
铁杉属花粉形态及其聚类分析研究
2024-09-12 00:00:00陈芷涵尚鑫谢淦张鑫
南京林业大学学报(自然科学版) 2024年4期



摘要:【目的】对铁杉属(Tsuga)现存所有10种植物花粉形态特征进行观察比较,结合花粉化石分析铁杉属系统发育关系及花粉形态的演化过程。【方法】通过扫描电子显微镜(SEM)观察铁杉属的花粉形态特征,并利用7个定量指标和1个定性指标进行系统聚类分析(HCA)。【结果】铁杉属花粉为N1P3C1型花粉,远极面具薄壁区,赤道轴长度为21.20~58.00 μm,多无气囊,稀有气囊;花粉粒表面多疣状,稀微疣状,表面有棘突或无。系统聚类分析显示铁杉属花粉可以分类为3类:①有气囊,无棘突类:山地铁杉(T. mertensiana)与长苞铁杉(Nothotsuga longibracteata);②无气囊,无棘突类:卡罗莱纳铁杉(T. caroliniana)、加拿大铁杉(T. canadensis)与化石种2 (T. sp.2);③无气囊,有棘突类: 异叶铁杉(T. heterophylla)、郁林铁杉(T. ulleungensis)、米铁杉(T. diversifolia)、丽江铁杉(T. forrestii)、日本铁杉(T. sieboldii)、云南铁杉(T. dumosa)、铁杉(T. chinensis)、化石种1(T. sp.1 )和化石种3 (T. sp.3 )。【结论】铁杉属花粉是从有气囊向无气囊,无棘突向有棘突演化的,且花粉特征与分布地有密切关系,同一地区分布的花粉形态相似。基于花粉形态的聚类分析结果与分子系统发育树大体符合,可以通过花粉形态对铁杉属植物及化石进行物种鉴定,并为其系统发育研究提供参考。
关键词:铁杉属;花粉形态;扫描电镜;聚类分析
中图分类号:Q944.5"""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)04-0037-09
Phylogenetic relationship and evolutionary patterns of Tsuga pollen morphology: a cluster analysis-based study
CHEN Zhihan1,3, SHANG Xin1,2, XIE Zin2, ZHANG Xin1*
(1. College of Forestry, Northwest Aamp;F University, Yangling" 712100,China; 2. Institute of Botany, Chinese Academy of Sciences, Beijing" 100093," China; 3. Kunming Institute of Botany, Chinese Academy of Sciences," Kunming" 650201, China" )
Abstract: 【Objective】 To analyze the phylogenetic relationship and the evolution pattern of pollen morphology in Tsuga, pollen morphology characteristics of all 10 extant species of Tsuga were observed and compared in combination with pollen fossils.【Methods】 Pollen morphology characteristics of Tsuga were observed by scanning electron microscopy (SEM), and hierarchical cluster analysis (HCA) was performed using seven quantitative indicators and one qualitative indicator. 【Results】 Pollen of Tsuga belongs to the N1P3C1 type, leptoma on the distal face, the equatorial length ranging from 43.74 to 95.30 μm, mostly without sacci and rare saccate, surface mostly warty and sparsely micro-warty, surface with spines or absent. According to the hierarchical cluster analysis, Tsuga can be divided into three categories: ①T. mertensiana and Nothotsuga longibracteata; ②T. caroliniana and T. canadensis; ③T. heterophylla, T. ulleungensis, T. diversifolia, T. forrestii, T. sieboldii, T. dumosa and T. chinensis. 【Conclusion】 Tsuga pollen evolved from having sacci to without sacci, from no spines to having spines. The pollen characteristics of Tsuga had a close relationship with the distribution place, which means that pollen distributed in the same region were similar. The clustering analysis results based on pollen morphology is largely consistent with the molecular phylogenetic tree, which can be used to identify extant species and fossils of Tsuga and provide reference for Tsuga phylogenetic studies.
【Objective】 To analyze the phylogenetic relationships and evolutional patterns of pollen morphology in Tsuga, we observed and compared the pollen characteristics of all 10 extant species of Tsuga along with pollen fossils. 【Method】 Pollen morphology was examined using scanning electron microscopy (SEM), and hierarchical cluster analysis (HCA) was conducted using seven quantitative indicators and one qualitative indicator. 【Result】 Tsuga pollens typically exhibits characteristics of the N1P3C1 type, with a leptoma on the distal face. The equatorial length ranges from 21.20 to 58.00 μm, predominantly lacking sacci but occasionally with echinae saccate forms. The pollen surface is typically warty or sparsely micro-warty,"" with spines or without. According to the hierarchical cluster analysis, Tsuga could be divided into three categories: ①saccate but lacking echinae type: T. mertensiana and Nothotsuga longibracteata; ②lacking sacci and echinae type: T. caroliniana, T. canadensis and pollen fossil T. sp.2; ③lacking sacci but echinate type: T. heterophylla, T. ulleungensis, T. diversifolia, T. forrestii, T. sieboldii, T. dumosa, T. chinensis and pollen fossil T. sp.1, T. sp.3.【Conclusion】 Pollen morphology in Tsuga has evolved from saccate to non-saccate forms, and from lacking spines to possessing spines. These pollen characteristics are closely related to geographic distribution, indicating similarity among pollen from the same regions. The clustering analysis based on pollen morphology largely aligns with molecular phylogenetic trees, offering a method to distinguish extant species and fossils of Tsuga and providing valuable insights for phylogenetic studies of Tsuga.
Keywords:Tsuga; pollen morphology; scanning electron microscopy(SEM); cluster analysis
铁杉属(Tsuga)隶属于松科,为东亚—北美间断分布类群[1]。铁杉属植物的主要鉴定特征为苞鳞形态、珠鳞形态、叶型和花粉形态[2-5]。现存铁杉属植物共10种,其中丽江铁杉(T. forrestii)、米铁杉(T. diversifolia)、日本铁杉(T. sieboldii)、铁杉(T. chinensis)、云南铁杉 (T. dumosa)和郁陵铁杉[6](T. ulleungensis)6种分布于东亚地区,而山地铁杉(T. mertensiana)、加拿大铁杉(T. canadensis)、卡罗来纳铁杉(T. caroliniana)、异叶铁杉(T. heterophylla)4种分布于北美地区[7- 8]。长苞铁杉(Nothotsuga longibracteata)是中国特有种,因其诸多分类特征与铁杉属植物相似,早期的分类学家将其归入铁杉属[9-10],近来分子系统学证据表明,长苞铁杉应独立为长苞铁杉属(Nothotsuga)[11]。
分子系统学的发展为阐明该属的系统发育提供了更多的思路[12]。Havill等[13]利用核糖体ITS和叶绿体DNA数据分析了铁杉属的系统发育和生物地理学。Feng等[14]利用881个核基因、60个叶绿体基因和23个线粒体基因重建铁杉属植物高分辨率的系统发育关系。叶绿体基因组分析结果表明[15],铁杉可能不是一个单系物种,有必要对其展开进一步系统发育学研究。众多学者从形态学[2,5,16-21]、分子系统学[11,14-15]等角度对铁杉属进行了研究,探讨了铁杉属的系统发育关系,为深入研究铁杉属系统发育框架奠定了基础[9,22]。
花粉表面具有形态各异的萌发沟和表面纹饰,且有关花粉形态的基因序列具有高度保守性和稳定性,是种上分类阶元划分的重要参考[23-24]。例如,利用花粉形态特征可将顶冰花属(Gagea)分为两组[25];通过形态聚类分析,朱槿(Hibiscus rosa-sinensis)[26]、风毛菊属(Saussurea)[27]、睡莲属(Nymphaea)[28]的分类和进化地位得以进一步确定。
铁杉属花粉具有一定的分类意义,但有关铁杉属花粉研究较少。孙爱芝等[29]只将铁杉属花粉描述为具周囊类型,张金谈[30]观察了铁杉属铁杉与云南铁杉2种植物花粉,发现铁杉属花粉粒无气囊,有皱边,皱边表面具有蠕虫状纹饰和小刺,与王伏雄等[31]对云南铁杉的观察结果一致。Sivak[2]用光学显微镜和扫描电子显微镜详细研究了铁杉属12种花粉粒表面的纹饰特征,并根据花粉形态对铁杉属进行分类,但其研究的多数物种被认为是同一种或某变种,且未建立铁杉属演化关系。铁杉属花粉形态与其授粉方式密切相关,可分为Sect. Hesperopeuce和Micropeuce(=Sect. Tsuga)两组[19](以下简称H组和M组),前者只包含山地铁杉1种,花粉有气囊,在珠孔处萌发形成花粉管;后者包括除山地铁杉外其他种,花粉无气囊,在珠鳞或苞鳞上萌发形成花粉管。还有学者根据花粉形态差异认为铁杉属应分为两属[32]。但是,该属花粉形态的研究还缺乏物种的全面取样,也很少在系统发育框架内探讨其形态演化。因此,有必要在全面取样基础上,开展铁杉花粉形态的研究,在该属系统发育框架下探究其分类意义。
本研究通过扫描电子显微镜观察现存铁杉属10种的花粉形态特征,结合3种铁杉属花粉化石标本信息,通过聚类分析,从花粉形态角度为铁杉属的分类提供了佐证,为该属化石标本分类地位确定提供参考。
1 材料与方法
1.1 材料采集
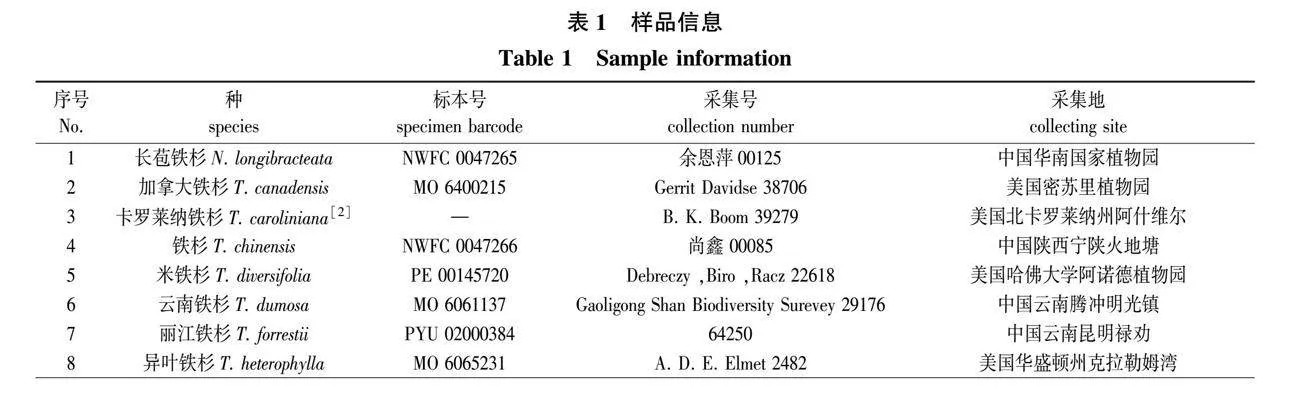
本研究共收集到10种铁杉属植物及近缘种长苞铁杉花粉。由于时间、地域限制,除铁杉外,其余花粉通过蜡叶标本上的雌球花获得,具体采样信息见表1。
铁杉花粉取样于秦岭火地塘,于2022年4月于火地塘实验林场铁杉混交林中采集铁杉的小孢子叶球,待其自然散粉后收集,保存于-20 ℃冰箱用作后续实验。新鲜花粉均采集自胸径大于30 cm的无明显病虫害的健康植株。
1.2 扫描电子显微镜观察
将带有花粉的雌球花样本与花粉样本,用导电双面胶粘在金属台上,25 mA真空喷金80 s两次后,使用日立S-3400N扫描电子显微镜(SEM)进行观察、拍照记录花粉形态特征。
1.3 聚类分析方法
1.3.1 形态特征数据集的建立
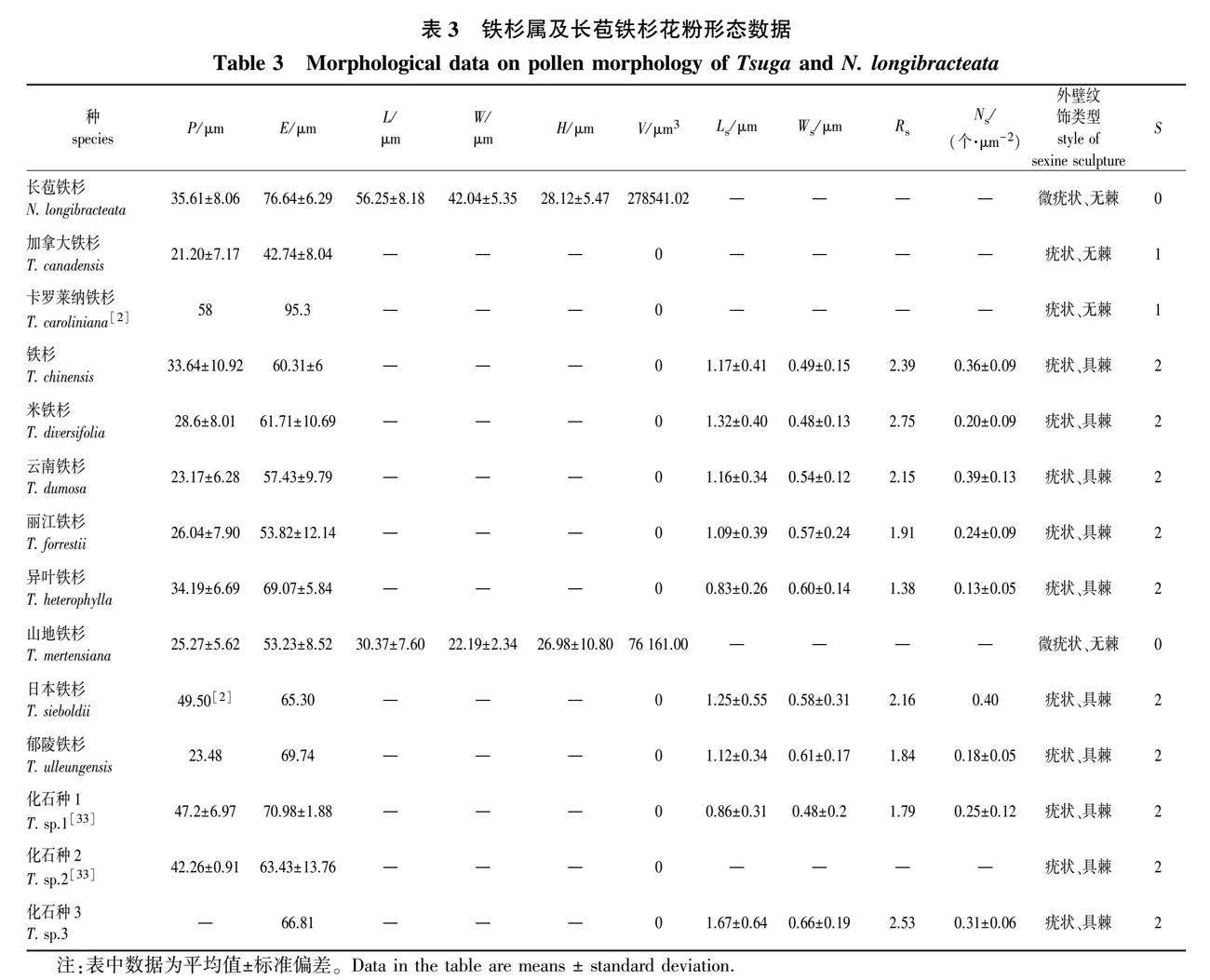
每种随机选取10粒花粉粒,利用Image J软件测其极轴长度(P)、赤道轴长度(E)、气囊长(L)、气囊宽(W)、棘突长(Ls)、棘突宽(Ws)、棘突密度(Ns)。对于部分铁杉属花粉化石的形态数据,通过查阅相关记录文献获得。计算各项指标的平均值±标准偏差。花粉形态学描述术语参考王伏雄等[23]、王开发和王宪曾[24]、埃尔特曼[34]。
1.3.2 性状的编码
采用等级数量编码方法进行编码[35],共选取11个性状(表2),其中数值性状(numerical character,简称“N”)8个,数值性状不编码,直接以原始数据形式进行运算;导出性状(induced character,简称“I”)2个,以数值性状原始数据按公式计算得到;无序多态性状(disordered multistate character,简称“D”)1个,采用综合评分法编码,取连续排列的正整数1、2、3[28]进行编码。
1.3.3 数据处理
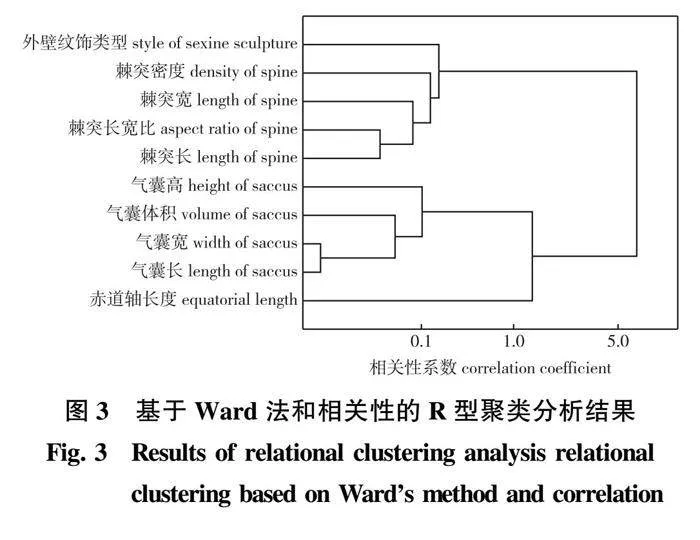
使用Origin 2022软件,对原始数据进行标准化(STD)处理以消除不同量纲,以10个性状为变量,种类为观测值,以各种的所属组(H组、M组)作为分组依据进行主成分分析。R型聚类分析选用Ward法,以相关性系数为度量距离生成性状指标聚类的树状图,选择用于Q型聚类的指标;Q型聚类分析选用Ward法,以欧氏距离为度量标准制作树状图。
2 结果与分析
2.1 花粉形态特征
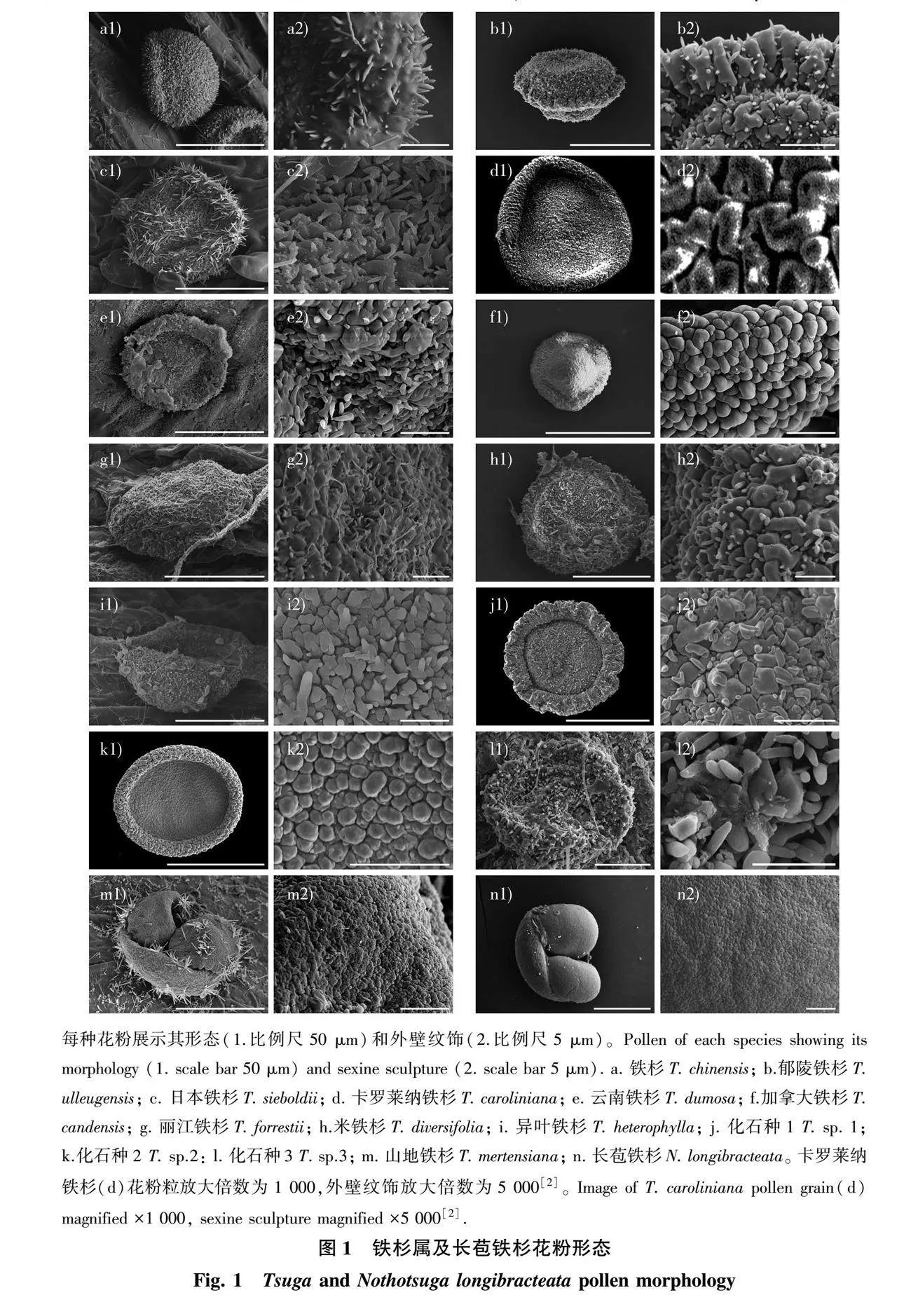
铁杉属花粉为N1P3C1型,远极面具有一薄壁区(图1),极轴长21.20~58.00 μm,赤道轴长42.74~95.30 μm,多无气囊,稀有气囊;花粉粒表面多疣状,稀微疣状;表面有棘突或无,棘突长0.83~1.67 μm,宽0.48~0.66 μm,长宽比值为1.38~2.75,有0.13~0.40个/μm2棘突(表3)。
长苞铁杉和山地铁杉花粉具气囊,其余各铁杉均无气囊(图1)。近缘种长苞铁杉与山地铁杉气囊大小差异明显,约为山地铁杉的3.66倍。加拿大铁杉赤道轴最短,极轴也最短,卡罗莱纳铁杉赤道轴最长,极轴最长。日本铁杉、卡罗莱纳铁杉、化石种1和化石种2极轴较长,在40 μm以上。长苞铁杉、异叶铁杉和化石种1赤道轴较长,在70 μm左右。
长苞铁杉花粉表面微疣状,较光滑;铁杉属花粉表面均具疣,山地铁杉表面较光滑,微疣状;加拿大铁杉、卡罗莱纳铁杉表面较粗糙,呈疣状;其余7种铁杉花粉外壁粗糙,表面有棘突,棘突长短、个数不尽相同。铁杉、米铁杉、云南铁杉、日本铁杉棘突较狭长,棘突长是棘突宽2倍以上;丽江铁杉、郁陵铁杉棘突稍短,棘突长接近棘突宽的2倍,其中异叶铁杉棘突最为短粗,棘突长宽比最小,仅为1.38。异叶铁杉花粉表面棘突个数最少,约0.13个/μm2;日本铁杉最多,为0.40个/μm2。此外,铁杉、云南铁杉和化石种3棘突也在0.30个/μm2以上,郁陵铁杉则在0.20个/μm2以下。
每种花粉展示其形态(1,比例尺50 μm)和表面纹饰(2,比例尺5 μm),A. T. chinensis; B: T. ulleugensis; C: T. sieboldii; D: T. caroliniana; E: T. dumosa; F: T. candensis; G: T. forrestii; H: T. diversifolia; I: T. heterophylla; J: T. sp. 1; K: T.sp.2: L: T. sp.3; M: T. mertensiana; N: N. longibracteata。 T. caroliniana(D)花粉粒放大倍数为1000倍,表面纹饰放大倍数为5000倍[2]。
Each pollen species showing its morphology (left,1, scale bar 25 μm) and sculpture (right,2,scale bar 5 μm), Image of T. caroliniana pollen grain(D) magnified ×1000, sculpture magnified ×5000[2]. A: T. chinensis; B: T. ulleugensis; C: T. sieboldii; D: T. caroliniana; E: T. dumosa; F: T. candensis; G: T. forrestii; H: T. diversifolia; I: T. heterophylla; J: T. sp. 1; K: T.sp.2: L: T. sp.3; M: T. mertensiana; N: N. longibracteata.
每种花粉展示其形态(1.比例尺50 μm)和外壁纹饰(2.比例尺5 μm)。Pollen of each species showing its morphology (1. scale bar 50 μm) and sexine sculpture (2. scale bar 5 μm). a. 铁杉T. chinensis; b.郁陵铁杉T. ulleugensis; c. 日本铁杉T. sieboldii; d. 卡罗莱纳铁杉T. caroliniana; e. 云南铁杉T. dumosa; f.加拿大铁杉T. candensis; g. 丽江铁杉T. forrestii; h.米铁杉T. diversifolia; i. 异叶铁杉T. heterophylla; j. 化石种1 T. sp. 1; k.化石种2 T. sp.2: l. 化石种3 T. sp.3; m. 山地铁杉T. mertensiana; n. 长苞铁杉N. longibracteata。 卡罗莱纳铁杉(d)花粉粒放大倍数为1 000,外壁纹饰放大倍数为5 000[2]。 Image of T. caroliniana pollen grain(d) magnified ×1 000, sexine sculpture magnified ×5 000[2].
2.2 主成分分析和聚类分析
2.2.1 主成分分析
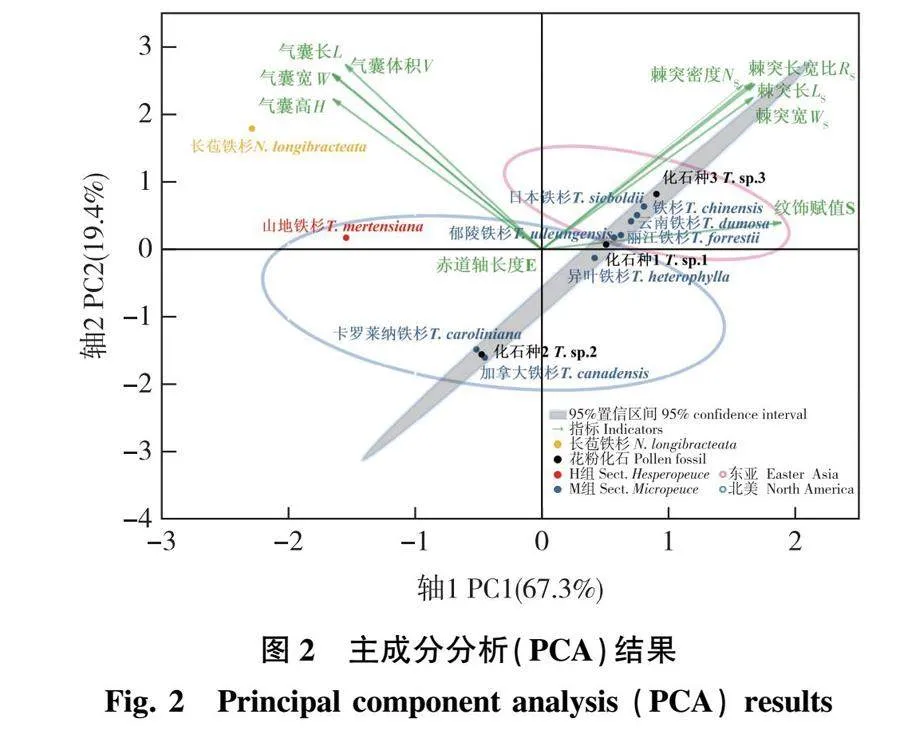
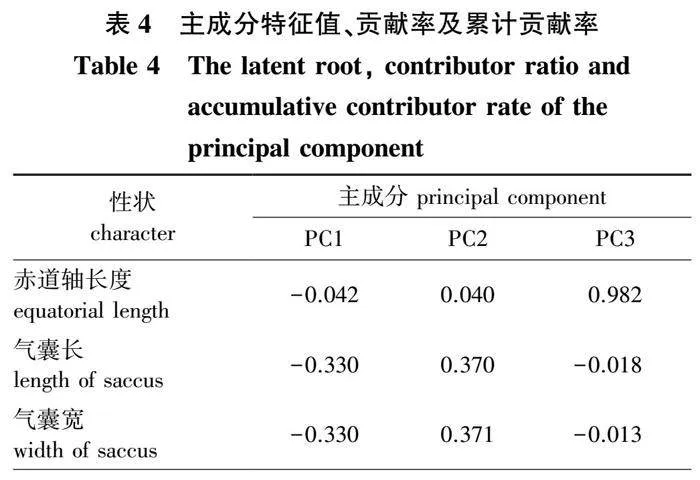
主成分分析贡献率结果(表4)显示,第1主成分的贡献率为67.259%,其中最显著的变量是纹饰赋值、棘突长宽比、棘突宽;第2主成分的贡献率为19.415%,其中最显著的变量是气囊体积、气囊宽、气囊长。前2个主要成分的总贡献率为86.674%,大于85%且两个主成分特征值均大于1,说明所选取的指标能较好解释样本的差异性,可以用于下一步分析。
主成分分析结果(图2)表明,M组与H组显著分离,M组主要集中在第1象限,与棘突有关指标距离较近;H组与长苞铁杉则在第3象限,与气囊指标关系较大。加拿大铁杉与卡罗莱纳铁杉与M组其他铁杉相距较远,处于第4象限(图2)。
化石种1与异叶铁杉、郁陵铁杉相似,化石种2则与卡罗莱纳铁杉、加拿大铁杉更加相似,化石种3与云南铁杉、日本铁杉和铁杉距离较近。此外,主成分分析结果将铁杉属按不同分布区分离开,东亚分布的各类群明显聚集在一起,北美分布各种较分散,但仍与东亚各种分离。
2.2.2 R型聚类分析
R型聚类结果(图3)表明,棘突长与棘突长宽比相关度较高,气囊长、气囊宽相关度较高,本试验将气囊长与棘突长宽比保留,用于Q型聚类分析。其他指标较分散,基本可以用于下一步Q型聚类分析。
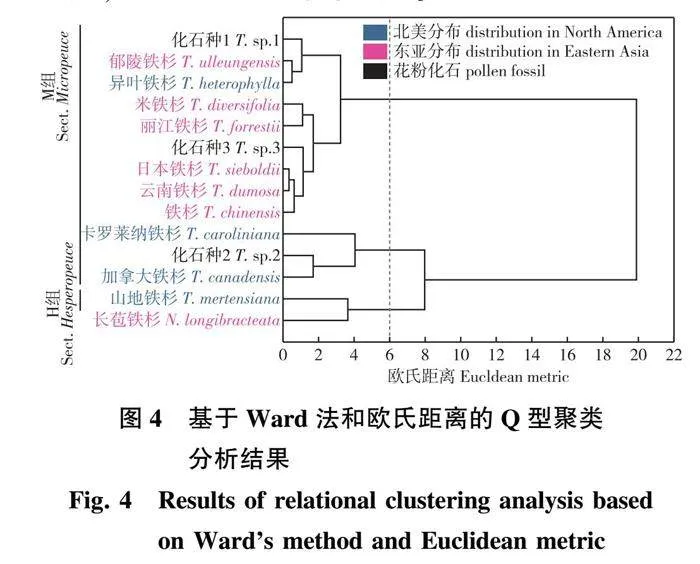
2.2.3 Q型聚类分析
由Q型聚类分析结果(图4)显示,欧氏距离为12时,分为两大类:①无棘突,包括长苞铁杉、山地铁杉、卡罗莱纳铁杉、加拿大铁杉;②有棘突,包括铁杉、云南铁杉、日本铁杉、丽江铁杉、米铁杉、异叶铁杉和郁陵铁杉。化石种1与化石种3属于第①大类,化石种2属于第②大类。
欧氏距离为6时,分为3类:①有气囊、无棘突,包括山地铁杉、长苞铁杉;②无气囊、无棘突,包括卡罗莱纳铁杉、加拿大铁杉、化石种2;③无气囊、无棘突,仍包括铁杉、云南铁杉、日本铁杉、丽江铁杉、米铁杉、异叶铁杉和郁陵铁杉。
欧氏距离为2时,共分为6类:①长苞铁杉;②山地铁杉;③加拿大铁杉、化石种2;④卡罗莱纳铁杉;⑤铁杉、云南铁杉、日本铁杉、丽江铁杉、米铁杉;⑥异叶铁杉和郁陵铁杉。
综上,本研究认为根据棘突与气囊的有无,将铁杉属分为3类(欧氏距离为6)最为合适,可以较好地判断和区分铁杉属内各种花粉。
根据聚类分析结果(图4),化石种1与郁陵铁杉、异叶铁杉相似,化石种2花粉与加拿大铁杉最为相似,化石种3在花粉形态上与日本铁杉、云南铁杉和铁杉组为姐妹群。
将Q型聚类分析结果与铁杉属分子系统发育树[14]比较(图5)发现,两者具有一定的相似性。长苞铁杉、山地铁杉、加拿大铁杉与卡罗莱纳铁杉都位于基部位置,云南铁杉、日本铁杉、丽江铁杉与铁杉关系较近。但形态学聚类分析将长苞铁杉与山地铁杉支系、卡罗莱纳铁杉与加拿大铁杉支系视为姊妹群,而系统发育树则认为它们是嵌套结构。此外,形态学分析认为异叶铁杉与郁陵铁杉较为相似,分子系统树支持异叶铁杉与山地铁杉亲缘关系较近。
3 讨 论
3.1 花粉形态特征的分类学及生物学意义
铁杉属植物花粉形态呈现出较为丰富的多样性。Q型聚类分析结果表明,气囊的有无与棘突特征是区分铁杉属花粉的重要依据。欧氏距离为6时,可据此将铁杉属花粉分为3类:①有气囊、无棘突类,为长苞铁杉与山地铁杉;②无气囊、无棘突类,为加拿大铁杉、卡罗莱纳铁杉和化石种2;③无气囊、有棘突类,为铁杉、云南铁杉、日本铁杉、丽江铁杉、米铁杉、异叶铁杉和郁陵铁杉和化石种1、3。邹惠渝等[36]将铁杉属划分为3个组:铁杉组(Sect. 1 Tsuga)、北美铁杉组(Sect. 2 Canadensis)和大果铁杉组(Sect. 3 Hesperopeuce),将卡罗莱纳铁杉、加拿大铁杉与异叶铁杉从铁杉组中分离为北美铁杉组Sect. 2 Canadensis,并将长苞铁杉独立成属。李楠[1]则将长苞铁杉纳入铁杉属,分为长苞铁杉组(Sect. Heopeuce)、铁杉组(Sect. Tsuga)和大果铁杉组(Sect. Hesperopeuce),本研究依据花粉形态所得的分类结果更为支持邹惠渝等[36]的分组方案,但不同意其将长苞铁杉独立成属的观点。
花粉特征在一定程度上可以表现植物演化的一般规律[37]。目前,一般认为花粉形态的演化顺序是气囊从有到无、体积由大到小、外壁纹饰由光滑到短棘突再到长棘突[38]。分子与形态系统发育树中,有气囊的长苞铁杉和山地铁杉位于最基部,无棘突的加拿大铁杉和卡罗莱纳铁杉也位于发育树底端,而各类无气囊、有棘突的类群则位于发育树顶端。说明铁杉属中有气囊、无棘突为原始类型,无气囊、有棘突则为进化类型,铁杉属花粉的演化趋势为有气囊至无气囊、无棘突至有棘突。
铁杉属植物花粉形态的演化过程与其授粉方式密切相关。在有气囊花粉类群中,植株直立,依靠授粉滴捕获花粉,花粉无需被其他部位捕获。而在无气囊花粉类群中,花粉不能被授粉滴捕获,而是利用棘突附着于苞鳞背面,因此,棘突的数量和形态有利于花粉稳定附着于苞鳞背面。相较于有气囊花粉类群,棘突的出现无疑增加了铁杉属植物的受精成功率和生态适应性,极大保障了铁杉属植物的繁殖优势。
3.2 花粉形态特征的亲缘关系分析
花粉形态的稳定性在一定程度上可以表现种属间的亲缘关系[23-24]。根据聚类分析结果显示,山地铁杉与长苞铁杉互为姊妹群,卡罗莱纳铁杉则与加拿大铁杉互为姐妹群,异叶铁杉与郁陵铁杉亲缘关系最近,铁杉则与云南铁杉关系最紧密。Havill等[13]与Feng等 [14]建立的分子发育树中,卡罗莱纳铁杉与加拿大铁杉嵌套进入了亚洲支系中,本研究则发现两者花粉形态与北美支系关系更紧密,与地理位置相对应。此外,本研究建立的铁杉属花粉形态发育树,其东亚分布的种类虽然聚类到了1个分支,但拓扑结构与分子系统发育树存在一定差异,这可能是东亚分布的铁杉属植物经历了多次杂交和演化,具有复杂的网状演化模式,为亲缘关系的界定带来较大的困难[14]。
长苞铁杉是中国特有种,自发表以来,其分类地位具有颇多争议。郑万钧先生[10]发现长苞铁杉苞鳞露出种鳞之外,不同于其他铁杉属植物,将其列为新种并和北美分布的山地铁杉纳入大果铁杉组中;Flous[39]发现长苞铁杉叶上有散生斑点,薄壁细胞中有石细胞,与其他铁杉属植物明显不同,应独立成组。胡先骕先生[40]首次提出建立长苞铁杉属(Nothotsuga),得到了Page[41]的支持。但又有学者通过染色体核型[42-45]、木材结构[46-47]方面的研究,认为应当将长苞铁杉纳入铁杉属长苞铁杉组。近来分子系统学证据表明,长苞铁杉区别于铁杉属,应独立为长苞铁杉属[11]。本研究中发现,长苞铁杉花粉与山地铁杉花粉形态相近,只在花粉和气囊体积上略有差异,以花粉形态为分类标准,长苞铁杉并不能脱离铁杉属单独成属,支持将其纳入铁杉属的观点。
铁杉属花粉特征与其分布地有密切的关系。主成分分析与聚类分析结果中,北美分布的种类(异叶铁杉除外)与东亚分布的各种明显分离,说明同一地区分布的花粉存在一定的相似性。北美分布的铁杉属植物表面大多较光滑,而东亚分布种类表面均粗糙,附着有长短粗细不一的棘突。值得注意的是,北美分布的异叶铁杉具有棘突,与东亚分布郁陵铁杉亲缘关系最近,而与北美分布的其他种类亲缘关系较远。分子系统学研究支持异叶铁杉是同样分布于北美的山地铁杉的姊妹群[13-14],二者位于铁杉属的基部位置。花粉形态聚类只强调了不同种类花粉形态上的相似性,无法从遗传物质角度判断植物间的亲缘关系,仍具有一定的局限性。此外,分布于韩国郁陵岛的郁陵铁杉在2017年才被发现,其花粉形态与北美分布的异叶铁杉相似,可能与铁杉属的起源与扩散有关:始新世到中新世时期,白令陆桥是北美洲与亚洲进行物种交流的重要走廊,两个地区的植物可能通过此通道进行了交流[48]。
在铁杉属花粉形态研究基础上,笔者尝试了对化石花粉的鉴定。化石种1花粉无气囊、外壁有棘突,聚类分析中其处于异叶铁杉和郁陵铁杉支系的基部,因为花粉来源于奥地利中新世地层[33],推测其可能为异叶铁杉和郁陵铁杉祖先类型曾经在欧洲分布的遗迹[48]。化石种2形态上与加拿大铁杉、卡罗莱纳铁杉相近,均无气囊、外壁纹饰疣状,根据聚类分析结果,鉴定化石种2为加拿大铁杉花粉。聚类分析结果将化石种3嵌套在包含铁杉、云南铁杉和日本铁杉支系的基部位置,推测该化石与该分支其他种的祖先具有密切关系。
综上所述,根据花粉形态特征对铁杉属进行分类和亲缘关系探究具有一定的科学性与准确性;尤其是依据花粉气囊的有无与外壁纹饰特征,可以较好地对大多数铁杉属花粉进行分类,对铁杉属花粉的鉴定与亲缘关系的探究也具有积极意义。
致谢:
西北农林科技大学重点实验室的张国云老师、园艺学院陈蕊红老师提供扫描电镜,兰州大学孙柏年教授和韩磊博士提供化石样本,中科院植物所的孔宏智研究员和于宁宁博士、华南植物园的余恩平同学、云南大学的周新茂副教授及蒋璐遥同学、南京林业大学的袁禹婷同学、美国密苏里植物园的张丽兵研究员和韩国国家植物园Chang Kae Sun教授协助样品采集。
参考文献(reference):
[1]李楠. 论松科植物的地理分布、起源和扩散[J]. 植物分类学报, 1995, 33(2): 105-130. LI N. Studies on the geographic distribution, origin and dispersal of the family Pinaceae Lindl.[J]. Acta Phytotaxon Sin, 1995, 33(2): 105-130.
[2]SIVAK J. Observations nouvelles sur les grains depollen de Tsuga[J]. Pollen et Spores, 1973, 15(3-4): 397-457.
[3]WU M X, HUANG J, SU T, et al.Tsuga seed cones from the late Paleogene of southwestern China and their biogeographical and paleoenvironmental implications[J]. Palaeoworld, 2020, 29(3): 617-628. DOI: 10.1016/j.palwor.2019.07.005.
[4]GAUSSEN H. Les Gymnospermes actuelles etfossiles.[M]. Toulouse: Travaux du Laboratoire Forestier de Toulouse, 1967.
[5]LEPAGE B A.The evolution,biogeography and palaeoecology of the Pinaceae based on fossil and extant representatives[J].Acta Hortic,2003(615):29-52.DOI: 10.17660/actahortic.2003.615.1.
[6]HOLMAN G,DEL TREDICI P,HAVILL N,et al.A new species and introgression in Eastern Asian hemlocks (Pinaceae:Tsuga)[J].Syst Bot,2017,42(4):733-746.DOI: 10.1600/036364417x696474.
[7]FARJON A. Pinaceae. drawing and description of the generaAbise, Cedrus, Pseudolarix, Keteleeria, Nothotsuga, Tsuga, Cathaya, Pseudotsuga, larix and Picea[M]. Knigstein: Koeltz Scientific Books, 1990.
[8]FARJON A. A handbook of the world’s conifers[M]. Leiden: Brill, 2010.
[9]FU LG, LI Y, MILL RR. Flora of China (Vol.4)[M]. Beijing: Science Press amp; St. Louis: Missouri Botanical Garden Press, 1999.
[10]郑万钧. 贵州铁杉之一新种[J]. 中国科学社生物研究所论文集:植物辑, 1932, 7(1): 1-2. CHENG W C. A new Tsuga from southwestern China [J]. Contributions from the Biological Laboratory of the Science Society of China: Botanical Series, 1932, 7(1): 1-2.
[11]RAN J H, SHEN T T, WU H, et al. Phylogeny and evolutionary history of Pinaceae updated by transcriptomic analysis[J]. Mol Phylogenet Evol, 2018, 129: 106-116. DOI: 10.1016/j.ympev.2018.08.011.
[12]LI M, BADGER J H, CHEN X, et al.An information-based sequence distance and its application to whole mitochondrial genome phylogeny[J]. Bioinformatics, 2001, 17(2):149-154. DOI: 10.1093/bioinformatics/17.2.149.
[13]HAVILL N P, CAMPBELL C S, VINING T F, et al. Phylogeny and biogeography of Tsuga (Pinaceae) inferred from nuclear ribosomal ITS and chloroplast DNA sequence data[J]. Syst Bot, 2008, 33(3): 478-489. DOI: 10.1600/036364408785679770.
[14]FENG Y Y, SHEN T T, SHAO C C, et al. Phylotranscriptomics reveals the complex evolutionary and biogeographic history of the genus Tsuga with an East Asian-North American disjunct distribution[J]. Mol Phylogenet Evol, 2021, 157: 107066. DOI: 10.1016/j.ympev.2020.107066.
[15]CHEN L,LIU X,WANG Z B, et al. Comparative chloroplast genome analyses of six hemlock trees in East Asia: insights into their genomic characterization and phylogenetic relationship[J]. Forests, 2023, 14(11): 2136. DOI: 10.3390/f14112136.
[16]LEPAGE B A. A new species of Tsuga (Pinaceae) from the middle Eocene of Axel Heiberg Island, Canada, and an assessment of the evolution and biogeographical history of the genus[J]. Bot J Linn Soc, 2003, 141(3): 257-296. DOI: 10.1046/j.1095-8339.2003.00131.x.
[17]OWENS J N,TAKASO T,RUNIONS C J. Pollination in conifers[J]. Trends Plant Sci,1998, 3(12): 479-485. DOI: 10.1016/s1360-1385(98)01337-5.
[18]TAYLOR R J. The relationship and origin of Tsuga heterophylla and Tsuga mertensiana based on phytochemical and morphological interpretations[J]. Am J Bot, 1972, 59(2): 149. DOI: 10.2307/2441394.
[19]OWENS J N, BLAKE M D. Pollen morphology and development of the pollination mechanism in Tsuga heterophylla and T. mertensiana[J]. Can J Bot, 1983,61(12): 3041-3048. DOI: 10.1139/b83-341.
[20]FERNANDO D D, QUINN C R, BRENNER E D, et al. Male gametophyte development and evolution in extant gymnosperms[J]. Int J Dev Biol, 2010, 4(Spec.1): 47-63. DOI: 10.3969/j.issn.1000-6281.2011.03.018.
[21]DING S T, WU J Y, TANG D L, et al. Seed cones of Tsuga (Pinaceae) from the upper Miocene of Eastern China:biogeographic and paleoclimatic implications[J].Rev Palaeobot Palynol, 2021, 285: 104358. DOI: 10.1016/j.revpalbo.2020.104358.
[22]FOWELLS H. Silvics of forest trees of the United States[M]. Washington, DC: Us Department of Agriculture, Forest Service, 1965.
[23]王伏雄, 钱南芬, 张玉龙, 等. 中国植物花粉形态[M]. 2版. 北京: 科学出版社, 1995: 1-10, 197-202. WANG F X. Pollen flora of China[M]. 2nd ed. Beijing:Science Press, 1995: 1-10, 197-202.
[24]王开发, 王宪曾. 孢粉学概论[M]. 北京:北京大学出版社,1983:23,32-34. WANG K F, WANG X Z. Introduction to sporology[M]. Beijing: Peking University Press, 1983: 23, 32-34.
[25]LIN M S, QIU J, XIE K Q, et al. Palynological features and taxonomic significance for 16 species of Gagea (Liliaceae) from Xinjiang, China[J]. PhytoKeys, 2023, 225: 53-68. DOI: 10.3897/phytokeys.225.101518.
[26]薛美玲, 郝秀东, 黄旭光,等. 20个朱槿品种花粉形态扫描电镜观察分析[J]. 分子植物育种, 2023, 1-17. XUE M L, HAO X D, HUANG X G, et al. Morphology analysis of pollen grains of 20 species of Hibiscus using scanning electron microscopy[J]. Molecular Plant Breeding, 2023, 1-17.
[27]张淑钧, 杨欣欣, 罗建. 西藏色季拉山区风毛菊属植物花粉形态特征及其分类学意义[J]. 植物研究, 2023, 43(5): 741-755. ZHANG S J, YANG X X, LUO J. Micromorphological characteristics of pollen and its taxonomic significance of Saussurea in Sedgera Mountains of Tibet[J]. Bull Bot Res, 2023, 43(5): 741-755. DOI: 10.7525/j.issn.1673-5102.2023.05.011.
[28]曹婧, 戴忠良, 徐迎春, 等. 睡莲属植物花粉形态研究[J]. 西北植物学报, 2023, 43(7): 1227-1235. CAO J, DAI Z L,XU Y C,et al.Studies on pollen morphology of Nymphaea[J].Acta Bot Boreali Occidentalia Sin,2023,43(7):1227-1235.DOI: 10.7606/j.issn.1000-4025.2023.07.1227.
[29]孙爱芝,马玉贞,黄昌庆,等.松科花粉鉴定要点及其生态环境特征研究[J].兰州大学学报,2006,42(5):18-21.SUN A Z,MA Y Z,HUANG C Q,et al.Study of the pollen morphologic appraisable point of pinaceaeous main genera and their ecological environment[J].J Lanzhou Univ,2006,42(5):18-21.DOI: 10.13885/j.issn.0455-2059.2006.05.004.
[30]张金谈.中国松科花粉形态研究[J].植物研究,1989,9(3):87-95,97-98.ZHANG J T.Studies on the pollen morphology of Pinaceae in China[J].Bull Bot Res,1989,9(3):87-95,97-98.
[31]王伏雄, 钱南芬, 张金谈. 花粉形态的研究 Ⅱ.松科及落羽杉科的花粉[J].植物学报, 1955, 4(1): 47-62. WANG F X, QIAN N F, ZHANG J T. Studies on pollen morphologyⅡ. Pollen of Pinaceae and Taxodaceae[J]. Journal of Integrative Plant Biology, 1955, 4(1): 47-62.
[32]HO R,SZIKLAI O.On the pollen morphology of Picea and Tsuga species[J].Grana,1972,12(1):31-40.DOI: 10.1080/00173137209427643.
[33]GRMSSON F,ZETTER R.Combined LM and SEM study of the Middle Miocene (Sarmatian) Palynoflora from the Lavanttal Basin,Austria:part II.Pinophyta (Cupressaceae,Pinaceae and Sciadopityaceae)[J].Grana,2011,50(4):262-310.DOI: 10.1080/00173134.2011.641450.
[34](瑞典)埃尔特曼(G.ERDFMAN). 孢粉学手册[M].中国科学院植物研究所古生物研究室孢粉组, 译.北京:科学出版社,1978. ERDFMAN G. Handbook of palynology[M]. Spore Pollen Group, Paleontological Research Laboratory, Institute of Botany, Chinese Academy of Sciences, Translation. Beijing: Science Press,1978.
[35]徐克学.数量分类学[M].北京:科学出版社,1994:27-28,47-50.XU K X.Numerical taxonomy[M].Beijing:Science Press,1994:27-28,47-50.
[36]邹惠渝,张凤春.长苞铁杉和铁杉属数量分类的研究[J].南京林业大学学报(自然科学版),1996,20(1):43-47.ZOU H Y,ZHANG F C.A study on numerical taxonomy of longbracted hemlock[J].J Nanjing For Univ,1996,20(1):43-47.
[37]CALIC D,DEVRNJA N,KOSTIC I,et al.Pollen morphology,viability,and germination of Prunus domestica cv.Pozegaca[J].Sci Hortic,2013,155:118-122.DOI: 10.1016/j.scienta.2013.03.017.
[38]WODEHOUSE" R P. Pollen grains[M]. New York: McGraw-Hill Book Company, 1935: 323-340.
[39]FLOUS F.Révision du genre Tsuga[M]. Toulouse: Faculté des sciences-Laboratoire forestier, 1936.
[40]胡先骕.种子植物分类学讲义[M].上海:中华书局,1951.HU X S.Lecture notes on seed plant taxonomy [M].Shanghai: Zhonghua Book Company,1951: 64.
[41]PAGE C. New and maintained genera in the conifer families Podocarpaceae and Pinaceae[J]. Notes Roy. Bot. Gard. Edinburgh, 1988, 45(2):377-395.
[42]李林初.若干铁杉属植物核型的比较研究[J].广西植物,1988,8(4):324-328.LI L C.The comparative karyotypic studies in some species of Tsuga (Pinaceae) [J].Guihaia,1988,8(4):324-328.
[43]李林初.长苞铁杉的核型分析及其分类学意义[J].云南植物研究,1991,13(3):309-313.LI L C.The karyotype analysis of Tsuga longibracteata and its taxonomic significance[J].Acta Bot Yunnanica,1991,13(3):309-313.
[44]李林初.松科的核型和系统发育研究[J].植物分类学报,1995,33(5):417-432.LI L C.Studies on the karyotype and phylogeny of the Pinaceae[J].Acta Phytotaxon Sin,1995,33(5):417-432.
[45]李林初,杨凤辉,蔡星星.大果铁杉的核型分析及铁杉属的细胞分类学研究[J].复旦学报(自然科学版),2000,39(4):432-435.LI L C,YANG F H,CAI X X.Karyotype analysis of Tsuga mertensiana and cytotaxonomic study on Tsuga (Pinaceae)[J].J Fudan Univ,2000,39(4):432-435.DOI: 10.15943/j.cnki.fdxb-jns.2000.04.017.
[46]喻诚鸿. 铁油杉的木材结构和它在分类上的位置[J]. 植物学报, 1956, 5(2): 243-248. YU C H. The wood structure of Tsuga-Keteleeria longibracteata special reference to its taxonomy[J]. Journal of Integrative Plant Biology, 1956, 5(2): 243-248.
[47]周崟,姜笑梅.木材构造特征在裸子植物系统学中的意义[J].植物分类学报,1992, 30(5):405-414.ZHOU Y, JIANG X M. Characteristics of wood structure in gymnosperms and their systematic significance[J]. Acta Phytotaxon Sin, 1992, 30(5): 405-414.
[48]肖良,王宏宇,国丽妍,等.铁杉属的起源、扩散与传播:基于化石证据[J].地球环境学报,2023,14(1):9-26.XIAO L,WANG H Y,GUO L Y,et al.Origin and spread of Tsuga (Endl.) Carrière based on fossil records[J].J Earth Environ,2023,14(1):9-26.
(责任编辑 吴祝华)
收稿日期Received:2024-06-17""" 修回日期Accepted:2024-07-10
基金项目:国家植物标本资源库项目(E0117G1001)。
第一作者:陈芷涵(13657974778@163.com)。
*通信作者:张鑫(xin.zhang@nwafu.edu.cn),讲师,博士。
引文格式:陈芷涵,尚鑫, 谢淦,等.
铁杉属花粉形态及其聚类分析研究[J]. 南京林业大学学报(自然科学版),2024,48(4):37-45.
CHEN Z H, SHANG X, XIE Z, et al.
Phylogenetic relationship and evolutionary patterns of Tsuga pollen morphology: a cluster analysis-based study[J]. Journal of Nanjing Forestry University (Natural Sciences Edition),2024,48(4):37-45.
DOI:10.12302/j.issn.1000-2006.202406025.
猜你喜欢
中国棉花(2023年9期)2023-12-17 02:08:05
智富时代(2018年7期)2018-09-03 03:47:26
电线电缆(2017年4期)2017-07-25 07:49:48
商情(2016年39期)2016-11-21 08:45:54
新媒体研究(2016年19期)2016-11-18 19:48:34
大经贸(2016年9期)2016-11-16 16:16:46
中国市场(2016年33期)2016-10-18 12:16:58
科技视界(2016年20期)2016-09-29 12:32:48
企业导报(2016年9期)2016-05-26 21:08:14
岩矿测试(2015年3期)2015-12-21 03:57:04
