体蛋白和核糖体大小亚基的组成部分,2 个蛋白参与铁代谢稳态。GO 功能富集分析,互作蛋白参与的生物过程(BP) 富集在细胞过程、代谢过程、生物学调节、定位和对刺激的反应等;细胞组分主要是细胞解剖成分和含蛋白复合物;分子功能集中在结合、催化活性、结构分子活性和转运活性等方面。KEGG 信号通路富集分析,蛋白在遗传信息处理途径的翻译、折叠、分类和降解信号通路中高度富集,在机体系统中主要与消化系统有关联;与多种病毒性传染病和癌症有关联。STRING 分析,互作蛋白涉及核糖体蛋白类、蛋白修饰类、代谢类和免疫蛋白类等功能蛋白。结论:成功筛选出与NBV σNS蛋白互作的蛋白,宿主蛋白在病毒复制中具有重要作用。
[关键词] 纳尔逊海湾正呼肠孤病毒; 酵母双杂交实验; σNS 蛋白; 蛋白互相作用; 生物信息学
[中图分类号] R373 [文献标志码] A
呼肠孤病毒是最大的双链RNA (doublestrandedRNA,dsRNA) 病毒,可以感染很多不同的生物,包括真菌、植物、昆虫和哺乳动物,甚至人类[1]。根据诱导感染细胞之间的融合能力,正呼肠孤病毒属可以进一步划分为融合原性和非融合原性。纳尔逊海湾正呼肠孤病毒(Nelson Bayorthoreovirus,NBV) 是一种从脊髓灰质炎翼狐心脏血液中发现的融合性呼肠孤病毒[2]。近年来,NBV 已经越过物种障碍,演变成一种新兴的跨种传播频繁的人畜共患传染病病毒,可引起人类急性呼吸系统疾病[3],研究NBV 复制机制具有重要意义。NBV 直径为60~80 nm[4-5],其dsRNA 基因组由10 个大小不一的片段组成, 其中4 个小片段为S1、S2、S3 和S4[1,6]。NBV 非结构蛋白σNS 由S3基因编码[7]。σNS 是相对分子质量约为40 000 的RNA 结合蛋白,氨基酸序列高度保守,也是病毒包涵体形成和基因组复制的必需蛋白[8-12]。作为胞内寄生的病原体,宿主蛋白对NBV 的复制至关重要[13]。研究[14-15] 显示:NBV 可以感染其自然宿主蝙蝠、人类、小鼠和猴子的各种类型细胞。其中小鼠成纤维L929 细胞对NBV 感染的耐受程度较低,原因是NBV 和L929 细胞之间独有的病毒-宿主相互作用能够高效抑制病毒感染及复制并限制合胞体的形成。目前,病毒-宿主相互作用(互作) 的分子机制尚不清楚。酵母双杂交系统鉴定方法已经发展了30 余年, 但仍然是研究和鉴定蛋白质互作最直接且最具性价比的方法之一[16]。
本研究通过酵母双杂交技术结合质谱分析筛选出与NBV σNS 互作的蛋白,探讨互作蛋白的生物学功能,预测其参与的信号通路,为阐明宿主蛋白影响NBV 复制的分子机制和σNS 的致病机制提供依据。
1 材料与方法
1. 1 细胞、主要试剂和仪器 NBV-MB病毒株和成纤维L929 细胞由锦州医科大学病原生物实验室惠赠。凝胶回收试剂盒和限制性内切核酸酶(NdeⅠ和BamH Ⅰ) 购自加拿大MBI Fermentas 公司,cDNA 文库构建试剂盒、T4 DNA 连接酶和TaqDNA 聚合酶购自美国Clontech 公司, Pfu DNA 聚合酶购自香港生命科技有限公司,质粒提取试剂盒购自美国Axygen 公司,核糖核酸内切酶 H(Ribonuclease H,RNase H) 购自宝生物工程(大连) 有限公司,DNA 回收试剂盒采购自天根生化科技(北京) 有限公司,其他化学试剂均为国产分析纯。电热恒温水浴锅和精密生化培养箱购自上海精宏实验设备有限公司,超低温冰箱购自青岛海尔特种电器有限公司,倒置显微镜购自上海精密仪器仪表有限公司, PCR 仪购自北京东林昌盛科技有限责任公司,离心机购自湖南湘仪实验室仪器开发有限公司。
1. 2 小鼠 L929 细胞 cDNA 文库的构建和质量检测 采用TRIzol 法提取小鼠RNA,采用磁珠分离法纯化mRNA。以分离出的mRNA 为模板, 通过长距离PCR (long-distance PCR,LD-PCR) 反应,其产物采用分子筛原理纯化并进行均一化处理,得到cDNA。RNA、mRNA 和cDNA 均经琼脂多糖凝胶电泳检测其质量。随后,通过Infusion 酶同源重组的方式,将ds cDNA 片段分别与线性化的pGADT7-Smal-1、pGADT7-Smal-2 和pGADT7-Smal-3 载体连接, 50 ℃ 恒温培养1 h, 采用电转方式转化至DH10B 感受态细胞,37 ℃、225~250 r·min-1 摇床上培养至少1 h, 构建cDNA 文库。采用该文库菌液涂布于Luria-Bertani (LB) 培养基(含氨苄青霉素) 中培养,统计菌落数。计算小鼠L929 细胞cDNA 文库的滴度:滴度(CFU·mL-1) =培养基中的克隆数/涂布板体积(mL) ×稀释系数。随机挑取20 株单克隆,提取质粒后经PCR 扩增,检测插入片段长度, 计算cDNA 文库阳性率。cDNA文库阳性率=PCR 扩增为阳性的单克隆株数/20×100%。
1. 3 pGBKT7-S3诱饵质粒的构建 以NBV S3基因为模板, 采用Pfu DNA 聚合酶进行PCR 扩增。随后,对扩增产物进行检测、切胶和胶回收处理。采用限制性内切核酸酶EcoRⅠ和BamHⅠ分别对目的基因片段和pGBKT7 载体进行双酶切, 再采用T4 DNA 连接酶进行连接。将获得的重组质粒通过42 ℃热激法转化至TOP10 感受态细胞中。涂布于LB 固体培养基上, 37 ℃ 恒温孵育箱中倒置培养14~16 h。挑取单菌落进行PCR 验证,并对阳性克隆进行测序验证。
1. 4 诱 饵 质 粒 毒 性 检 测 和 自 激 活 检 测 将pGBKT7-S3 质粒转化至Y2HGold 酵母细胞中,同时将pGBKT7-S3 质粒和pGADT7 质粒共转化至Y2HGold 酵母细胞中。诱饵单质粒转化的菌液涂布于酵母菌色氨酸合成缺陷型培养基(syntheticdropout medium-tryptophan, SD/-Trp), 共转化的菌液涂布于选择性氨基酸缺陷型培养基(doubledropout plate, DDO)、添加3'AT 抑制剂的3 种氨基酸缺陷型培养基(triple dropout plate/3'AT,TDO/3'AT) 和4 种氨基酸缺陷型培养基(quadruple dropout plate,QDO)。30 ℃恒温培养4 d,观察固体培养基中菌落生长情况,以判断诱饵蛋白σNS 是否具有自行激活特性和酵母细胞毒性作用。
1. 5 转化 文 库 质 粒 和 阳 性 克 隆 的 筛 选 挑取SD/-Trp 固体培养基上的单个菌落,接种至SD/-Trp液体培养基中,30 ℃ 恒温225 r·min-1 摇床中培养18 h。随后转接至50 mL 酵母蛋白胨葡萄糖腺嘌呤培养基(yeast peptone dextrose adenine medium,YPDA) 中,使初始600 nm 波长处的吸光度(A) 值A (600) 达到2, 30 ℃ 恒温225 r·min-1 摇床中培养12~14 h,直至A (600) 达到0. 6~0. 8。菌液室温4 000 r·min-1离心5 min,弃上清液。采用30 mL无菌水重悬菌体,充分混匀,菌液室温4 000 r·min-1离心5 min, 弃掉上清液。采用1. 5 mL TE/LiAC混合液(0. 1 mol·L-1 TE/LiAc) 充分混匀菌体沉淀,菌液室温4 000 r·min-1离心5 min,弃上清。采用600 μL TE/LiAc 混合液(0. 1 mol·L-1 TE/LiAc)再次重悬菌体,获得诱饵菌株感受态细胞。感受态细胞与cDNA 文库质粒混合, 同时加入鲑鱼精DNA 和乙酸锂/聚乙二醇混合液(LiAc/PEG) 剧烈振荡1 min,直至完全混匀,30 ℃水浴锅中孵育45 min, 然后加入160 μL 二甲基亚砜(dimethylsulfoxide, DMSO), 42 ℃ 水浴锅中热激30 min。菌液再次离心, 弃上清。加入3 mL YPDA,30 ℃ 、225 r·min-1 培养90 min, 再次将菌液室温4 000 r·min-1离心1 min,弃上清。采用0. 9% NaCl重悬菌体,轻轻混匀,cDNA 文库质粒转化至诱饵菌株。将菌液涂布于TDO/X/3'AT 固体培养基上,观察菌落生长情况。若诱饵蛋白和文库蛋白在酵母核内互作,可激活报告基因的转录和表达,出现阳性菌落。挑取TDO/X/3'AT 固体培养基上的单个菌落,转接至QDO/X 固体培养基上,30 ℃恒温培养4~5 d,进行进一步筛选。
1. 6 猎物质粒的提取和测序比对 挑取 QDO/X固体培养基上的单个菌落,分别接种至TDO 液体培养基中,振荡培养12 h,采用酵母小量抽提试剂盒提取猎物质粒。提取的猎物质粒作为模板,利用PCR 反应进行扩增。将PCR 结果与相同体积的1×上样缓冲液充分混匀,沸水浴10 min,进行琼脂多糖凝胶电泳检测,鉴定文库的插入片段和阳性率。若扩增产物出现阳性条带,则对该样品进行通用引物 测 序,并 对 测 序 结 果 进 行 BLAST 比 对 分析。PCR 扩增和测序引物序列:pGADT7 正向引物TAATACGACTCACTATAGGGCGAGCGCCGCCATG,pGADT7 反向引物GTGAACTTGCGGGGTTTTTCAGTATCTACGATT。
1. 7 猎物蛋白的生物信息学分析 对与诱饵蛋白互作的阳性测序结果进行质谱检测。采用Uniprot(https://www. uniprot. org/) 中对应的目标物种为检索参考数据库, 进行基因本体论(GeneOntology,GO)(http://www. geneontology. org)功能富集分析和京都基因和基因组百科全书(KyotoEncyclopedia of Genes and Genomes, KEGG)(http://www. genome. ad. jp/kegg/) 信号通路富集分析。统计学分析与功能注释采用自编脚本perl(v5. 32. 1) 完 成,功 能 注 释 绘 图 采 用 R version4. 0. 2 版(2020-06-22) 和相关ggplot2 模块。以Plt;0. 05 为差异有统计学意义。采用STRING 数据库(https://string-db. org) 为检索参考数据库,识别互作蛋白,分析互作蛋白间的相互作用。采用Cytoscape 软件(V. 3. 8. 1) 进行采用蛋白-蛋白互作网络(protein-protein interaction networks, PPI)可视化。
2 结 果
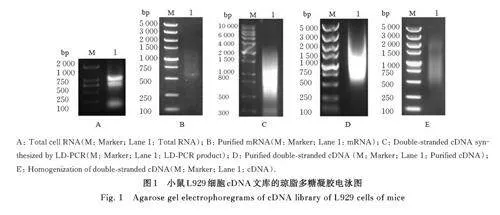
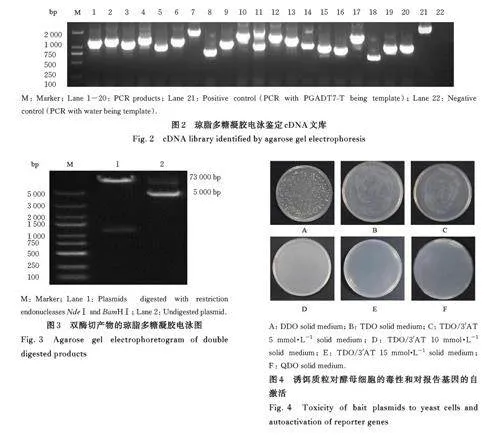
2. 1 小鼠L929细胞cDNA文库的质量检测 琼脂多糖凝胶电泳检测结果显示:cDNA 文库相对分子质量分布正常,带型分布正确,浓度和纯度数据较好,表明该文库可以用于后续实验,见图1。cDNA文库滴度为1. 86×107 CFU·mL-1。经过PCR 法鉴定,80% 的文库插入片段长度超过750 bp,文库阳性率超过90%,说明已成功构建了高质量的cDNA文库,见图2。
2. 2 构建诱饵质粒PGBKT7-S3 以NBV S3为模板, 通过PCR 扩增获得的目的片段序列大小约为1 113 bp,与预期片段大小一致。产物经纯化、双酶切和连接,获得诱饵质粒PGBKT7-S3。诱饵质粒经限制性内切核酸酶NdeⅠ和BamHⅠ双酶切鉴定,获得大小分别约为7 300 bp 和1 113 bp 的线性片段, 电泳结果与预期相符。由于展开的形状不同,超螺旋环状质粒相较于线性质粒受到的阻力较小,见图3。测序结果与已知序列一致,表明目的基因已成功插入 pGBKT7 载体。
2. 3 诱饵质粒 PGBKT7-S3的毒性和自激活 将PGBKT7 质粒和PGBKT7-S3 诱饵质粒共同转化至Y2H Gold 酵母细胞中,涂布于DDO 固体培养基上培养,观察到菌落生长,表明诱饵质粒成功转入酵母细胞且对酵母细胞无毒性, 见图4A。共转染PGBKT7-S3 和PGBKT7 的Y2H Gold 感受态在TDO 固体培养基上生长,表明诱饵质粒激活了酵母细胞报告基因HIS3 的表达, 见图4B。TDO/3'AT 5 mmol·L-1 固体培养基上观察到菌落生长,而在 TDO/3'AT 10 mmol·L-1 和15 mmol·L-1 固体培养基上未见菌落生长, 表明10 mmol·L-1 3'AT即可使诱饵质粒不能自行激活报告基因HIS3的表达,见图4C~4E。因此可以添加10 mmol·L-1 3'AT 进行下一步筛库实验。在QDO 固体培养基上未见菌落生长,表明诱饵质粒不能自行激活酵母细胞报告基因ADE2 的表达,见图4F。
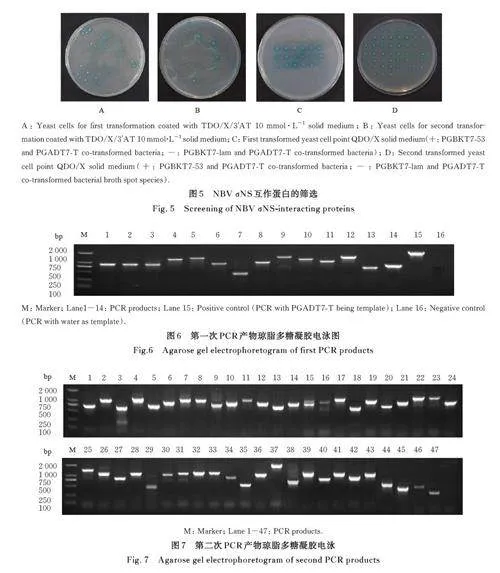
2. 4 NBV σNS 互作蛋白的筛选 将诱饵质粒pGBKT7-S3 与小鼠L929 细胞cDNA 文库质粒共转至感受态细胞中,涂布于TDO/X/3'AT 10 mmo·l L-1固体培养基上进行筛选培养,见图5A 和5B。挑选阳性克隆,点种至新的QDO/X 固体培养基上,筛选与NBV σNS 互作的候选蛋白,见图5C 和5D。
2. 5 猎物质粒提取和测序分析 提取阳性克隆的质粒,采用通用引物对提取的质粒进行PCR 扩增,并对扩增产物进行琼脂多糖凝胶电泳检测,结果显示:文库插入片段长度各不相同,猎物质粒所携带的基因片段大小不一,为500~2 000 bp,见图6 和7。
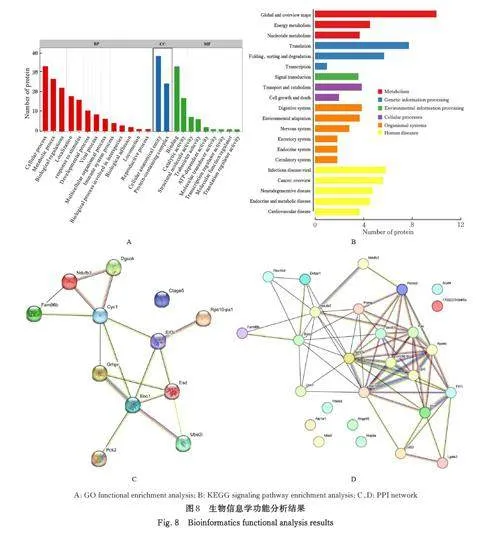
2. 6 生物信息学分析 阳性测序结果通过Uniprot数据库检索筛选得到38 个NBV σNS 互作蛋白。31 个蛋白参与细胞生物过程,36 个蛋白是细胞解剖成分,31 个蛋白具有结合功能,5 个蛋白是线粒体呼吸链组成部分,7 个蛋白是核糖体蛋白和核糖体大小亚基的组成部分, 2 个蛋白参与铁代谢稳态。对这38 个互作蛋白进行GO 功能富集分析、KEGG信号通路富集分析和STRING 生物信息学分析。GO 功能富集分析结果显示:互作蛋白参与的生物过程(biological process, BP) 富集在细胞过程、代谢过程、生物学调节、定位和对刺激的反应等;细胞组分(cellular component, CC) 主要是细胞解剖成分和含蛋白复合物; 分子功能(molecularfunction,MF) 集中在结合、催化活性、结构分子活性和转运活性等方面(图8A)。KEGG 信号通路富集分析结果显示:互作蛋白在遗传信息处理途径的翻译、折叠、分类和降解信号通路中高度富集,在机体系统中主要与消化系统相关联;与多种病毒性传染病和癌症存在关联, 见图8B。STRING 分析构建了互作蛋白之间的PPI 网络见图8C 和8D。互作蛋白涉及核糖体蛋白类、蛋白修饰类、代谢类和免疫蛋白类等功能蛋白。
3 讨 论
本课题组构建了pGBKT7-S3 质粒并通过测序验证了其准确性,将质粒转化至酵母细胞中,通过固体培养基进行筛选和检测。本研究结果显示:诱饵质粒pGBKT7-S3 对酵母细胞无毒性,且不能自行激活报告基因HIS3 和ADE2。此外, 本课题组成功构建了小鼠成纤维L929 细胞cDNA 文库。采用10 mmol·L-1 TDO /X/3’AT 和QDO/X 固体培养基进行筛选, 并进行菌落PCR 检测, 提取阳性克隆质粒, 最终得到61 个与诱饵蛋白σNS 互作,且不能自行激活报告基因HIS3 和ADE2 的cDNA文库质粒。将筛选出的文库质粒进行测序、比对和分析,最终筛选出38 个互作蛋白,采用GO 功能富集分析、BLAST 比对分析和KEGG 信号通路富集分析对上述蛋白注释、整理和分类,采用STRING数据库对PPI 挖掘,建立PPI 网络。
NHP2 样蛋白1 (NHP2-like protein 1,Snu13)、真核翻译起始因子3 亚基I (eukaryotictranslation initiation factor 3 subunit I, Eif3i)、核糖体蛋白L3 (ribosomal protein L3, Rpl3)、核糖体蛋白L19 (ribosomal protein L19,Rpl19)、核糖体蛋白L10a (ribosomal protein L10a,Rpl10a)、核糖体蛋白S4X (ribosomal protein S4X, Rps4x) 和核糖体蛋白S10 (ribosomal protein S10, Rps10) 均是核糖体蛋白,与rRNA 结合组成核糖体的大小亚基,是细胞中维持蛋白质合成过程的关键。上述蛋白与 NBV σNS 蛋白互作,可能影响 NBV 蛋白合成,继而影响病毒复制。Rpl19 定位于细胞质大核糖体亚基上,主要参与靶向膜的信号肽识别粒子信号识别颗粒(signal recognition particle, SRP) 依赖性共翻译蛋白生物过程,PPI 网络图显示其可与Rpl10a、Rpl3、Rps4x 和Snu13 互作, 共同影响核糖体功能。Rpl19 是肿瘤的生物标志物[17],也在黄热病病毒、西尼罗河病毒和寨卡病毒等多种RNA病毒复制过程中起关键作用[18], 可能促进σNS 蛋白的合成; Eif3i 参与翻译起始前复合物的形成过程,与酶结合发挥生物学功能,可与Rps10 相互作用。Eif3 复合体可以影响病毒生长,与病毒蛋白作用还可影响宿主基因的表达, 如Eif3i 是水泡性口炎病毒M 蛋白的分子伴侣, 影响病毒复制和转录[19];狂犬病病毒M 蛋白可以通过与Eif3h 互作来抑制真核翻译的启动[20], 与σNS 蛋白互作所产生的影响尚无相关研究。
细胞色素c1 (cytochrome C, Cyc1)、NADH脱氢酶[ 泛醌] 1α 亚复合物亚基7 (NADHdehydrogenase [ubiquinone] 1 α subcomplexsubunit 7, Ndufa7)、NADH 脱氢酶[泛醌] 1β 亚复合物亚基3 (NADH dehydrogenase[ubiquinone] 1 β subcomplex subunit 3, Ndufb3)、NADH 脱氢酶[ 泛醌] 黄素蛋白3 (NADHdehydrogenase [ubiquinone] flavoprotein 3,Ndufv3) 和脱氧鸟苷激酶(deoxyguanosine kinase,Dguok) 是线粒体呼吸链的关键组成部分,共同参与线粒体电子传递过程,对细胞能量代谢产生明显影响,与心血管病、神经退行性疾病和内分泌代谢病等有关联。上述蛋白之间互作可调控线粒体功能,从而影响疾病的发生发展,其中Ndufb3 在复发性流产发展过程中具有重要意义[21] 。GUARNIERI 等[22] 在啮齿动物和人类宿主中发现:严重急性呼吸系统综合征冠状病毒2 蛋白可与宿主线粒体蛋白结合,导致线粒体功能紊乱,包括下调氧化磷酸化和上调糖酵解。σNS 蛋白与线粒体蛋白结合,可能导致线粒体功能失调。
铁蛋白轻链1 (ferritin light chain 1, Ftl1) 和铁蛋白重链1 (ferritin heavy chain 1,Fth1) 在铁代谢稳态中起关键作用,主要定位于自溶酶体,协同参与细胞内铁离子隔离和运输过程,与细胞生长和死亡有密切关联。Fth1 基因启动子的低甲基化程度是一种潜在的新型冠状病毒感染诊断和预后指标[23]; 人副流感病毒2 型V 蛋白干扰Fth1 和核受体共激活剂4 (nuclear receptor coactivator 4,NCOA4) 之间的互作, 抑制NCOA4 介导的铁蛋白降解,破坏铁稳态[24],σNS 与Fth1 结合可能对铁稳态调节产生影响,具体机制有待进一步研究。
综上所述,本实验采用酵母双杂交系统结合高通量测序技术筛选出与NBV σNS 互作的蛋白,采用生物信息学工具对互作蛋白进行分析,深入了解猎物蛋白的功能、定位、活动及参与的信号通路,为进一步探讨NBV 复制过程中宿主和病毒蛋白互作的后续研究提供依据,为寻找NBV 感染治疗的新靶点提供思路。
利益冲突声明:所有作者声明不存在利益冲突。
作者贡献声明:孙绿茵、李永刚和陶晓莉参与实验设计、实验数据分析、结果讨论和论文审校,孙绿茵、马竹萍和李润林参与文献检索、实验数据收集及分析和论文撰写。
[参考文献]
[1] DAY J M. The diversity of the orthoreoviruses:
molecular taxonomy and phylogentic divides[J]. Infect
Genet Evol, 2009, 9(4): 390-400.
[2] GARD G, COMPANS R W. Structure and cytopathic
effects of Nelson Bay virus[J]. J Virol, 1970, 6(1):
100-106.
[3] KUANG G P, XU Z Q, WANG J, et al. Nelson bay
reovirus isolated from bats and blood-sucking arthropods
collected in Yunnan Province, China [J]. Microbiol
Spectr, 2023, 11(4): e0512222.
[4] MERTENS P. The dsRNA viruses[J]. Virus Res,
2004, 101(1): 3-13.
[5] BENAVENTE J, MARTÍNEZ-COSTAS J. Avian
reovirus: structure and biology[J]. Virus Res, 2007,
123(2): 105-119.
[6] HOTANI T, TACHIBANA M, MIZUGUCHI H, et al.
Reovirus double-stranded RNA genomes and poly Ⅰ:
C induce down-regulation of hypoxia-inducible
factor 1α[J]. Biochem Biophys Res Commun, 2015,
460(4): 1041-1046.
[7] DUNCAN R. Extensive sequence divergence and
phylogenetic relationships between the fusogenic and
nonfusogenic orthoreoviruses: a species proposal[J].
Virology, 1999, 260(2): 316-328.
[8] ZAMORA P F, HU L Y, KNOWLTON J J, et al.
Reovirus nonstructural protein σNS acts as an RNA
stability factor promoting viral genome replication[J].
J Virol, 2018, 92(15): e00563-18.
[9] TENORIO R, FERNÁNDEZ DE CASTRO I,
KNOWLTON J J, et al. Function, architecture, and
biogenesis of reovirus replication neoorganelles [J].
Viruses, 2019, 11(3): 288.
[10]TENORIO R, FERNÁNDEZ DE CASTRO I,
KNOWLTON J J, et al. Reovirus σNS and μNS
proteins remodel the endoplasmic reticulum to build
replication neo-organelles[J]. mBio, 2018, 9(4):
e01253-18.
[11]LEE C H, RAGHUNATHAN K, TAYLOR G M ,
et al. Reovirus nonstructural protein σNS recruits viral
RNA to replication organelles[J]. mBio, 2021, 12(4):
e0140821.
[12]GORAL M I, MOCHOW-GRUNDY M, DERMODY
T S. Sequence diversity within the reovirus S3 gene:
reoviruses evolve independently of host species,
geographic locale, and date of isolation[J]. Virology,
1996, 216(1): 265-271.
[13]KUMAR R, MEHTA D, MISHRA N, et al. Role of
host-mediated post-translational modifications (PTMs)
in RNA virus pathogenesis[J]. Int J Mol Sci, 2020 ,
22(1): 323.
[14]MOK L, WYNNE J W, GRIMLEY S, et al. Mouse
fibroblast L929 cells are less permissive to infection by
Nelson Bay orthoreovirus compared to other mammalian
cell lines[J]. J Gen Virol, 2015, 96(Pt 7): 1787-1794.
[15]MOK L, WYNNE J W, TACHEDJIAN M, et al.
Proteomics informed by transcriptomics for
characterising differential cellular susceptibility to Nelson
Bay orthoreovirus infection[J]. BMC Genomics, 2017,
18(1): 615.
[16]CHIEN C T, BARTEL P L, STERNGLANZ R, et al.
The two-hybrid system: a method to identify and clone
genes for proteins that interact with a protein of
interest[J]. Proc Natl Acad Sci U S A, 1991, 88(21):
9578-9582.
[17]RAO B C, LI J H, REN T, et al. RPL19 is a
prognostic biomarker and promotes tumor progression in
hepatocellular carcinoma [J]. Front Cell Dev Biol,
2021, 9: 686547.
[18]PETROVA E, GRACIAS S, BEAUCLAIR G, et al.
Uncovering flavivirus host dependency factors through a
genome-wide gain-of-function screen[J]. Viruses, 2019,
11(1): 68.
[19]PAN W, SONG D G, HE W Q, et al. EIF3i affects
vesicular stomatitis virus growth by interacting with
matrix protein[J]. Vet Microbiol, 2017, 212: 59-66.
[20]KOMAROVA A V, REAL E, BORMAN A M, et al.
Rabies virus matrix protein interplay with eIF3, new
insights into rabies virus pathogenesis[J]. Nucleic Acids
Res, 2007, 35(5): 1522-1532.
[21]YIN X J, HONG W, TIAN F J, et al. Proteomic
analysis of decidua in patients with recurrent pregnancy
loss (RPL) reveals mitochondrial oxidative stress
dysfunction[J]. Clin Proteomics, 2021, 18(1): 9.
[22]GUARNIERI J W, DYBAS J M, FAZELINIA H, et al.
Core mitochondrial genes are down-regulated during
SARS-CoV-2 infection of rodent and human hosts[J].
Sci Transl Med, 2023, 15(708): eabq1533.
[23]MUHAMMAD J S, ELGHAZALI G, SHAFARIN J,
et al. SARS-CoV-2-induced hypomethylation of the
ferritin heavy chain (FTH1) gene underlies serum
hyperferritinemia in severe COVID-19 patients [J].
Biochem Biophys Res Commun, 2022, 631: 138-145.
[24]OHTA K, SAKA N, NISHIO M. Human
parainfluenza virus type 2 V protein modulates iron
homeostasis[J]. J Virol, 2021, 95(6): e01861.
猜你喜欢
中成药(2017年8期)2017-11-22 03:19:00
中国校外教育(下旬)(2016年11期)2016-12-27 11:42:42
中国教育信息化·基础教育(2016年10期)2016-12-20 20:09:27
今传媒(2016年11期)2016-12-19 11:35:50
电脑知识与技术(2016年27期)2016-12-15 19:19:25
中国科技纵横(2016年17期)2016-11-30 21:40:27
科技视界(2016年23期)2016-11-04 10:07:53
中国继续医学教育(2015年4期)2016-01-07 07:37:57
中国现代医学杂志(2015年26期)2015-12-23 11:04:22
医学研究杂志(2015年7期)2015-06-22 11:01:42
