
基于广义加性混合模型(GAMM)的沙柳特征因子动态变化
2024-11-11 00:00:00王晓华许昊锁岚马俊杰
中南林业科技大学学报 2024年9期
摘 要:【目的】对沙柳特征因子的动态变化进行研究,分析环境因子影响下的地径、枝高动态变化过程。【方法】采用样地调查、样本采集、气象数据收集等手段,基于广义加性混合模型(GAMM),以灌丛、枝条以及二者的嵌套作为随机效应,探究灌木地径、枝高与土壤水分(SM)、年平均降水量(MAP)、年平均气温(MAT)及年龄等影响因子的动态变化规律。【结果】1)对于不考虑随机效应的广义加性模型(GAM),灌木地径与影响因子呈较强的非线性关系(有效自由度E均大于8.20,且P<0.001),枝高仅与时间呈非线性关系,与其他影响因子均为线性关系。2)相较于GAM,GAMM在随机效应的影响下,地径与各影响因子之间非线性显著降低(E变小),但在以灌丛为随机效应的模型中,地径与年平均气温趋于线性关系(E为1),而枝高与时间的非线性关系更强,与其余影响因子仍呈线性关系。3)考虑随机效应的GAMM比GAM的拟合结果更优,且嵌套模式下的GAMM拟合效果最好。【结论】沙柳不同特征因子对环境因子的响应有差异,而相比枝高,地径的变化程度更大。研究结果有助于掌握沙柳特征因子的动态变化对环境因素的响应机制,为进一步探究沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据。
关键词:沙柳特征因子;环境因子;广义加性模型;广义加性混合模型;动态变化规律
中图分类号:S718.45 文献标志码:A 文章编号:1673-923X(2024)09-0060-11
基金项目:宁夏自然科学基金项目(2022AAC03033);宁夏哲学社会科学规划项目(23NXBGL06)。
Dynamic change of characteristic factors of Salix cheilophila based on generalized additive mixed models (GAMM)
WANG Xiaohuaa, XU Haob, SUO Lana, MA Junjiea
(a. School of Forestry and Heraling; b. School of Economics and Management, Ningxia University, Yinchuan 750021, Ningxia, China)
Abstract:【Objective】The dynamic change of the Salix cheilophila characteristic factors was studied, and the dynamic change process of ground diameter and branch height was analyzed under the influence of environmental factors.【Method】Based on sample survey, sample collection and meteorological data collection, we explored the dynamic variation of shrub diameter, branch height, soil moisture(SM), mean annual temperature (MAT), mean annual precipitation (MAP) and age.【Result】1) For the generalized additive model(GAM) without random effects, diameter had a strong non-linear relationship with the influence factor (effective degrees of freedom E was greater than 8.20, and P<0.001, it was significant), branch height was only non-linear with age, and linear with the other influence factors (E=1); 2) Compared with GAM, GAMM under the influence of random effect, the nonlinearity between ground diameter and each influence factor decreased significantly (E became smaller), but in the model with shrub as the random effect, diameter and MAT tended to be linear relationship (E=1), while the branch height and age were more nonlinear relationship, and its residual factor was still linear relationship; 3) GAMM considering random effects gave a better fit than GAM, and the nested mode.【Conclusion】The different characteristic factors of S. cheilophila had different effect response to the environmental factors, while the ground diameter was more variable than the branch height. This study was helpful to grasp the response mechanism of the dynamic changes of the characteristic factors to environmental factors, and to provide a scientific basis for further exploring the change of plant population, vegetation succession and vegetation management in the process of habitat change in sandy land.
Keywords: characteristic factors of Salix cheilophila; environmental factors; generalized additive models; generalized additive mixed models; dynamic change law
地径、树高等关键特征因子对森林资源的科学评估至关重要,不仅为监测林分的生长状况提供了可靠的数据支持,而且有助于揭示林分生长的动态变化规律[1]。沙柳以其出色的抗逆性、卓越的固沙保土能力以及显著的经济利用价值被誉为沙荒地区生态恢复的先锋树种,是森林生态系统中不可或缺的重要成员[2]。随着灌木研究领域的不断拓展,沙柳等灌木的特征因子已经成为灌木群落结构、功能及其与生态系统相互作用研究的关键指标[3]。
环境变化对灌木生长产生多方面的影响,林分因存在巨大的空间异质性以及受自然和人为扰动的易变性等特点,不同的环境条件下灌木生长过程也会存在差异[4]。通过了解环境影响下灌木特征因子的动态变化,从而对其经营管理进行适应性调整[5]。但目前对于建立更加精确、可靠的动态变化模型没有统一的标准和规范,且以往的模型对于环境因子的考虑多是以附加变量或哑变量的形式考虑。如鞠灵等[6]基于哑变量模型构建了立地因子及其交互作用的生物量模型;闵志强等[7]将竞争等级和立地质量等级作为哑变量逐步参与胸径、树高生长过程拟合;罗鹏飞等[8]在基础模型中引入立地因子构建桉树的胸径哑变量生长模型,均得到考虑哑变量进行研究模型精度可显著提高。但传统模型只考虑了变量间的线性关系,显然是不足的[9]。有研究发现,非线性模型是研究环境影响的一个有效工具[10]。宁金魁等[11]通过建立回归模型发现个体大小、试验林、林层等因素对阔叶复层次生林胸高断面积年均生长量(BAI)存在显著差异。近年来,一些研究引入了环境因子去建立广义加性模型(GAM)并对特征因子动态变化进行研究。例如,何培等[12]结合温度、降水量变化构建了广义加性模型,研究自变量与因变量之间的非线性关系;赵俊卉[13]将立地因子(海拔、坡度等)引入异龄林的生长动态广义加性模型,得到了较好的结果;张晓玉等[14]利用广义加性模型建立了不同微地貌类型和深度的土壤水分与地形-植被因子之间的关系,得到了较好的结果。以上研究证明,基于气候因子的广义加性模型明显优于基础模型,但并未考虑随机效应的影响。有学者提出,引入随机效应构建广义加性混合模型(GAMM,其能够对非线性、非正态、非独立分布的数据建模,使得模型更加灵活,可捕捉到真实数据中的复杂关系)可以显著提高模型的精准性。例如李春明等[15]考虑样地的随机效应,并与传统模型的模拟效果进行比较,得到了更加精准的模型。肖武鹏[16]将采样站位作为随机效应,充分考虑与固定效应之间的交互作用,构建了浮游植物各类群关于解释变量的最优GAMM。
为了探究灌木特征因子动态变化与环境因子间的关系,本研究以宁夏盐池县的沙柳为研究对象,以地径(D)、枝高(H)作为因变量,以年龄(t)、土壤水分(SM,SM)、年平均降水量(MAP,MAP)、年平均气温(MAT,MAT)作为自变量,建立广义加性模型(GAM),并在此基础上引入灌丛、枝条、嵌套模式作为随机效应建立广义加性混合模型(GAMM),分析环境因子对灌木沙柳生长过程的影响,可对研究环境变化的响应机制和适应策略给予参考,为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据。
1 数据与方法
1.1 研究区概况
宁夏盐池县位于宁夏回族自治区东部(37°04′~38°10′E,106°30′~107°47′N),地势南高北低,北接毛乌素沙漠[17],属鄂尔多斯台地,南靠黄土高原,属典型的过度地带。该地日照长,温差大,干旱少雨,光能丰富,日照充足,气候为温带大陆性半干旱气候[18]。全县干旱少雨,蒸发量大,年平均降水量为295 mm,降水量由南向北递减[19]。土壤类型主要是灰钙土,其次是黑垆土和风沙。其中南部黄土高原丘陵区以黑垆土为主,其次为灰钙土,北部鄂尔多斯缓坡丘陵区以风沙土、灰钙土为主[20]。主要植被有达乌里胡枝子Lespedeza davurica、短花针茅Stipa breviflora、白草Pennisetum centrasiaticum、赖草Leymus secalinus、老瓜头Cynanchum komaroviil、柠条Caragana korshinskii、沙柳Salix cheilophila等[21]。
1.2 数据来源
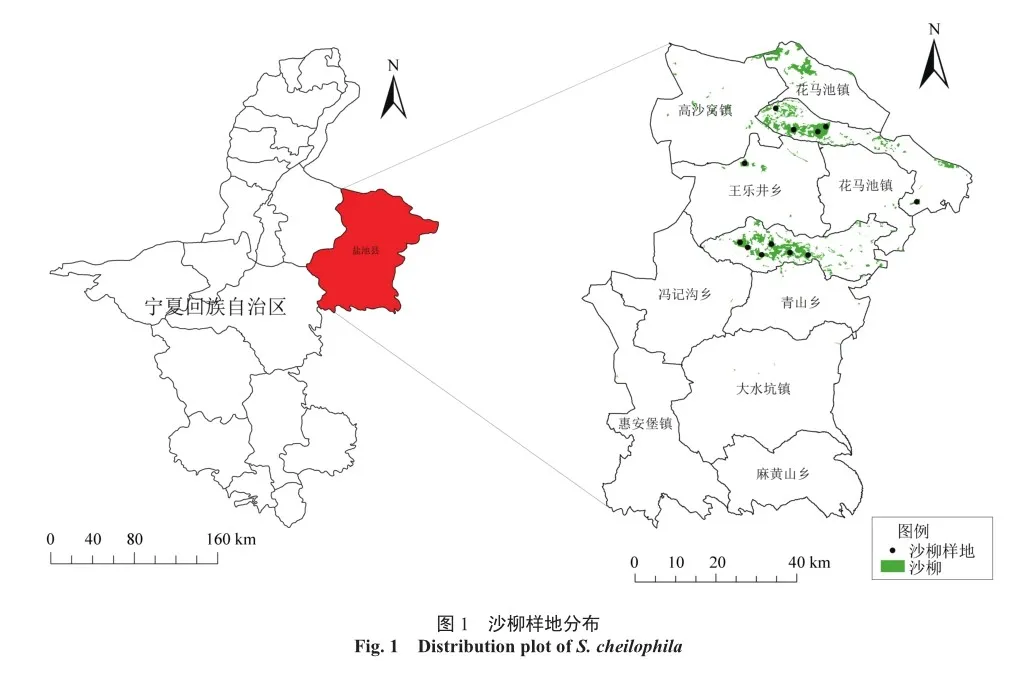
沙柳抗旱性强,是盐池地区水土保持和固沙造林的重要灌木树种之一[22],对盐池沙漠地区进行生态修复有着明显效果,及时采取有效措施,不仅可以保障沙柳林区生态功能的持续发挥,还能为当地获得可观的生物资源,实现生态、经济效益的双丰收[23]。本研究根据盐池县沙柳分布情况,共设置20 m×20 m样地12个(图1),并对样地进行信息采集。每个样地根据调查数据选择2~3株标准丛进行样本采集,共采集30丛灌木,调查灌丛的地径、枝高、分枝数以及地理坐标等(表1)。在每一标准丛中,选取3~4个标准枝,共选择94个标准枝,从基部起每隔20 cm制取圆盘,经打磨抛光后圆盘表面年轮界线清晰,使用WinDENDRO软件进行扫描,测量地径年生长量,求得枝高年生长量,并计算年龄[24]。SM、MAP、MAT等数据基于Terra Climate的数据集,利用QGIS将研究区域内20年的动态变化数据进行提取整理后获得。
1.3 研究方法
1.3.1 GAM及GAMM的构建
广义加性模型(generalized additive models,简称GAM)的拟合主要考虑使用不同的光滑样条函数[25],模型中的每一加项可解释因变量如何随自变量变化而变化,模型也可兼顾自变量较多且存在非线性关系的情况。随着研究的不断深入,广义加性模型在林业上进一步得到应用[26],成为分析气候因子影响的一个有效工具。比起线性模型,能更好地体现自变量与因变量之间复杂的非线性关系[27]。本研究将沙柳的年龄、土壤水分、年平均降水量、年平均温度等因子作为自变量,地径、枝高等因子作为因变量,建立广义加性模型。



2 结果与分析
2.1 基于GAM对地径与枝高动态变化过程的拟合
由表2可知,地径与所有环境因子之间均存在显著的非线性关系(E>8.2),且均具有极显著性意义(P<0.001),其中,地径与年平均降水量之间的关系最为显著(P<0.001,F=11.37),这表明年平均降水量在地径生长方面起着至关重要的作用。对于枝高而言,GAM的分析结果表明,它仅与年龄这一因子呈现显著的非线性关系(E=6.7),并具有极显著意义(P<0.001)。相比之下,枝高与土壤水分、年平均降水量、年平均温度之间的关系表现为线性(E=1),并且这种线性关系并不显著(P>0.05),说明这些环境因子对枝高的影响相对有限。

由图2可知,地径生长量随着沙柳年龄的增长而持续上升,表现出明显的正相关性,在不同环境因子的作用下波动较显著,这表明环境因子与地径年生长量之间存在较强的非线性关系。当土壤水分达到6 mm时,地径年生长量达到峰值;而当土壤水分增至8 mm左右时,生长量降至最低点,这一现象可能与土壤水分在不同阶段对植物生长的不同作用机制有关。随着年平均温度升至2.5 ℃,地径年生长量达到最大值,这可能与沙柳在该温度范围内的生理活动最为活跃有关。年平均降水量对地径年生长量的影响则呈现一定的波动性,在年降水量约为250 mm时,地径年生长量降至最低,而当年降水量为290~340 mm时,地径年生长量保持在较高水平。这表明适量的降水对沙柳的生长是有益的,但过量或过少的降水都可能对其生长产生不利影响。对于枝高年生长量而言,随着年龄的增长,其后期呈现出缓慢的增长趋势,整体形态接近“S”形生长曲线,这表明枝高的生长速度在成熟期逐渐减缓。此外,在环境因子的影响下,枝高年生长量的反应并不敏感,这可能意味着枝高的生长更多受到遗传因素和个体差异的控制。
整体来看,地径生长过程中整个灌丛以及枝条之间的差异更加显著,枝高生长过程受环境因子影响无明显变化。
2.2 基于GAMM对地径与枝高动态变化过程的拟合
2.2.1 以灌丛作为随机效应
由表3可知,在引入灌丛作为随机效应后,所有因子与地径仍然保持着极显著的关系(P< 0.001),在这些因子中,年龄对地径的影响最为显著(P<0.001,F=507.99),环境因子中年平均温度影响最为明显(P<0.001,F=14.57);对于枝高而言,引入灌丛作为随机效应后其与各因子之间的显著性关系并未发生明显变化,环境因子对枝高的影响依旧不显著(P>0.05),这表明枝高的生长可能更多地受到非环境因素的影响,如遗传特性或内在生理过程。进一步分析地径与各因子之间的非线性关系,发现引入随机效应灌丛后,地径与土壤水分之间的非线性关系得到了加强(E=7.72),与年龄和年平均降水量的非线性关系次之(E=7.24,E=5.79),年平均温度与地径之间的关系在引入随机效应后趋于线性(E=1,表3),这可能意味着在考虑灌丛间差异后,温度对地径生长的影响变得更加不敏感。对于枝高,与年龄之间的非线性关系在引入灌丛后有所增强(E=7.38),而与环境因子之间的关系则趋于线性(E=1)。

如图3所示,考虑了灌丛的随机效应之后,发现不同环境因素对地径生长的影响整体上有所减弱,而枝高的增长在环境因素影响下没有表现出明显的变化。年平均温度对地径的影响以及年平均降水量和土壤水分对枝高的影响都显示出趋向于线性的关系。在灌木生长过程中,随机效应在后期变得更加显著,这表明在环境条件较为优越的情况下,不同灌丛之间的地径和枝高的年生长量差异变得更加明显。随着时间增长到第10年,灌丛之间的差异性开始显著增加,枝高的“S”形生长模式也变得更加明显;土壤水分对地径的影响在中期尤为显著,特别是当地径年生长量达到6 mm时,不同灌丛之间的差异最为明显。而对于枝高而言,不同灌丛在后期的生长差异较大;随着年平均温度的升高,不同灌丛之间地径和枝高的生长量在生长的早期也表现出显著的区别;此外,年平均降水量在生长早期对地径的影响较大,这一时期不同灌丛之间的生长差异尤为显著,枝高在前期与后期也表现出较大的差异性。
整体来看,引入随机效应灌丛后,特征因子生长变化幅度缩小,其中年平均温度与年龄之间由非线性关系趋于线性关系。
2.2.2 以枝条作为随机效应
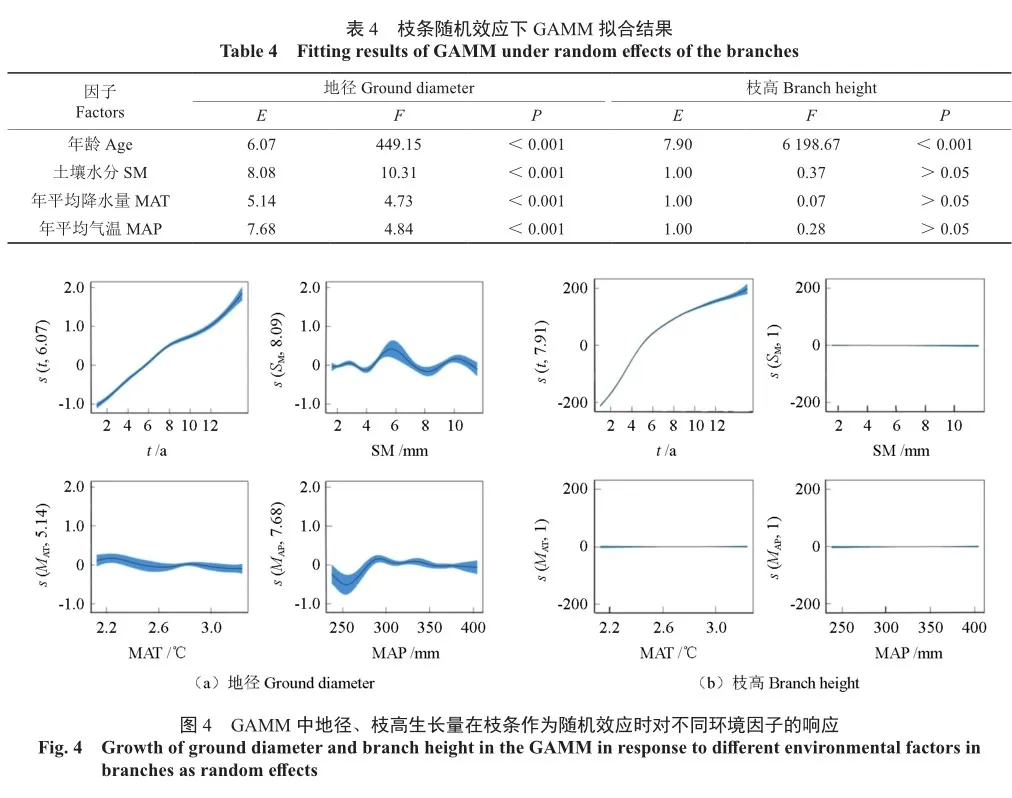
由表4可知,引入枝条作为随机效应后,各因子对地径以及枝高的显著情况均不变,其中土壤水分成为对地径最具有显著性意义的因子(F=10.31);非线性关系中,年平均温度与地径又转变为非线性关系(E=5.14)。
由图4可知,在考虑了枝条作为随机效应后,在各因子影响下地径、枝高生长过程无太大变化,变化范围略微增大;在年平均温度影响下,枝条地径生长出现波动,枝高生长随时间变化更加接近“S”形。
2.2.3 以嵌套模式作为随机效应
由表5可知,引入嵌套模式作为随机效应后,相比枝条作为随机效应,所有因子显著性非线性关系无明显变化,但E值与F值均有所提高。
由图5可知,在考虑了嵌套模式作为随机效应后,各因子影响下特征因子的生长情况与考虑随机效应-枝条影响时无明显变化,随着环境因子的增加,地径与枝高的变化趋势和变化节点基本保持一致。
2.3 模型评价分析
由表6可以看出,无论对于沙柳地径还是枝高生长,构建GAM拟合的非线性模型表达特征因子动态变化效果较好,在考虑了随机效应后建立的GAMM拟合结果更加精准,其中,嵌套模式作为随机效应时adj-R2值最大,AIC、AMR均为最小值,因此嵌套模式下的GAMM最优。

通过建立GAMM,研究结果揭示了特征因子与多种环境因子之间存在的显著非线性关系。在地径生长的过程中,年龄、土壤水分、年平均降水量以及年平均温度对地径的影响呈现出递减的顺序,其中土壤水分对地径的影响最为显著,且环境因子与地径之间的关系均具有极显著相关性。对于枝高的生长而言,其与年龄的关系呈现出明显的非线性特征,并且这种关系在统计学上具有极显著意义。相比之下,枝高与年平均温度、土壤水分以及年平均降水量之间的关系则更趋于线性,环境因子对枝高的影响并不显著。
在相同的环境条件下,地径对环境变化因素(如土壤水分、年平均温度和年平均降水量)表现出更高的敏感性,而枝高对这些环境变化因素的响应则显得较为迟钝。这一发现表明,在灌木生长的研究中,地径可能是一个更为敏感和有效的生物指标,可用于评估环境因素对灌木生长的影响。而枝高的生长变化研究可能需要参考更长的时间跨度或特定的环境条件。综上所述,本研究的GAMM分析结果为理解灌木生长与环境因子之间复杂的相互作用提供了新的视角,并为生态学和植物生理学领域的进一步研究奠定了理论基础。
3 结论与讨论
3.1 讨 论
地径与环境的非线性关系较强,而枝高与环境因子趋于线性关系,这与王子婷等[29]、杨建伟等[30]的研究结果一致。干旱地区环境对灌木地径的影响大于对灌木枝高的影响,可能有以下原因:
1)水分胁迫。干旱环境下,水分稀缺是一个主要限制因素。孟诗原[31]的研究指出水分胁迫对卫矛属植物的高生长和地径生长均有影响,可减缓植物生长,而植物的变化程度与其对水分胁迫的抗性有关,抗逆性较强则受到的影响较小,特征因子的变化程度相应减小,而抗逆性较弱则受到的影响较大,特征因子的变化幅度也相应增高。结合本研究的结果来看,灌木地径的增长通常受到较少水分供应的限制,因为较宽的直径能够存储更多的水分和养分,以供植物在干旱时期使用,相比之下,灌木的枝高增长更容易受到水分限制,因为较高的枝干需要更多的水分来支持生长[32],因而在应对水分胁迫时枝高的抗逆性是比较强的,与环境因子呈线性关系,而地径的抗逆性相对较弱,与环境因子之间呈非线性关系。
2)养分利用。不同灌木对坡面地形所作出的生长反应也会存在差异性,主要影响因素就包括养分条件,在干旱地区土壤中的养分通常也较为稀缺。由于灌木地径较大,它能够在干旱条件下存储更多的养分,以支持其生长和维持基本代谢活动[33]。而灌木的枝高增长也可能受到养分供应的限制,因为较高的枝干需要更多的养分来维持生长。灌木可以通过调控生长和器官之间的营养物质分配来应对各种变化,因此地径与枝高响应出现差异。这与杨建伟等[30]的研究结果一致。
3)生物适应策略。灌木通常具有较低的生长速率和较高的生存能力,这是一种在干旱环境中的适应策略。较低的生长速率意味着灌木地径的增长相对稳定,可以更好地适应干旱条件下的水分限制。而较快的枝高增长可能会增加水分蒸散和养分消耗,这对于灌木来说在干旱环境中可能是不利的[34]。例如赵龙等[35]的研究表明,针对不同灌木的生长速率,限制灌木生长方向是不同的。其中,生长较快的灌木主要限制株高和冠幅宽度的生长。结合本研究,之所以枝高对环境变化响应敏感性略弱,可能是因为研究所选取的灌木体形偏中上。
因此,在研究区进行人工沙柳种植时,不仅要结合立地条件,也要对土壤水分与养分的植被承载力进行判断,从而把控植株生长不同阶段的植株密度和空间配置,目的是维持人工植被恢复的可持续性。另外,在植被的不同恢复阶段实施定期的管理措施(如平茬、间伐等)是有必要的,这能够为灌木生长提供更良好的生境,保障灌木最大的生态效益、环境效益,更有效地抑制林地退化导致的水土流失[36]。
本研究只考虑了土壤水分、年平均气温、年平均降水量等3种立地因子的影响,而海拔、坡位、坡度等立地因子同样对灌木生长有不同程度的影响[37]。除此之外,灌木本身研究条件受限,因年轮不清晰、平茬等问题使取得的数据存在误差,最后得到的结果准确性不高。
针对这些局限性,未来的研究计划将考虑包括海拔、坡位和坡度等其他可能影响灌木动态变化的因素。同时,研究也将探索更加先进且适合灌木生长研究的模型,以期提高研究的准确性和可靠性。通过综合考虑更多的影响因素,并采用更先进的研究方法,更全面地理解灌木生长的动态变化,为灌木的管理和保护提供更加科学的依据。
3.2 结 论
本研究选取宁夏盐池县的灌木沙柳作为研究对象,通过GAM和GAMM对其特征因子(地径和枝高)的动态变化过程进行了深入研究。在模型建立中,年龄、土壤水分、年平均温度和年平均降水量作为自变量,地径和枝高作为因变量。在GAM的基础上,进一步考虑了灌丛、枝条以及嵌套模式作为随机效应,以建立GAMM进行分析。
1)在GAM中,地径与年龄、土壤水分、年平均温度和年平均降水量之间存在较强的非线性关系,而枝高则主要与年龄呈非线性关系,与土壤水分、年平均温度和年平均降水量的关系则更接近线性。地径与各因子之间具有极显著的相关性(P<0.001),而枝高仅与年龄有显著的相关性(P<0.001)。此外,不同灌丛和枝条的特征因子在生长的后期表现出明显的差异,且变化范围较大。
2)GAMM相较于GAM,在考虑随机效应后,地径和枝高的动态变化幅度有所减小,各因子之间的非线性关系显著降低。特别是在随机效应灌丛中,年平均温度与地径的关系趋于线性。当进一步考虑灌丛、枝条和嵌套效应作为随机效应时,各因子的E值上升,而P值下降。
3)综合考虑随机效应的GAMM比传统的GAM得到了更优的拟合结果,尤其是嵌套模式下的GAMM拟合效果最佳。这表明,通过引入随机效应,模型能够更准确地捕捉到灌木沙柳生长过程中的变化,从而提供更为精确的生长动态描述。
研究结果不仅加深了对灌木沙柳生长动态的理解,而且为灌木生态系统的管理和保护提供了科学依据。未来研究可进一步探索其他潜在影响因子,以期构建更为全面和精确的灌木生长模型。
参考文献:
[1] 项文化,田大伦,闫文德.森林生物量与生产力研究综述[J].中南林业调查规划,2003,22(3):57-60. XIANG W H, TIAN D L, YAN W D. Review of researches on forest biomass and productivity[J]. Central South Forest Inventory and Planning,2003,22(3):57-60.
[2] FANG J M, WU F Z, YANG W Q, et al. Effects of drought on the growth and resource use efficiency of two endemic species in an arid ecotone[J]. Acta Ecologica Sinica,2012,32(4):195-201.
[3] CHENG L, WU B, PANG Y J, et al. Shrub growth improves morphological features of nebkhas: a case study of Nitraria tangutorumin the Tengger Desert[J]. Plants,2024,13(5):624.
[4] SUN S C, JIN D M, SHI P L. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship[J]. Annals of Botany, 2006,97(1):97-107.
[5] CHEN A, XU C, ZHANG M, et al. Cross-scale mapping of above-ground biomass and shrub dominance by integrating UAV and satellite data in temperate grassland[J]. Remote Sensing of Environment,2024,304:114024.
[6] 鞠灵,许昊,金学娟,等.基于立地因子及其交互作用的柠条生物量模型研究[J].西北林学院学报,2023,38(2):1-7. JU L, XU H, JIN X J, et al. Establishment of the biomass models of Caragana korshinskii based on site factors and their interaction[J]. Journal of Northwest University of Forestry,2023,38(2): 1-7.
[7] 闵志强,胡云云,王得军,等.基于哑变量的秦巴山区天然栎类林胸径和树高生长模型研究[J].林业资源管理,2020(5): 89-99. MIN Z Q, HU Y Y, WANG D J, et al. Study on growth model of DBH and height based on dummy variable for natural oak forest in Qinba mountain area[J]. Forestry Resources Management,2020(5): 89-99.
[8] 罗鹏飞,龙时胜.基于立地哑变量的桉树胸径生长模型构建[J].福建林业科技,2021,48(3):62-67. LUO P F, LONG S S. Developing a DBH growth model of Eucalyptus based on site dummy variable[J]. Journal of Fujian Forestry Science and Technology,2021,48(3):62-67.
[9] 王占军,许昊,何建龙,等.干旱风沙区沙柳灌木生物量可加性动态模型构建[J].东北林业大学学报,2019,47(9):52-57. WANG Z J, XU H, HE J L, et al. Additive dynamic biomass models for Salix psammophila in the arid sandy area in Ningxia[J]. Journal of Northeast Forestry University,2019,47(9): 52-57.
[10] ZALATAN R, GAJEWSKI K. Dendrochronological potential of Salix alaxensis from the Kuujjua River area, western Canadian arctic[J]. Tree-Ring Research,2006,62(2):75-82.
[11] 宁金魁,王光玉,许诺,等.基于分位数回归模型的阔叶次生林单木胸高断面积生长研究[J].中南林业科技大学学报, 2023,43(12):24-34. NING J K, WANG G Y, XU N, et al. Study on growth of basal area increment of individual trees in broad-leaved secondary forest based on quantile regression model[J]. Journal of Central South University of Forestry & Technology,2023,43(12):24-34.
[12] 何培,辛士冬,姜立春.基于广义加性模型的樟子松树干削度方程研建[J].北京林业大学学报,2020,42(12):1-8. HE P, XIN S D, JIANG L C. Research on stem taper equation of Scots pine based on generalized additive model[J]. Journal of Beijing Forestry University,2020,42(12):1-8.
[13] 赵俊卉.长白山云冷杉混交林生长模型的研究[D].北京:北京林业大学,2010. ZHAO J H. Growth modeling for spruce-fir forest in Changbai Mountain[D]. Beijing: Beijing Forestry University,2010.
[14] 张晓玉,张定海,宁婷,等.古尔班通古特沙漠土壤水分-地形-植被关系研究[J].草地学报,2024,32(5):1548-1557. ZHANG X Y, ZHANG D H, NING T, et al. Study on the relationship between soil moisture, topography and vegetation in the Gurbantunggut Desert[J]. Acta Agrestia Sinica,2024,32(5): 1548-1557.
[15] 李春明,张会儒,王卓晖.基于混合效应模型及生存分析方法的落叶松云冷杉林单木生存模型研究[J].北京林业大学学报, 2022,44(1):1-8. LI C M, ZHANG H R, WANG Z H. Study on single tree survival model of mixed stands of Larix olgensis, Abies nephrolepis and Picea jazoensis based on mixed effect model and survival analysis method[J]. Journal of Beijing Forestry University,2022,44(1):1-8.
[16] 肖武鹏.广义加性混合模型(GAMMs)应用于东海浮游植物群落动态研究[D].厦门:厦门大学,2013. XIAO W P. Generalized additive mixed models (GAMMs) were applied to study the dynamics of phytoplankton communities in the East Sea[D]. Xiamen: Xiamen University,2013.
[17] 许昊,吴宏玥,王占军,等.立地条件对宁夏中部干旱风沙区柠条生物量分配格局的影响[J].北京林业大学学报,2020, 42(12):91-100. XU H, WU H Y, WANG Z J, et al. Effect of site conditions on biomass allocation patterns of Caragana korshinskii in arid aeolian sand regions, middle area of Ningxia of northwestern China[J]. Journal of Beijing Forestry University,2020,42(12): 91-100.
[18] 李婷婷.荒漠草原柠条沙柳灌木林生长特征与土壤水分关系[D].银川:宁夏大学,2019. LI T T. Relationship between growth characteristics of different artificial shrubs and soil moisture in desert steppe[D]. Yinchuan: Ningxia University,2019.
[19] ISHII H, CLEMENT J P, SHAW D C. Branch growth and crown form in old coastal Douglas-fir[J]. Forest Ecology and Management,2000,131(1-3):81-91.
[20] WANG Y S, QU L L, WANG J Y, et al. Sustainable revitalization and green development practices in China’s Northwest arid areas: a case study of Yanchi County, Ningxia[J]. Land,2022,11(11): 1902-1902.
[21] 路丽宁,张定海,李功麟.古尔班通古特沙漠固沙灌木的冠幅预测模型[J].生态科学,2022,41(5):81-89. LU L N, ZHANG D H, LI G L. Crown prediction models of the sand-fixing shrubs in Gurbantunggut Desert[J]. Ecological Science,2022,41(5):81-89.
[22] 姚雪玲,姜丽娜,李龙,等.浑善达克沙地6种灌木生物量模拟[J].生态学报,2019,39(3):905-912. YAO X L, JIANG L N, LI L, et al. Biomass simulation of six shrub species in Otindag sandy land[J]. Acta Ecologica Sinica, 2019,39(3):905-912.
[23] UROSEVIC J, STANKOVIC D, JOKANOVIC D, et al. Phytoremediation potential of different genotypes of Salix alba and S. viminalis[J]. Plants,2024,13(5):735.
[24] 王佳燕.海南东寨港几种红树植物主要生长特征因子间的关系研究[D].海口:华南热带农业大学,2007. WANG J Y. Analysison relationship among majoy factors of several mangroves in Dongzhai harbour, Hainan island[D]. Haikou: South China University of Tropical Agricultural,2007.
[25] 王昕.现代统计分析方法及其应用[J].大众投资指南, 2019(20):236-237. WANG X. Modern methods of statistical analysis and their applications[J]. Popular Investment Guide,2019(20):236-237.
[26] 余黎,雷相东,王雅志,等.基于广义可加模型的气候对单木胸径生长的影响研究[J].北京林业大学学报,2014,36(5): 22-32. YU L, LEI X D, WANG Y Z, et al. Impact of climate on individual tree radial growth based on generalized additive model[J]. Journal of Beijing Forestry University,2014,36(5): 22-32.
[27] 李杰,张鹏,王腾,等.基于广义加性模型研究影响罩网沉降性能的因子[J].南方水产科学,2021,17(4):74-81. LI J, ZHANG P, WANG T, et al. A study on settlement performance of falling-net based on GAM[J]. South China Fisheries Science,2021,17(4):74-81.
[28] 王军锋,张丽华,赵锐锋,等.荒漠草原区不同生活型植物生长对降水变化的响应[J].应用生态学报,2020,31(3):778-786. WANG J F, ZHANG L H, ZHAO R F, et al. Responses of plant growth of different life-forms to precipitation changes in desert steppe[J]. Chinese Journal of Applied Ecology,2020,31(3): 778-786.
[29] 王子婷,杨磊,蔡国军,等.半干旱黄土区坡面尺度柠条生长状况及影响要素分析[J].生态学报,2017,37(23):7872-7881. WANG Z T, YANG L, CAI G J, et al. Spatial patterns of Caragana korshinskii growth on hillslope scale and influencing factors in the semi-arid Loess Plateau[J]. Acta Ecologica Sinica, 2017,37(23):7872-7881.
[30] 杨建伟,孙桂芳,赵丹,等.干旱胁迫对3种灌木生长及叶水分生理的影响[J].林业与生态科学,2018,33(4):423-428. YANG J W, SUN G F, ZHAO D, et al. Effects of drought stress on the growth of three shrubs and their leaf water physiology[J]. Forestry and Ecological Sciences,2018,33(4):423-428.
[31] 孟诗原.低温和水分胁迫对卫矛属主要树种生长及生理特性的影响[D].泰安:山东农业大学,2019. MENG S Y. Effects of cold and water stress on growth and physiology characteristics of major species in Euonymus Linn [D]. Taian: Shandong Agricultural University,2019.
[32] 闵梓骁,张建新,范文波,等.不同立地条件下沙棘土壤水分分布特征及动态生长研究[J].水土保持学报,2022,36(4): 204-210. MIN Z X, ZHANG J X, FAN W B, et al. Soil moisture distribution and dynamic growth of Hippophae rhamnoides under different site conditions[J]. Journal of Soil and Water Conservation, 2022,36(4):204-210.
[33] LACERTE V, LAROCQUE G R, WOODS M, et al. Calibration of the forest vegetation simulator (FVS) model for the main forest species of Ontario, Canada[J]. Ecological Modelling, 2006,199(3):336-349.
[34] AUDUN ?. In-forest assessment of timber stiffness in Norway spruce (Picea abies (L.) Karst.)[J]. European Journal of Wood and Wood Products,2013,71(4):429-435.
[35] 赵龙,郭忠升,郭满才,等.黄土丘陵半干旱区柠条林生长过程研究[J].水土保持通报,2012,32(6):192-195. ZHAO L, GUO Z S, GUO M C, et al. Growing process of Caragana microphylla shrub land in semi-arid hilly region of Loess Plateau[J]. Bulletin of Soil and Water Conservation,2012,32(6): 192-195.
[36] 郝珉辉,张忠辉,赵珊珊,等.吉林蛟河针阔混交林树木生长与生境的关联性[J].生态学报,2017,37(10):3437-3444. HAO M H, ZHANG Z H, ZHAO S S, et al. Habitat associations of tree growth in a coniferous and broad-leaved mixed forest in Jiaohe, Jilin province[J]. Acta Ecologica Sinica,2017,37(10): 3437-3444.
[37] 程杰,刘永辉,田瑛.宁夏半干旱区柠条锦鸡儿灌木林生长特征[J].水土保持通报,2016,36(1):332-336. CHENG J, LIU Y H, TIAN Y. Growth characteristics of Caragana korshinskii shrubbery in semi-arid regions of Ningxia Hui Autonomous region[J]. Bulletin of Soil and Water Conservation, 2016,36(1):332-336.
[本文编校:谢荣秀]
