
基于iTRAQ蛋白质组学的OsAAP6不同转基因水稻胚乳差异蛋白筛选
2024-08-22 00:00:00彭波孙晓宇邱静彭娟赵强孙艳芳宋晓华张志国周伟汪全秀
南方农业学报 2024年4期



摘要:【目的】基于iTRAQ蛋白质组学筛选不同OsAAP6转基因水稻胚乳的差异蛋白,明确胚乳中的差异蛋白,以期为后续利用OsAAP6基因提升稻米的营养品质提供理论依据。【方法】以水稻OsAAP6超量表达阳性[OX(+)]和阴性 对照[OX(-)]材料及OsAAP6互补表达阳性[ZpZc(+)]和阴性对照[ZpZc(-)]为试验材料,采用同位素标记相对与绝对定量(iTRAQ)技术对其进行蛋白质组学检测分析,并利用考马斯亮蓝G-250法对其胚乳中的4类储藏蛋白(谷蛋白、醇溶蛋白、清蛋白和球蛋白)进行含量检测。【结果】与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性胚乳中OsAAP6基因的表达量分别在Plt;0.01和Plt;0.001水平极显著升高。与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性水稻共有58种差异蛋白共同上调或下调表达,其中有39种蛋白显著增加(Plt;0.05,下同),有19种蛋白显著降低。在39种共同上调表达的蛋白中有27种蛋白参与水稻种子储藏底物的合成与积累,且有16种上调表达蛋白参与种子储藏蛋白的代谢和积累。与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性水稻胚乳中的谷蛋白、醇溶蛋白、清蛋白和球蛋白含量均显著增加。【结论】OsAAP6基 因上调表达,能抑制与支链淀粉合成相关蛋白的积累,但能促进其胚乳中储藏蛋白的积累,进而提高胚乳中谷蛋白、 醇溶蛋白、清蛋白和球蛋白含量,可用于高蛋白水稻新品种的分子育种和遗传改良。
关键词:水稻;iTRAQ;蛋白质组学;OsAAP6基因;差异蛋白;营养品质
中图分类号:S511.035.3
文章编号:2095-1191(2024)04-1060-10
文献标志码:A
Screening differential proteins in OsAAP6 different transgenic rice endosperms based on iTRAQ proteomics
PENG Bo1, SUN Xiao-yu1, QIU Jing1, PENG Juan2, ZHAO Qiang3, SUN Yan-fang1, SONG Xiao-hua4, ZHANG Zhi-guo5, ZHOU Wei1, WANG Quan-xiu1*
(1College of Life Sciences, Xinyang Normal University/Dabie Mountain Agricultural Biological Resource Protection and Utilization Research Institute, Xinyang, Henan 464000, China; 2Xinyang Plant Protection and Inspection Station, Xinyang, Henan 464000, China; 3Henan Scientific Research Platform Service Center, Zhengzhou, Henan 450003, China;4Xinyang Academy of Agricultural Sciences, Xinyang, Henan 464000, China; 5Henan Lingrui PharmaceuticalCompany Limited, Xinyang, Henan 465550, China)
Abstract:【Objective】The purpose of the study was to screen the differential proteins in the endosperm of different OsAAP6 transgenic rice based on iTRAQ proteomics, and to clarify the differential proteins in the endosperm, in a bid to provide a theoretical basis for the future use of the OsAAP6 gene to enhance the nutritional quality of rice. 【Method】Using rice OsAAP6 overexpression positive [OX(+)] and negative control [OX(-)] materials, as well as OsAAP6 complemen-tary expression positive [ZpZc (+)] and negative control [ZpZc(-)] experimental materials, the isotope marker relative and absolute quantification (iTRAQ) technology was used for proteomic detection and analysis. And the contents of four types of storage proteins (gluten, gliadin, albumin and globulin) in its endosperm was detected using the Coomassie brilliant blue G-250 method. 【Result]Compared with the ZpZc (-) and OX(-) transgenic negative control, the expression of OsAAP6 gene in ZpZc(+) and OX(+) transgenic positive endosperm was extremely significantly increased at Plt;0.01 and Plt;0.001 levels, respectively. Compared with the ZpZc(-) and OX(-) transgenic negative control, a total of 58 differen- tial proteins were simultaneously up-regulated or down-regulated for expression in ZpZc(+) and OX(+) transgenic posi- tive rice, of which 39 proteins significantly increased (Plt;0.05, the same below), and 19 proteins significantly decreased.Among the 39 co-up-regulated proteins, 27 proteins were involved in the synthesis and accumulation of rice seed storage substrates, and 16 up-regulated proteins were involved in the metabolism and accumulation of seed storage proteins.Compared with the ZpZc(-) and OX(-) transgenic negative control, the content of glutelin, gliadin, albumin and globulin in the endosperm of ZpZc (+) and OX(+) transgenic positive rice was significantly increased. 【Conclusion】 Up- regulation of the OsAAP6 gene expression can inhibit the accumulation of proteins related to amylopectin synthesis, but promote the accumulation of storage proteins in the endosperm, thereby increasing the content of gluten, gliadin, albu- min and globulin in the endosperm. It can be used for molecular breeding and genetic improvement of new high protein rice varieties.
Key words: rice; iTRAQ; proteomics; OsAAP6 gene; differential proteins; nutritional quality
Foundation items: National Natural Science Foundation of China (U2004141) ; Henan Science and Technology Re-search Project (222102110141,222102110115,212102110249)
0 引言
【研究意义]水稻(Oryza sativa L.)是世界上最重要的粮食作物之一,我国有超过三分之二的人口以稻米为主食(Tian et al.,2009;彭波等,2023)。稻米富含优质蛋白,是人们能量及营养物质的重要来源(Peng et al.,2021),故改良并提升稻米营养品质对改善人类自身营养与健康具有重要作用。利用图位克隆策略,从水稻基因组中分离克隆到1个正向调控其种子蛋白质含量且对其稻米营养品质具有提升作用的氨基酸透性酶基因OsAAP6,其高效表达有利于提高水稻籽粒中总氨基酸含量(gt;6.0mg/g)、总必需氨基酸含量(gt;5.2mg/g)和蛋白质含量(>5.5mg/g),进而提升其稻米的营养品质(Peng et al.,2014;彭波等,2019)。但OsAAP6基因对不同储藏表达蛋白的积累及其分子调控机制尚不清楚。因此,针对水稻重要的品质性状基因OsAAP6,利用同位素标记相对和绝对定量(iTRAQ)技术和考马斯亮蓝G-250法对OsAAP6转基因稻米胚乳蛋白进行定量和定性检测,揭示OsAAP6转基因稻米中的差异蛋白,对全面阐明OsAAP6基因的生物学功能及开展高营养品质水稻新品种的分子育种具有重要意义和潜在应用价值。【前人研究进展】随着经济社会的持续发展,人们生活质量不断提升,对优质稻米的需求将会持续增加(罗同平等,2023;Sung et al.,2023)。稻米的品质性状主要包括营养品质、研磨加工品质、外观品质、安全品质、蒸煮与食味品质(Li et al.,2018;Peng etal.,2019;Bharali et al.,2022),其中稻米营养品质主要取决于水稻种子中的蛋白质含量。稻米中的蛋白质按溶解性主要分为四大类:溶于稀酸或稀碱的谷蛋白、溶于稀盐酸的球蛋白、溶于醇与水混合物的醇溶蛋白及溶于水的清蛋白(Shewry and Halford,2006;Zhou et al.,2017;兰艳等,2019),其中谷蛋白含量最高且最容易被人体消化吸收的蛋白(彭波等,2017;Du et al.,2022)。水稻种子中的蛋白质种类与含量不仅是决定其稻米营养品质的关键因素,还能影响稻米的其他品质性状,例如稻米的研磨加工品质、蒸煮与食味品质等(Chen et al.,2018;杨陶陶等,2023)。已有研究表明,水稻种子内的氨基酸组成较合理,且必需氨基酸含量相对较多(唐小涵等,2021;高渐飞等,2023)。因此,提高水稻种子内的蛋白质含量,能有效改善稻米的营养品质。水稻种子蛋白质含量属于一个典型的数量性状,其遗传基础与分子调控机制相对复杂,同时外界环境因素(如气候条件和营养物质等)对水稻种子蛋白质含量具有明显的影响作用(Yang et al.,2019;Long et al.,2023)。水稻中调控种子蛋白质含量的基因通常以基因家族的形式存在,其表达具有发育和组织的特异性(Li etal.,2022)。不论是种子中的谷蛋白还是醇溶蛋白,均由多个基因编码,如水稻种子中编码谷蛋白的基因家族至少有15个成员,主要分为四大类:A型、B型、C型和D型,故相应的谷蛋白也分为4个类型:A型谷蛋白(Glutelin type-A,GluA)、B型谷蛋白(Glute-lin type-B,GluB)、C型谷蛋白(Glutelin type-C,GluC)和D型谷蛋白(Glutelin type-D,GluD),其中A型谷蛋白包括GluA-1、GluA-2、GluA-3和GluA-4,B型谷蛋白包括GluB-1、GluB-2、GluB-3、GluB-4、GluB-5和GluB-6(Chen et al.,2018;Li et al.,2022)。水稻种子中的醇溶蛋白含量仅次于谷蛋白含量,根据分子量的不同分为3种类型,即10、13和16kD(Chen et al.,2018;Guo et al.,2022)。水稻种子中的球蛋白和清蛋白主要分布于糊粉层,且这2类储藏蛋白的含量均较低。从水稻中克隆获得编码种子中清蛋白(16kDα-淀粉酶/胰蛋白酶抑制剂)的基因RAG2(Zhou et al.,2017),其在水稻成熟种子中特异表达,超表达或抑制其表达分别显著增加或降低种子中蛋白质含量和总脂肪含量,同时超表达RAG2基因能显著增加水稻籽粒大小。iTRAQ是一种高通量的蛋白质组学技术,在水稻组织、逆境胁迫、亚细胞水平和激素诱导等方面研究均有应用(陈凌华等,2017;Ma et al.,2020),如采用iTRAQ技术检测水稻胚胎发育过程的蛋白质组变化,结果显示共有867个蛋白发生了显著性变化(Ziet al.,2013);采用iTRAQ技术对孕穗期、开花期和乳熟期的水稻颖壳进行蛋白质组学分析,结果发现有563个差异表达蛋白,主要参与叶绿素的合成途径(Wang et al.,2015);利用iTRAQ技术检测垩白突变体材料的蛋白组变化,结果发现113个与籽粒垩白形成相关的蛋白,这些差异表达蛋白主要参与碳水化合物代谢、蛋白合成与降解等过程(Lin et al.,2014)。因此,iTRAQ蛋白质组学方法具有检测范围广泛、精度较高、检测限低、分析结果可靠等优点,为解析水稻胚乳中不同蛋白含量及其4类储藏蛋白(谷蛋白、醇溶蛋白、球蛋白和清蛋白)含量之间的关系提供新技术支持。在水稻基因组中氨基酸转运蛋白基因家族有85个成员,分布于水稻的12条染色体上(Zhao et al.,2018),其中,氨基酸透性酶基因亚家族共有19个成员(OsAAPI~OsAAP19),参与调控水稻的产量及其品质等多种生物学性状,在植物的生长发育中发挥极其重要的功能。如OsAAP1基因通过增加中性氨基酸的摄取和再分配来改善水稻的生长状况(Jietal.,2020);敲除OsAAP3基因能增加水稻的分蘖数,提高水稻产量(Lu et al.,2018);OsAAP4基因通过调节中性氨基酸分配,提高水稻分蘖数和产量(Fanget al.,2021);抑制水稻OsAAP5基因的表达,能提高水稻产量(Wang et al.,2019);OsAAP10突变体种子的直链淀粉含量与食味值显著下降(Wang et al.,2020);OsAAP14基因是调控水稻分蘖与芽伸长的关键基因(Yang et al.,2022);OsAAP15是一种响应氮浓度的氨基酸转运蛋白,在极端氮浓度下有助于提高水稻产量(Yang et al.,2023)。【本研究切入点】本课题组前期研究发现,OsAAP6基因可影响水稻根对多种氨基酸的吸收及其在体内不同部位的分布,是水稻种子蛋白质含量及其营养品质的正向调节因子
(Peng et al.,2014;彭波等,2019),但OsAAP6基因是如何精细调控稻米胚乳中不同储藏表达蛋白积累、胚乳中不同蛋白含量及其4类储藏蛋白含量的相关性尚不清楚。【拟解决的关键问题】以水稻OsAAP6超量表达阳性[OX(+)]和阴性对照[OX(-)]材料及OsAAP6互补表达阳性[ZpZc(+)]和阴性对照[ZpZc(-)]试验材料,采用iTRAQ技术和考马斯亮蓝G-250法对转基因材料进行蛋白定量和定性分析,揭示OsAAP6转基因稻米胚乳中的差异蛋白,为后期利用OsAAP6基因提升稻米的营养品质提供理论参考。
1材料与方法
1.1试验材料
供试材料为本课题组前期(Peng et al.,2014;彭波等,2019)已获得水稻OsAAP6超量表达阳性[OX(+)]和阴性对照[OX(-)]材料,OsAAP6互补表达阳性[ZpZc(+)]和阴性对照[ZpZc(-)]试验材料。其中,亲本材料南洋占为粳稻品种,将来源于珍汕97(种子蛋白质含量较高亲本)的OsAAP6等位基因转入南洋占(种子蛋白质含量较低)得到OsAAP6互补转基因材料。OsAAP6超量表达和互补转基因水稻从播种到最后的种子成熟,均实行常规的田间管理及种植。
主要试剂:牛血清蛋白、考马斯亮蓝G-250和DL-二硫苏糖醇(DTT)购自美国Sigma公司;TRIzolreagent RNA提取试剂盒、聚丙烯酰胺和SuperScriptⅡI反转录酶购自美国Invitrogen公司,其余试剂均购自中国医药集团有限公司。主要仪器设备:LC-20AD纳升液相色谱仪(日本岛津公司)、LTQOrbi-trap Velos串联ESI质谱仪(美国ThermoFisher Scien-tific公司)、NIRStm DS 2500近红外谷物分析仪(丹麦FOSS公司)、MH-LG2018砻谷机(长春华茂仪器仪表有限公司)、JXFM110-IⅡ高速全自动锤式旋风磨(吉林赛亚斯科技有限公司)、SH-6600分光光度计(江苏盛奥华环保科技有限公司)和T4799-10G胰蛋白酶(美国Sigma公司)。
1.2 试验方法
1.2.1转基因材料中OsAAP6基因表达分析
分别取OsAAP6超量表达和互补转基因水稻开花后7 d的胚乳,迅速放人液氮中,然后利用TRIzol reagentRNA提取试剂盒提取总 RNA,利用olig(dT)n引物和 SuperScript II反转录酶反转录成cDNA第一链,以其为模板,采用实时荧光定量 PCR检测 OsAAP6基因的相对表达量(Peng et al.,2023)。
1.2.2 水稻胚乳蛋白的提取与含量检测 水稻种
子成熟后,自然晾干并在室温放置3个月后用于总蛋白和4类储藏蛋白的提取。将水稻OsAAP6超量表达和互补转基因的种子除去胚后,分别提取其胚乳中的总蛋白:胚乳采用旋风磨进行研磨为米粉,过100目筛子后将样品转入50mL圆底离心管中,加入适量Lysis Buffer3;冰浴超声5 min,4℃下15000r/min离心20min,取上清;加入5倍体积的10%TCA/冷丙酮,-20℃沉淀2 h;4℃下15000 r/min离心20min,去上清,沉淀转入1.5mL离心管;加1mL冷丙酮,终浓度为10mmol/LDTT,捣碎沉淀,-20℃沉淀30min,4℃下30000r/min离心15min,去上清,后续总蛋白的提取参照Paraman等(2007)的方法进行。参考Peng等(2014)的方法提取水稻胚乳中的储藏蛋白(谷蛋白、醇溶蛋白、球蛋白和清蛋白),然后采用考马斯亮蓝G-250法测定各种蛋白质的浓度。
1.2.3iTRAQ蛋白质组学检测
水稻OsAAP6超 量表达和互补转基因胚乳总蛋白经还原性烷基化处 理、定量、SDS电泳、酶解、iTRAQ标记与混合、强阳离子交换色谱(SCX)分离和液相色谱与串联质谱联用(LC-MS/MS)分析。iTRAQ蛋白质组学检测是由深圳华大基因科技服务有限公司参照Gan等(2010) 的方法完成。对鉴定到的蛋白进行定量,当上调或 下调蛋白的差异倍数(Fold Change)在1.5倍以上,且经统计检验Plt;0.05时,视为差异蛋白。参考序列来源于水稻GenBank数据库中共134525条序列信息。
2结果与分析
2.1转基因水稻中OsAAP6基因表达分析结果
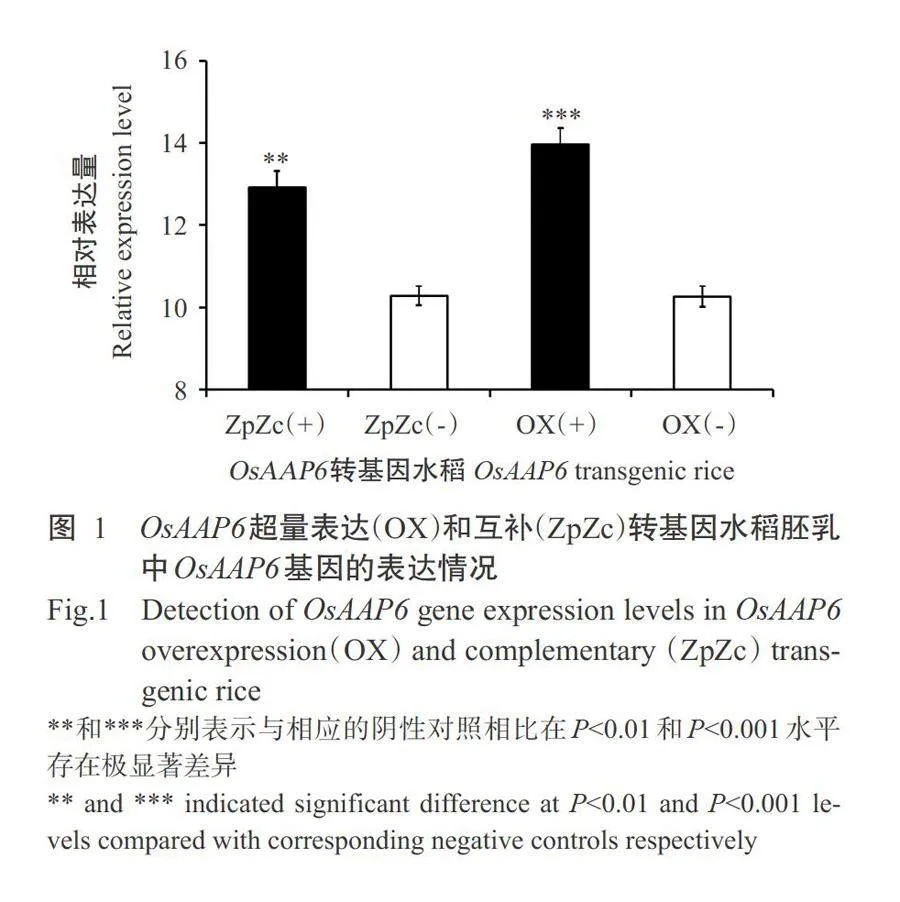
OsAAP6超量表达阳性[OX(+)]和OsAAP6互补表达阳性[ZpZc(+)]水稻开花后7d,分别取样检测其胚乳中的OsAAP6基因相对表达量,并进行比较分析,结果显示,与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性胚乳中OsAAP6基因的相对表达量分别在Plt;0.01和Plt;0.001水平极显著升高(图1),与本课题组前期研究结果(Peng et al.,2014;彭波等,2019)相吻合。
2.2OsAAP6转基因胚乳总蛋白提取质量检测结果
考马斯亮蓝G-250与蛋白在酸性条件下相结合,在一定范围内蛋白含量与595nm的吸光度呈正线性相关。将牛血清蛋白配成0~0.2μg/μL浓度梯度,与考马斯亮蓝G-250试剂反应2min后利用分光光度计在595nm检测其吸光度,进而绘制其标准曲线(图2)。根据水稻OsAAP6超量表达(OX)和互补(ZpZc)转基因水稻胚乳总蛋白溶液在595nm的吸光度,利用标准曲线回归方程得到OsAAP6转基因OX(+)、OX(-)、ZpZc(+)和ZpZc(-)胚乳总蛋白溶液浓度分别为1.36、0.85、0.76和0.92μg/μL。分别取OsAAP6转基因OX(+)、OX(-)、ZpZc(+)和ZpZc(-)胚乳总蛋白30.0μg,利用12%聚丙烯酰胺凝胶(PAGE)电泳检测,结果(图3)显示,提取的水稻OsAAP6超量表达(OX)和互补表达(ZpZc)转基因水稻胚乳总蛋白质量较好,能满足后续的酶解和iTRAQ蛋白组学分析等试验。
2.3iTRAQ蛋白质组学分析结果
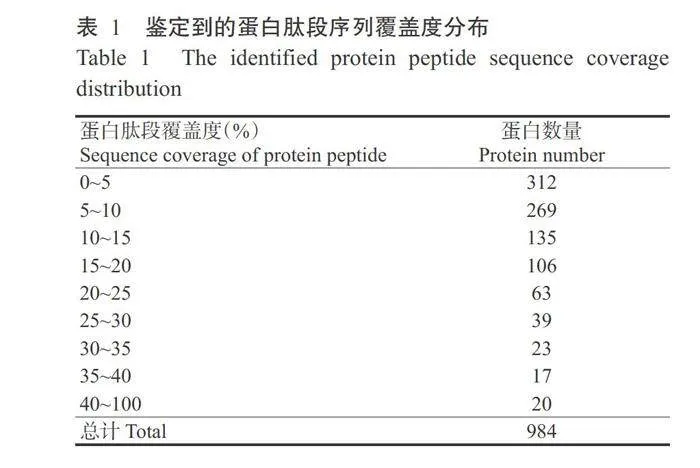
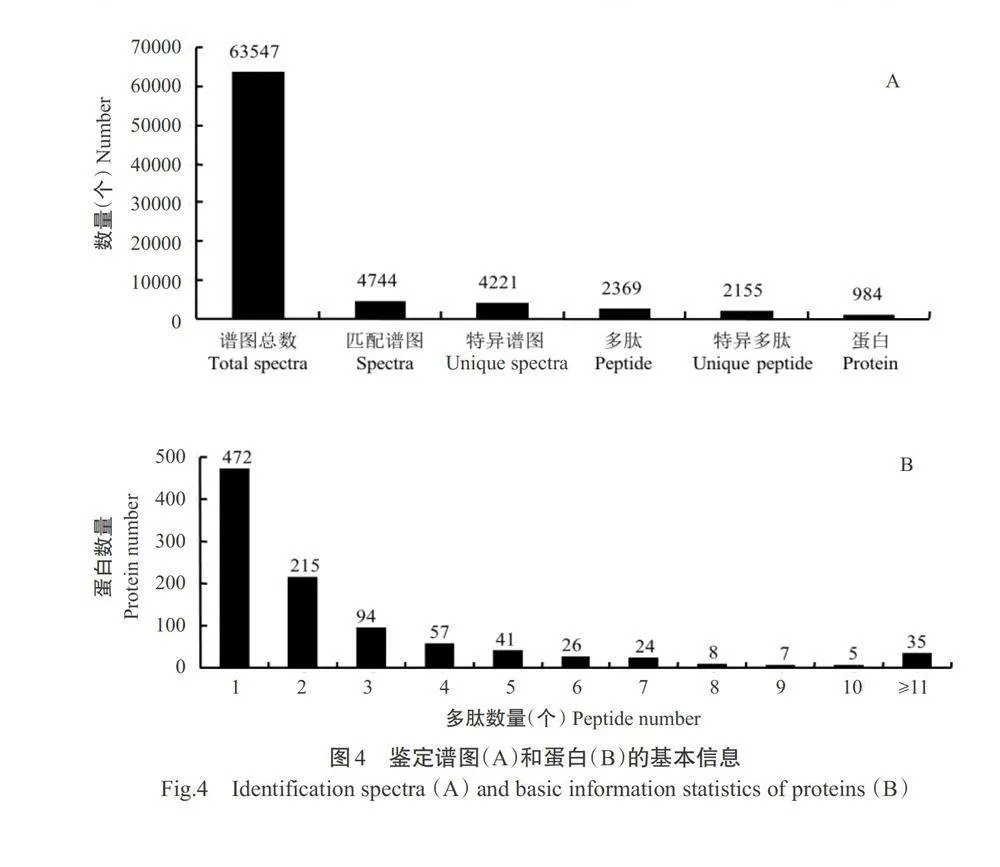
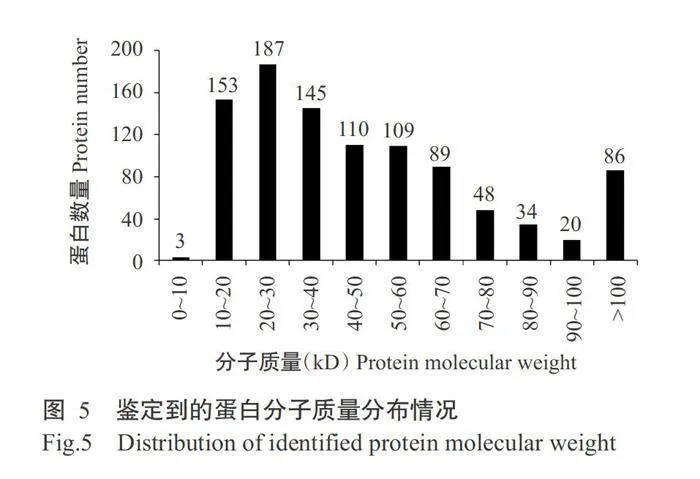
对水稻OsAAP6超量表达(OX)和互补(ZpZc)转基因水稻胚乳蛋白进行iTRAQ蛋白组学分析,共鉴定到63547个肽谱、2369个多肽和984种蛋白(图4-A)。通过对鉴定到的蛋白和多肽分析发现,超过47%的蛋白至少有2个或3个多肽支持(图4-B);鉴定到的所有蛋白依据其相对分子质量进行统计分析,结果显示超过70%的蛋白分子质量为10~70kD(图5)。针对鉴定到的蛋白进行肽段序列覆盖度分析,结果发现大部分鉴定到的蛋白均有多种多肽较好覆盖,其中超过30%的蛋白有10%~25%的多肽覆盖(表1)。
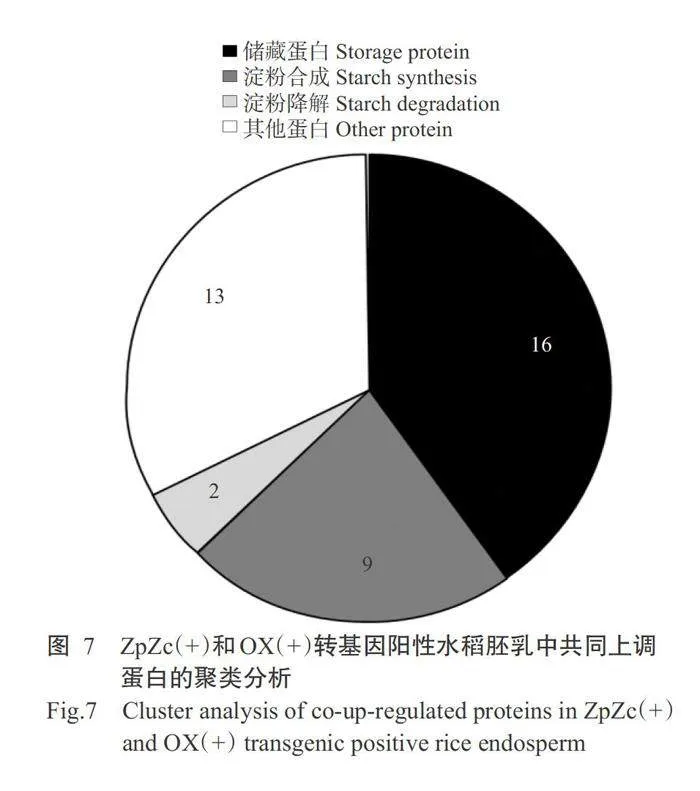
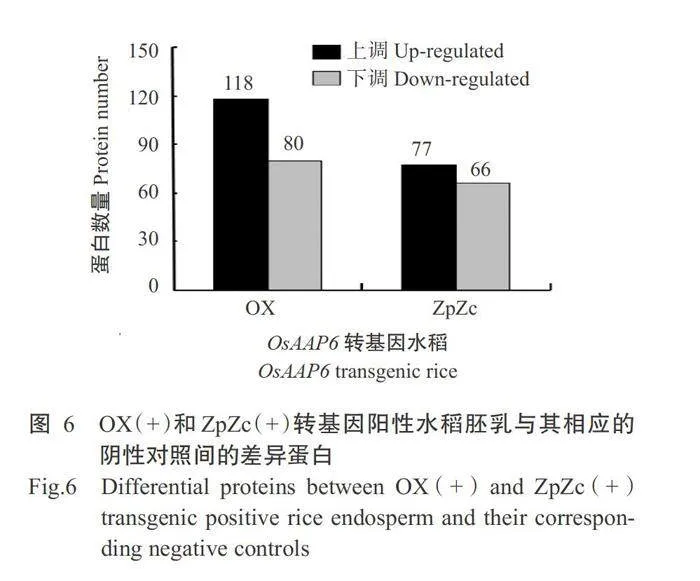
分别对OsAAP6超量表达(OX)和互补(ZpZc)转基因水稻胚乳蛋白中的差异表达蛋白进行分析,结果显示,与OX(-)和ZpZc(-)转基因阴性对照相比,在OX(+)和ZpZc(+)转基因阳性水稻胚乳中分别有118和77种蛋白上调,且分别有80和66种蛋白下调(图6)。由于OsAAP6超量表达(OX)和互补(ZpZc)转基因的亲本材料均为南洋占,即这2组转基因材料具有相同的遗传背景,能对其共同上调或者下调的蛋白进行深入分析。针对水稻OsAAP6超量表达(OX)和互补(ZpZc)转基因水稻胚乳中的差异蛋白进一步比较,结果发现,与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性水稻有58种差异蛋白共同上调或下调表达,其中有39种蛋白显著增加(Plt;0.05,下同),有19种蛋白显著降低(表2)。在39种共同上调表达的蛋白中有27种蛋白参与水稻种子储藏底物的合成和积累,且有16种共同上调表达的蛋白参与水稻种子储藏蛋白的代谢和积累过程(表2和图7)。
2.4OsAAP6转基因水稻胚乳储藏蛋白定量检测
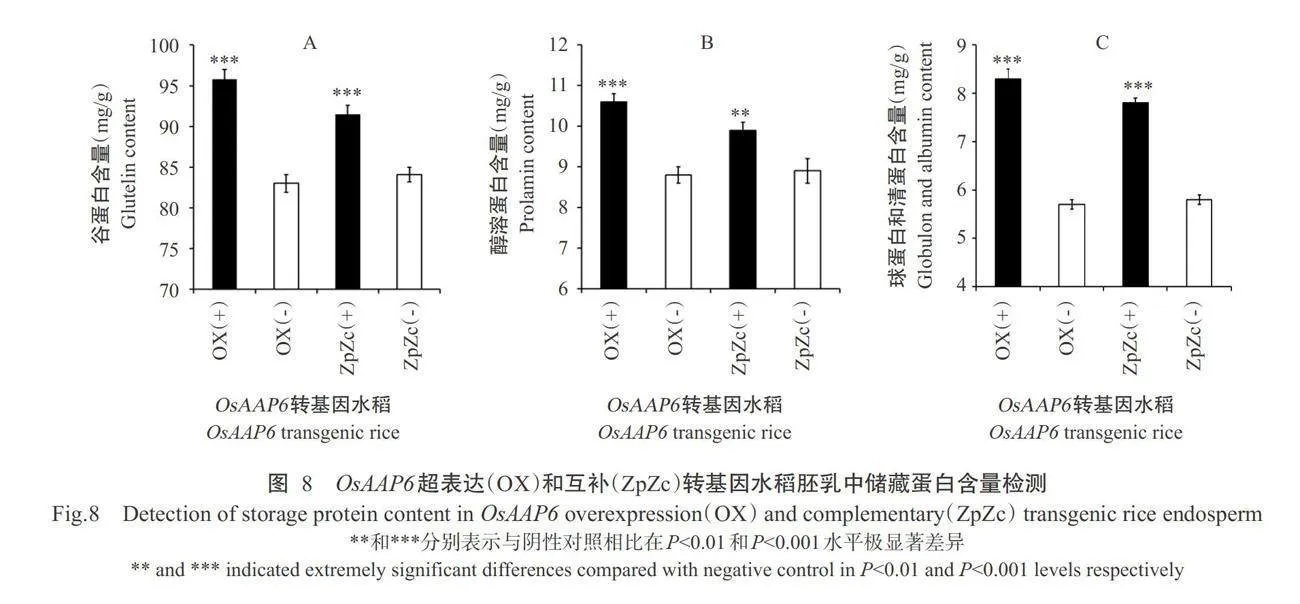
利用iTRAQ 蛋白质组学,针对OsAAP6超量表达(OX)和互补(ZpZc)转基因水稻胚乳中的蛋白进行分析,结果发现与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性水稻胚乳中有16种共同上调表达的蛋白均参与水稻种子储藏蛋白的代谢和积累过程,故再对 OsAAP6 超量表达(OX)和互补(ZpZc)转基因水稻胚乳中的4类储藏蛋白进行检测,结果如图8所示。与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性水稻胚乳中谷蛋白、醇溶蛋白、球蛋白和清蛋白含量均显著增加,特别是OX(+)转基因阳性水稻胚乳中谷蛋白、醇溶蛋白、球蛋白和清蛋白含量均高于ZpZc(+)转基因阳性水稻。
3讨论
从水稻基因组中,目前已筛选到与水稻种子蛋白质含量相关的数量性状位点(QTL)超过100个,在1~12号染色体上均有分布,针对这些QTLs位点的深入研究也取得积极进展(Varshney et al.,2020;Kim et al.,2021)。例如,调控水稻种子蛋白质含量的QTL基因qGPC-10,编码一个稻米储藏蛋白(谷蛋白)的前体蛋白OsGluA2,能显著影响稻米的营养品质性状(Yang et al.,2019;Wu et al.,2020)。本课题组前期分离克隆到OsAAP6基因,其位于水稻1号染色体上正向调控水稻种子蛋白质含量相关的QTL,编码的OsAAP6蛋白参与水稻根系对氨基酸的吸收与转运,并对稻米的营养品质等性状产生明显影响(Peng et al.,2014;彭波等,2019)。为解析OsAAP6基因对稻米胚乳中不同储藏表达蛋白积累的精细调控机制,本研究采用iTRAQ技术对OsAAP6超量表达(OX)和互补表达(ZpZc)转基因水稻胚乳进行蛋白质组学分析,结果显示,与ZpZc(-)和OX(-)转基因阴性对照相比,ZpZc(+)和OX(+)转基因阳性水稻共有58种差异蛋白共同上调或下调表达,其中有39种蛋白显著增加,有19种蛋白显著下调。针对这19种显著下调的蛋白进一步分析,结果发现有多种与支链淀粉合成和积累相关,如淀粉分支酶等,与本课题组前期研究结果(Peng etal.,2014)即OsAAP6超量表达(OX)和互补表达(ZpZc)转基因水稻种子中的总淀粉含量下降,直链淀粉含量升高相吻合,而对应的支链淀粉含量降低。因此,OsAAP6基因上调表达能抑制支链淀粉合成和积累相关的酶类,进而导致其种子中支链淀粉含量减少。
水稻种子主要营养物质包括蛋白质、淀粉和脂类物质,其中的淀粉和蛋白质占种子干重的90%左右(Chen et al.,2018;Yang et al.,2019;Long et al.,2023)。水稻胚乳中的蛋白质主要由储藏蛋白和功能蛋白两大类组成,储藏蛋白占总蛋白含量的90%左右(Taguchi et al.,2021)。水稻胚乳中的储藏蛋白主要是谷蛋白和醇溶蛋白,其中谷蛋白占胚乳中总蛋白含量的80%左右,且这些蛋白对稻米的营养品质及其他品质性状均具有重要影响(Zhou et al.,2017;彭波等,2019)。在OsAAP6转基因稻米胚乳中,OsAAP6超表达(OX)和互补(ZpZc)转基因阳性胚乳中的谷蛋白、醇溶蛋白、球蛋白和清蛋白含量均显著增加。大田增施氮肥条件下,OsAAP6互补(ZpZc)转基因阳性水稻籽粒中蛋白积累增加进而促进总蛋白含量的提高(彭波等,2019)。本研究通过iTRAQ蛋白质组学检测发现,与ZpZc(-)和OX(-)转基因阴性对照相比,OX(+)和ZpZc(+)转基因阳性水稻胚乳中有40种蛋白共同显著上调表达,其中有16种共同上调表达的蛋白(谷蛋白前体蛋白、谷蛋白B、醇溶蛋白前体蛋白和13kD醇溶蛋白等)参与水稻种子储藏蛋白的代谢和积累过程,与OX(+)和ZpZc(+)转基因阳性水稻胚乳中谷蛋白、醇溶蛋白、球蛋白和清蛋白含量均显著增加的结论相一致。上述试验结果与本课题组前期对OsAAP6基因表达及其对应胚乳中相关基因表达、种子蛋白质总含量的检测结果(Peng et al.,2014;彭波等,2019)相一致。因此,上调OsAAP6基因在水稻体内表达能促进其种子储藏蛋白的积累,进而提高胚乳中4类储藏蛋白含量。
4结论
水稻植株中OsAAP6基因的上调表达,能抑制与支链淀粉合成相关蛋白的积累,但促进其种子储藏相关蛋白的积累,最终提高胚乳中谷蛋白、醇溶蛋白、清蛋白和球蛋白含量,为高营养品质水稻新品种的培育提供了重要信息。
参考文献(References):
陈凌华,程祖锌,许明,郑金贵.2017.iTRAQ技术及其在水稻蛋白质组学中的应用研究进展[J].中国农业科技导报,19(12): 14-23. [Chen L H,Cheng Z X, Xu M,Zheng J G.2017. Research advance in iTRAQ technology and itsapplication in proteomics of rice [J]. Journal of Agricul-tural Science and Technology, 19(12) : 14-23.] doi: 10.13304/j.nykjdb.2017.0224.
高渐飞,周玮,刘妮,杨艳.2023.基于广泛靶向代谢组学分析黑老虎种子中氨基酸和脂质组分[J].中国油脂,48(1):87-93. [Gao J F, Zhou W, Liu N, Yang Y. 2023. Analysisof amino acids and lipids in Kadsura coccinea seed bywidely targeted metabolomics[J]. China Oils and Fats,48(1):87-93.J doi:10.19902/j.cnki.zgyz.1003-7969.210578.
兰艳,黄曌,胡明明,涂云彪,孙影影,隋晓东,龚静,李天.2019.施氮量对低谷蛋白水稻籽粒品质及蛋白质组分的影响[J].浙江农业学报,31(2):182-190.[LanY,HuangZ, Hu M M, Tu Y B, Sun Y Y, Sui X D, Gong J, Li T.2019. Effects of nitrogen application rate on quality andprotein components of low-gluten rice [J]. Acta Agricul-turae Zhejiangensis, 31 (2) : 182-190.] doi: 10.3969/j.issn.1004-1524.2019.02.02.
罗同平,周忠凤,庞朝雄.2023.低直链淀粉含量水稻三系不育系百香A的选育与应用[J].杂交水稻,38(1):40-43.[Luo T P,Zhou Z F,Pang C X. 2023. Breeding and appli-cation of CMS line Baixiang a with low amylose contentin rice[J]. Hybrid Rice, 38 (1) : 40-43.] doi: 10.16267/j.cnki.1005-3956.20220307.117.
彭波,何璐璐,田夏雨,辛晴晴,孙艳芳,宋晓华,孙玲,刘毓琛,庞瑞华,周棋赢,汪全秀,周伟,宋世枝.2019.OsAAP6基因在不同氮肥条件下的表达分析及其对水稻籽粒营养品质的影响[J].西南农业学报,32(9):1973-1979. [Peng B,He L L,Tian X Y,Xin Q Q,Sun Y F,SongX H, Sun L, Liu Y C,Pang R H,Zhou Q Y, Wang Q X,Zhou W, Song S Z. 2019. Expression analysis of OsAAP6gene under different nitrogen conditions and its effects ongrain nutritional quality in rice [J]. Southwest China Jour-nal of Agricultural Sciences, 32 (9) : 1973-1979.] doi: 10.16213/j.cnki.scjas.2019.9.002.
彭波,孙晓宇,张庆茜,彭娟,娄安琪,孙艳芳,庞瑞华,周伟,汪全秀.2023.水稻OsMDH基因的克隆及其生物信息学分析[J].信阳师范学院学报(自然科学版),36(2):243-248. [Peng B, Sun X Y,Zhang Q X,Peng J,Lou A Q, SunY F, Pang R H, Zhou W, Wang Q X. 2023. Cloning andbioinformatics analysis of OsMDH gene in rice [J]. Jour-nal of Xinyang Normal University (Natural Science Edi-tion),36(2):243-248.] doi:10.3969/j.issn.1003-0972.2023.02.014.
彭波,孙艳芳,庞瑞华,孔冬艳,宋晓华,李慧龙,李金涛,周棋赢,段斌,柳琳,宋世枝.2017.水稻种子蛋白质含量遗传研究进展[J].南方农业学报,48(3):401-407.[PengB,Sun Y F,Pang R H,Kong D Y,Song X H,Li H L,Li J T,Zhou Q Y, Duan B, Liu L, Song S Z. 2017. Genetic of riceseed protein content: A review[J]. Journal of SouthernAgriculture , 48 (3) : 401-407.] doi : 10.3969/j : issn.2095-1191.2017.03.004.
唐小涵,刘世家,刘喜,田云录,王云龙,滕烜,段二超,张元 燕,江玲,张文伟,王益华,万建民.2020.色氨酰-tRNA合成酶基因WRS1调控水稻种子发育[J].中国水稻科学,34(5):383-396.[Tang XH,Liu SJ,Liu X,Tian Y L, Wang Y L,Teng X,Duan E C,Zhang Y Y,Jiang L,Zhang W W,Wang Y H,Wan J M. 2020. Tryptophanyl-tRNA syn- thetase gene WRS1 regulates rice seed development [J].Chinese Journal of Rice Science, 34 (5) : 383-396.] doi:10.16819/j.1001-7216.2020. 0302.
杨陶陶,邹积祥,伍龙梅,包晓哲,江瑜,张楠,张彬.2023.开 放式增温对华南双季稻稻米品质的影响[J].中国水稻科学,37(1):66-77.[Yang TT,Zou JX,Wu LM,Bao XZ, Jiang Y, Zhang N, Zhang B. 2023. Effect of free air tem- perature increase on grainquality of double-cropping rice in south China[J]. Chinese Journal of Rice Science, 37(1):66-77.] doi:10.16819/j.1001-7216.2023.220402.
Bharali A, Baruah K K. 2022. Effects of integrated nutrient management on sucrose phosphate synthase enzyme activity and grain quality traits in rice [J]. Physiology and Molecular Biology of Plants, 28 (2) : 383-389. doi: 10.1007/s12298-022-01148-w.
Chen D D, Chai S C, McIntyre L C, Xue G P. 2018. Overexpression of a predominantly root-expressed NAC transcription factor in wheat roots enhances root length, biomass and drought tolerance [J]. Plant Cell Reports, 37 (2) : 225237. doi: 10.1007/s00299-017-2224-y.
Du J,Dang M Z,Jia YY,Xu Y J,Li C M.2022. Persimmontannin unevenly changes the physical properties, morphology, subunits composition and cross-linking types of glia-din and glutenin[J]. Food Chemistry, 387: 132913. doi: 10.1016/j.foodchem.2022.132913.
Fang Z M, Wu B W, Ji Y Y. 2021. The amino acid transporter OsAAP4 contributes to rice tillering and grain yield by regulating neutral amino acid allocation through two splicing variants[J]. Rice, 14(1) :2. doi: 10.1186/s12284-020-00446-9.
Gan C S, Chong P K, Pham T K, Wright P C. 2007. Technical experimental and biological variations in isobaric tags for relative and absolute quantitation (iTRAQ) [J]. Journal of Proteome Research, 6 (2) : 821-827. doi: 10.1021/pr060474i.
Guo Z Q, Huang Y Q,Huang J, Li S Y,Zhu Z Z, Deng Q C, Cheng S Y. 2022. Formation of protein-anthocyanin complex induced by grape skin extracts interacting with wheat gliadins: Multi-spectroscopy and molecular docking analysis [J]. Food Chemistry,385: 132702. doi: 10.1016/j.foodchem.2022.132702.
Ji Y Y,Huang W T,Wu B W,Fang Z M,Wang X L. 2020. The amino acid transporter AAP1 mediates growth and grain yield by regulating neutral amino acid uptake and reallocation in Oryza sativa [J]. Journal of Experimental Botany, 71(16):4763-4777. doi:10.1093/jxb/eraa256.
Kim E Y, Kim K D, Cho J. 2021. Harnessing epigenetic varia-bility for crop improvement:Current status and future prospects[J]. Genes amp; Genomics, 44(3) : 259-266. doi: 10.1007/S13258-021-01189-7.
Li H X,Li X Z,Wang G J,Zhang J H,Wang G Q. 2022. Analysis of gene expression in early seed germination of rice: Landscape and genetic regulation[J]. BMC Plant Biology,
22(1):70. doi:10.1186/s12870-022-03458-3.
Li Y,Xiao J H, Chen L L,Huang X H, Cheng Z K,Han B, Zhang Q F, Wu C Y. 2018. Rice functional genomics research: Past decade and future [J]. Molecular Plant, 11
(3):359-380. doi:10.1016/j.molp.2018.01.007.
Lin Z M,Zhang X C,Yang XY,Li G H,Tang S, Wang S H, Ding Y F, Liu Z H. 2014. Proteomic analysis of proteins related to rice grain chalkiness using iTRAQ and a novel comparison system based on a notched-belly mutant with white-belly[J]. BMC Plant Biology,14:163. doi:10.1186/
1471-2229-14-163.
Long X K, Guan C M, Wang L, Jia L T, Fu X J, Lin Q L, Huang Z Y, Liu C. 2023. Rice storage proteins: Focus on composition, distribution, genetic improvement and effects on rice quality [J]. Rice Science, 30 (3) : 207-221. doi: 10.
1016/J.RSCI.2023.03.005.
Lu K,Wu B W,Wang J,Zhu W,Nie H P,Qian J J,Huang W
T, Fang Z M. 2018. Blocking amino acid transporter OsAAP3 improves grain yield by promoting outgrowth buds and increasing tiller number in rice[J]. Plant Biotechnology Journal, 16(10): 1710-1722. doi:10.1111/pbi.12907.
Ma H Y, Sheng C, Qiao L L, Zhao H W, Niu D D. 2020. A comparative proteomic approach to identify defencerelated proteins between resistant and susceptible rice cultivars challenged with the fungal pathogen Rhizoctonia
solani[J]. Plant Growth Regulation,90(1):73-88. doi: 10.1007/s10725-019-00551-w.
Paraman I,Hettiarachchy N S, Schaefer C. 2007. Glycosylation and deamidation of rice endosperm protein for improved solubility and emulsifying properties[J]. Cereal Chemistry,84(6):593-599. doi:10.1094/CCHEM-84-6-0593.
Peng B,He L L,Tan J,Zheng L T,Zhang J T,Qiao Q W,Wang
Y,Gao Y,Tian X Y,Liu Z Y,Song X H,Sun Y F,Pang R
H, Li J T, Yuan H Y. 2019. Effects of rice aging on its main nutrients and quality characters[J]. Journal of Agricultural Science, 11 (17) : 44-56. doi: 10.5539/jas.vl1n17 p44.
Peng B,Kong H L,Li Y B,Wang L Q,Zhong M,Sun L,Gao G J,Zhang Q L, Luo L J,Wang G W,Xie W B, Chen J X, Yao W,Peng Y,Lei L,Lian X M,Xiao J H,Xu C G,Li X H, He Y Q. 2014. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice [J]. Nature Communications, 5(1) : 4847. doi: 10.1038/ ncomms5847.
Peng B,Sun X Y,Tian X Y,Kong D Y,He L L,Peng J,Liu Y, Guo G Y,Sun Y F,Pang R H,Zhou W,Zhao J H,Wang Q
X. 2023. OsNAC74 affects grain protein content and various biological traits by regulating OsAAP6 expression in rice[J]. Molecular Breeding, 43 (12) : 87. doi: 10.1007/s11032-023-01433-w.
Peng B,Zhang Q X, Tian X Y,Sun Y F,Huang X H,Pang R
H, Wang Q X,Zhou W, Yuan H Y, Yang F, Peng J, Li H
L, Song X H, A X X. 2021. Influencing factors of grain nutritional quality and its genetic improvement strategy in rice[J]. Journal of Biotechnology Research,7(1):1-11. doi: 10.32861/JBR.71.1.11.
Shewry P R, Halford N G. 2002. Cereal seed storage proteins: Structures, properties and role in grain utilization[J]. Journal of Experimental Botany, 53 (370) : 947-958. doi: 10.1093/jexbot/53.370.947.
Sung J,Kim W J,Oh T K,So Y S. 2023. Nitrogen (N) use efficiency and yield in rice under varying types and rates of N source: Chemical fertilizer, livestock manure compost and food waste-livestock manure compost[J]. Applied Biological Chemistry,66(1):4. doi:10.1186/S13765-022-00766-Y.
Taguchi S,Kawai T, Nakagawa T,Nakamura Y,Kamei J,Obinata D, Yamaguchi K, Kaneko T, Kakutani S, Tokunaga M, Uemura Y, Sato Y, Fujimura T, Fukuhara H, Enomoto
Y, Nishimatsu H, Takahashi S, Kume H. 2021. Prognostic significance of the albumin-to-globulin ratio for advanced urothelial carcinoma treated with pembrolizumab: A multicenter retrospective study [J]. Scientific Reports, 11 (1) :15623. doi:10.1038/S41598-021-95061-Z.
Tian Z X,Qian Q,Liu Q Q, Yan M X,Liu X F, Yan C F, Liu G
F,Gao Z Y,Tang S Z,Zeng D L,Wang Y H,Yu J M,Li J
Y. 2009. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities [J].
Proceedings of the National Academy of Sciences of theUnited States of America, 106 (51) : 21760-21765. doi: 10.1073/pnas.0912396106.
Varshney R K, Sinha P,Singh V K,Kumar A,Zhang Q F, Bennetzen J L. 2020. 5Gs for crop genetic improvement [J].Current Opinion in Plant Biology, 56: 190-196. doi: 10.1016/j.pbi.2019.12.004.
Wang J, Wu B, Lu K, Wei Q, Qian J J, Chen Y P, Fang Z M.
2019. The amino acid permease OsAAP5 regulates tiller number and grain yield in rice [J]. Plant Physiology, 180(2):1031-1045. doi:10.1104/pp.19.00034.
Wang S Y, Yang Y H, Guo M,Zhong C Y, Yan C J, Sun S Y.
2020. Targeted mutagenesis of amino acid transporter genes for rice quality improvement using the CRISPR/ Cas9 system[J]. Crop Journal, 8 (3) : 457-464. doi: 10.
1016/j.cj.2020.02.005.
Wang S Z, Chen W Y,Xiao W F,Yang C D,Xin Y,Qiu J R, Hu W M,Ying W,Fu Y P,Tong J X,Hu G C,Chen Z Z, Fang X P,Yu H,Lai W G,Ruan S L,Ma H S. 2015. Differential proteomic analysis using iTRAQ reveals alterations in hull development in rice (Oryza sativa L.) [J].
PLoS One, 10 (7) : e0133696. doi: 10.1371/journal. pone.
0133696. Wu Q, Liu Y F, Xie Z Z, Yu B, Sun Y, Huang J L. 2022. OsNAC016 regulates plant architecture and drought tolerance by interacting with the kinases GSK2 and SAPK8 [J]. Plant Physiology, 189 (3) : 1296-1313. doi: 10.1093/ plphys/kiac146.
Yang G, Wei X L, Fang Z M. 2022. Melatonin mediates axillary bud outgrowth by improving nitrogen assimilation and transport in rice [J]. Frontiers in Plant Science, 13: 900 262. doi: 10.3389/fpls.2022.900262.
Yang X Y, Yang G, Wei X L, Huang W T,Fang Z M. 2023. OsAAP15, an amino acid transporter in response to nitrogen concentration, mediates panicle branching and grain yield in rice[J]. Plant Science,330: 111640. doi: 10.1016/j.plantsci.2023.111640.
Yang Y H,Guo M,Sun S Y,Zou Y L, Yin S Y,Liu Y N,Tang S Z, Gu M H, Yang Z F, Yan C J. 2019. Natural variation of OsGluA2 is involved in grain protein content regulation in rice [J]. Nature Communications, 10 (1): 1949. doi: 10.1038/s41467-019-09919-y.
Zhao H M, Ma H L, Yu L, Wang X,Zhao J Z. 2018. Genomewide survey and expression analysis of amino acid transporter gene family in rice(Oryza sativa L.)[J]. PLoS One,
7(11) : e49210. doi: 10.1371/journal.pone. 0049210. Zhou W, Wang X,Zhou D, Ouyang Y, Yao J L. 2017. Overex-pression of the 16-kDa-amylase/trypsin inhibitor RAG2 improves grain yield and quality in rice[J]. Plant Biotechnology Journal,15(5):568-580. doi:10.1111/pbi.12654.
Zi J,Zhang J Y,Wang Q H,Zhou B J,Zhong J Y,Zhang C L, Qiu X M, Wen B, Zhang S Y, Fu X Q, Lin L, Liu S Q.
2013. Stress responsive proteins are actively regulated during rice (Oryza sativa) embryogenesis as indicated by quantitative proteomics analysis [J]. PLoS One, 8 (9):
e74229。doi:10.1371/journal . pone . 0074229。
(责任编辑陈燕)
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27 19:44:27
军事文摘(2021年22期)2021-11-26 00:43:51
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
肉类研究(2016年12期)2017-01-12 17:25:11
天津农业科学(2016年12期)2017-01-11 20:11:57
河北渔业(2016年5期)2016-09-08 10:52:23
课程教育研究·学法教法研究(2016年10期)2016-06-16 16:04:38
天津农业科学(2015年12期)2015-12-03 23:42:08
分析化学(2015年10期)2015-11-03 07:14:40
