
连作木薯对土壤理化性质及细菌群落演替的影响
2024-08-22 00:00:00彭晓辉黎亮武欧桂宁黄苑航彭晓雪阳太亿甘李申章佑韦茂贵
南方农业学报 2024年4期





摘要:【目的】研究连作木薯对非根际和根际土壤理化性质变化及细菌群落演替的影响,为系统阐明木薯连作障碍形成机制及其缓解措施研发提供理论依据。【方法】以木薯品种华南205为试验材料,在撂荒地上连续种植3年,每年木薯块根成熟期(植后250d)采集土壤样品。结合土壤农化分析、高通量测序技术和生物信息学手段,研究连作木薯对非根际和根际土壤理化性质及细菌群演替的影响。【结果】与种植第1年木薯相比,连作2年和3年的木薯非根际和根际土壤pH及有机质、碱解氮和速效磷含量降低,脲酶活性提高,除速效磷外,其他指标的变化均达显著水平(Plt;0.05,下同)。连作木薯会引起土壤细菌群落多样性及丰富度降低,其中根际土壤细菌群落多样性及丰富度下降速度比非根际土壤慢。连作木薯非根际与根际土壤细菌群落的分类和系统发育Beta多样性存在显著差异。连作木薯非根际和根际土壤的优势菌门均为放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、厚壁菌门(Firmicutes)、酸杆菌门(Acidobacteriota),主要优势菌属均为芽孢杆菌属(Bacillus)、norank f Roseiflexaceae、norank_f_norank_o_Vicinamibacterales、norank_f_norank_o_Gaiellales。在操作分类单元(OTU)水平,不同连作年限木薯的非根际和根际土壤细菌群落组成存在显著或极显著(Plt;0.01)差异。土壤的细菌群落结构与理化性质相互影响,对细菌群落影响较大的环境因子排序为碱解氮含量gt;有机质含量gt;pHgt;速效磷含量gt;脲酶活性。
【结论】连作会导致木薯非根际和根际土壤理化性质、细菌群落多样性和丰富度发生变化,土壤理化性质和细菌群落结构均在连作第3年出现明显劣化。木薯土壤理化性质变化与土壤细菌群落结构差异存在一定相关性,有益菌丰度降低可能是木薯连作造成土壤细菌群落劣化演替的重要原因之一。
关键词:木薯;连作;非根际土壤;根际土壤;细菌群落
中图分类号:S533
文献标志码:A
文章编号:2095-1191(2024)04-0942-12
Effects of continuous cropping of cassava on soil physicochemi-cal properties and bacterial community succession
PENG Xiao-hui1, LI Liang-wu1, OU Gui-ning1, HUANG Yuan-hang1, PENG Xiao-xue1,YANG Tai-yi1, GAN Li1, SHEN Zhang-you2*, WEI Mao-gui1,3*
(1College of Agriculture, Guangxi University, Nanning, Guangxi 530004, China; 2Cash Crops Research Institute, Guangxi Academy of Agricultural Sciences,Nanning, Guangxi 530007, China; 3Guangxi Key Laboratory of Agro-environment and Agro-product Safety, Nanning, Guangxi 530004, China)
Abstract:[Objective]The aim of this study was to investigate how continuous cropping of cassava affected the physicochemical properties of non-rhizosphere and rhizosphere soils, as well as the succession of bacterial communities, and thus providing a theoretical basis for understanding the formation of cassava continuous cropping obstacles. 【Method】The cassava variety South China 205 was continuously cultivated in an abandoned field for three years. The soil were sampled at cassava root tuber maturity stage (250 d after planting) in each year. Combining soil agrochemical analysis, high-throughput sequencing technology and bioinformatics methods, effects of continuous cassava on non-rhizosphere and rhizosphere soil physicochemical properties and bacterial group succession were studied. 【Result] The results showed that pH, organic matter, alkali-hydrolyzed nitrogen and available phosphorus contents of non-rhizosphere and rhizosphere soil of cassava were decreased and urease activity was increased in two and three years of continuous cropping compared with the first year of planting, and except available phosphorus content, the changes of other indexes reached significant level (Plt;0.05, the same below). Continuous cropping of cassava led to a decrease in the diversity and richness of soil bacterial communities. However, the decline rate in rhizosphere soil bacterial communities was slower than that in non-rhizosphere soils. There were significant differences in the classification and phylogenetic Beta diversity of non-rhizosphere and rhizo- sphere soil bacterial communities of continuous cropping cassava. The dominant bacterial phyla in non-rhizosphere and rhi- zosphere soils were Actinobacteriota, Proteobacteria, Chloroflexi, Firmicutes and Acidobacteriota. The main dominant bac- terial genera were Bacillus, norank f_Roseiflexaceae, norank f_norank_o_Vicinamibacterales and norank _f_norank_o_ Gaiellales. At the operational taxonomic unit (OTU) level, significant or extremely significant(Plt;0.01) differences were found in the composition of soil bacterial communities in non-rhizosphere and rhizosphere soils of cassava at different con- tinuous cropping years. The structure of soil bacterial communities was mutually affected by soil physicochemical proper- ties, the environmental factors affected bacterial community ranked as alkaline hydrolysis nitrogen content gt; organic mat- ter content gt; pH gt; available phosphorus content gt; urease activity. 【Conclusion ]Continuous cropping can lead to changes in non-rhizosphere and rhizosphere soil physicochemical properties, bacterial community diversity and richness, and soil physicochemical properties and bacterial community structure of cassava greatly deteriorate in the third year of continuous cropping. Soil physicochemical properties are correlated to structure of bacterial communities at certain extent. Notably, the decrease in the abundance of beneficial bacteria might be an important reason for the deterioration of soil bacterial community succession caused by cassava continuous cropping.
Key words: cassava; continuous cropping; non-rhizosphere soil; rhizosphere soil; bacterial community
Foundation items: National Natural Science Foundation of China (31960389,31860347)
0 引言
【研究意义】随着人口的不断增加和城镇化进程的加快,我国耕地资源日益减少。土地资源高度集约化规模管理、提高土地和肥料利用率已成为现代农业发展的趋势。现代农业逐渐形成了复种指数高、作物规模化种植且种类单一的特点。受土地资源、气候条件、种植习惯及经济效益等因素影响,同种或同科作物连作不可避免,许多作物存在着不同程度的连作障碍,已成为影响作物产量提高的关键问题之一(翁佩莹和郑红艳,2020)。木薯是我国重要的热带作物之一,属高光效C3作物,其理论产量最高可达120t/ha,大田种植高产记录也高达80t/haa但实际生产中因木薯具有耐旱耐贫瘠特性,我国木薯大多连年种植在南方相对贫瘠的边际土地上,广种薄收,连作现象普遍,部分地块连作年限长达15年以上,块根平均产量仅16~17 t/ha(梁海波等,2017)。连作障碍已被证实是我国木薯单产显著下降的重要因素之一(王战,2016;梁海波等,2017)。此外,为保证产量,化肥和农药使用量随木薯连作年限的增加而逐年增加,进而造成土壤酸化、重金属污染和水体污染等农业面源污染问题。因此,研究木薯连作障碍及其缓解措施,对提高木薯产量、减少化肥施用和保护生态环境均具有重要意义。【前人研究进展】连作障碍的形成是土壤、植物和微生物综合作用的结果,其形成机理相对复杂。根据前人研究结果,引起连作障碍的主要原因可归纳为作物化感物质的自毒作用、土壤理化性质劣化及土壤生物学环 境恶化(侯乾等,2019)。酚酸类、黄酮类和生物碱类 物质是作物的主要自毒物质种类(陈福慧等,2022)。 作物的化感物质主要通过影响植株的细胞膜透性、 光合作用、呼吸作用、有丝分裂、酶活性、蛋白质合成 等制约植物生长,进而造成作物产量及品质降低(杨 浩娜等,2022)。连作还会改变土壤理化性质,如连 作木薯使土壤容重变大,三相比例劣化,有机质、碱 解氮、速效钾和速效磷含量下降,根际土壤总酚酸含 量增加,土壤酸化严重(周贵靖,2017);连作大豆能 提高土壤中速效氮、速效磷和速效钾含量(刘株秀 等,2019);连作灵芝时土壤pH、有机质、速效钾、速 效磷和碱解氮含量均下降(卢孟召等,2022)。土壤 理化性质的改变会影响土壤微生物的丰富度(Hon- toria et al.,2019)。长年连作木薯的土壤中细菌种群 数量减少,真菌种群数量增加,根际微生物群落向真 菌型转化(刘珊廷,2020);苹果连作后土壤中的细菌 多样性减少,真菌多样性增加(王晓琪等,2020);连 作大豆和玉米均会显著降低土壤中真菌的丰富度和 多样性(宋秀丽等,2022)。此外,土壤微生物包含有 益菌群和有害菌群,有益菌群对作物生长发育起促 进作用,而有害菌群会影响作物正常的生长发育(孙 子欣和蔡柏岩,2022)。由此可见,不同作物连作会 对土壤理化性质及土壤微生物群落产生不同影响。 【本研究切入点】目前,关于木薯连作障碍的研究主 要集中在土壤理化性质方面(周贵靖,2017),也有少量连作对土壤微生物影响的报道(刘珊廷,2020),但 木薯连作对土壤理化性质和微生物群落演替及其相 关性影响的研究鲜有报道。【拟解决的关键问题】以 木薯品种华南205为试验材料,在撂荒地上连续种 植3年,利用高通量测序技术和生物信息学手段,研 究连作木薯对非根际和根际土壤理化性质变化及细 菌群落演替的影响,为系统阐明木薯连作障碍形成 机制及其缓解措施研发提供理论依据。
1材料与方法
1.1试验材料
供试材料为广西主栽木薯品种华南205。
1.2试验方法
试验于2019年4月-2021年12月在广西南宁 进行。试验地撂荒多年,其0~20cm土层pH6.24, 含有机质17.23 g/kg、碱解氮60.38 mg/kg、速效磷 45.09mg/kg、速效钾42.17mg/kg。常规耕作地块,犁地后使用卧式旋耕耙两轮。田块划分成12个面 积为70m²(14m×5m)的试验小区网格,第1年随机 挑选3个小区作为重复,此后2年仍定位原小区作 为当年重复。木薯分别于2019年4月16日、2020年4月10日和2021年4月11日种植。种茎长度约15cm, 株行距1m×1m,每小区共种植70株木薯。试验期间无人工灌水,木薯田间管理方式一致。试验期间 降水量和月平均气温如图1所示。
于木薯块根成熟期(植后250d)采集土壤样品。根际土壤:每小区随机选择3株木薯,挖出完整木薯 块根和纤维根,采用抖根法去除大块土壤,用毛刷采 集附着在根系上的土壤。非根际土壤:各小区按S 形随机选取5个点,分别用取土器采集0~20cm土 样,混匀后作为该小区的非根际土壤样品。土壤样 品置于塑料薄膜上,剔除植物残体、石砾等杂物,混 匀后以四分法分为3份。第1份立刻用液氮速冻,置于冰盒中带回实验室于-80°C保存,用于土壤微生 物群落多样性测定;第2份做好标记,带回实验室于 冰箱4°℃保存,用于土壤酶活性测定;第3份置于标 记好的自封袋内,常温保存带回实验室,风干后磨碎 过筛,用于土壤理化性质测定。
1.3土壤理化性质及酶活性测定
土壤pH采用酸度计法测定,有机质含量采用重 铬酸钾容量法测定,碱解氮含量采用碱解扩散法测 定,速效磷含量使用钼酸比色法测定,速效钾含量采 用NH2OAc浸提一火焰光度法测定。土壤脲酶活性 采用苯酚一次氯酸钠比色法测定。以上指标测定均 参照覃锋燕等(2022)的方法。
1.4土壤微生物高通量测序及生物信息学分析
参照覃锋燕等(2022)的方法,略有改动:使用E.Z.N.A.®Soil DNA Kit(美国Omega Bio-Tek公司) 提取土壤基因组DNA,用特异引物对16SrRNA序列的V3~V4可变区进行PCR扩增。PCR产物经2% 琼脂糖凝胶回收,用AxyPrep DNA Gel Extraction Kit(美国Axygen公司)进行纯化,2%琼脂糖凝胶 电泳检测,并用QuantusTMFluorometer(美国Promega 公司)进行定量检测。使用NEXTflex®Rapid DNA-Seq Kit(美国Bioo Scientific公司)构建文库,委托上海美吉生物医药科技有限公司利用Illumina MiSeqPE300平台进行土壤样品高通量测序。原始数据上传至NCBISRA数据库。分别在 Trimmomatic(version 0.32)和FLASH(version 1.2.11)上进行原始序列质控和拼接,在97%相似度下使用UPARSE Version7.1将序列进行操作分类单元(OTU)聚类。基于Silva数据库,设置70%的比对阈值,在RDP Classifier(http://rdp.cme.msu.edu/)上对筛选后的非嵌合序列进行物种分类注释。细菌OTU丰富度及Alpha多样性分析分别使用Past3(http://fold.uio.no/ohammer/past)和Mothur(ver-sion v.1.30.2)进行计算。细菌Beta多样性分析包含主成分分析、置换多因素方差分析和冗余分析,均使用R(version 3.3.1)vegan包完成分析并制作相应图片。用stats包进行细菌在属水平上的组间差异显著性检验并制图。
1.5统计分析
使用Excel 2010和SPSS 25.0进行数据整理和统计分析;使用SIMCA14.0进行主成分分析及制图。
2结果与分析
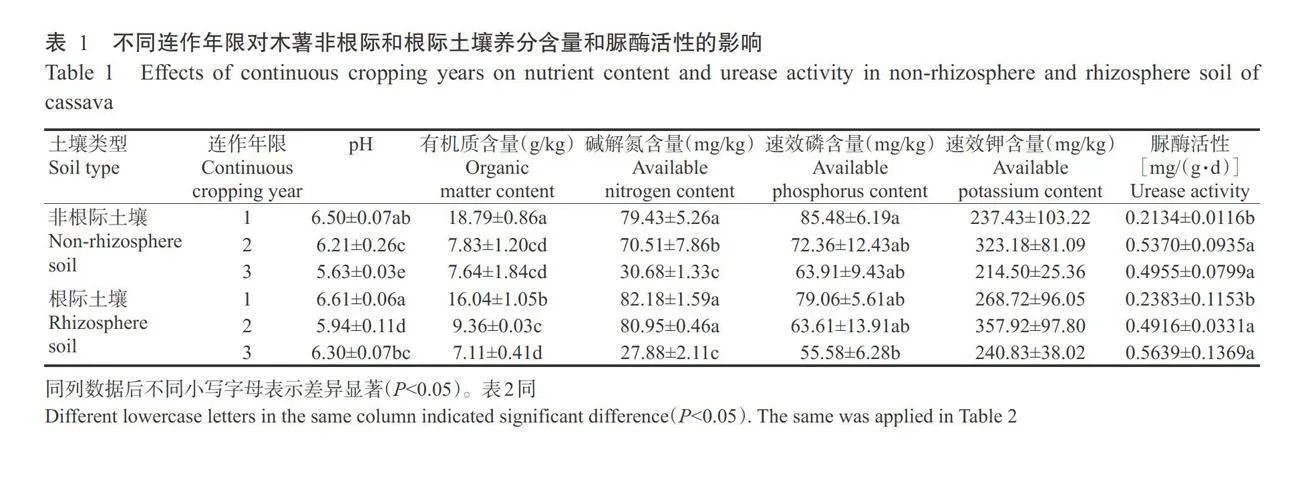
2.1连作木薯对非根际和根际土壤理化性质的影响
由表1可知,随连作年限增加,木薯非根际土壤的pH逐年下降,根际土壤的pH先下降后上升,年际间差异显著(Plt;0.05,下同);非根际土壤和根际土壤的有机质、碱解氮和速效磷含量均随连作年限的增加逐年下降,而速效钾含量在年际间均无显著差异(Pgt;0.05,下同)。随连作年限增加,非根际土壤的脲酶活性先升高后降低,根际土壤的脲酶活性逐年升高,连作2年和3年处理间无显著差异,但均显著高于连作1年处理。
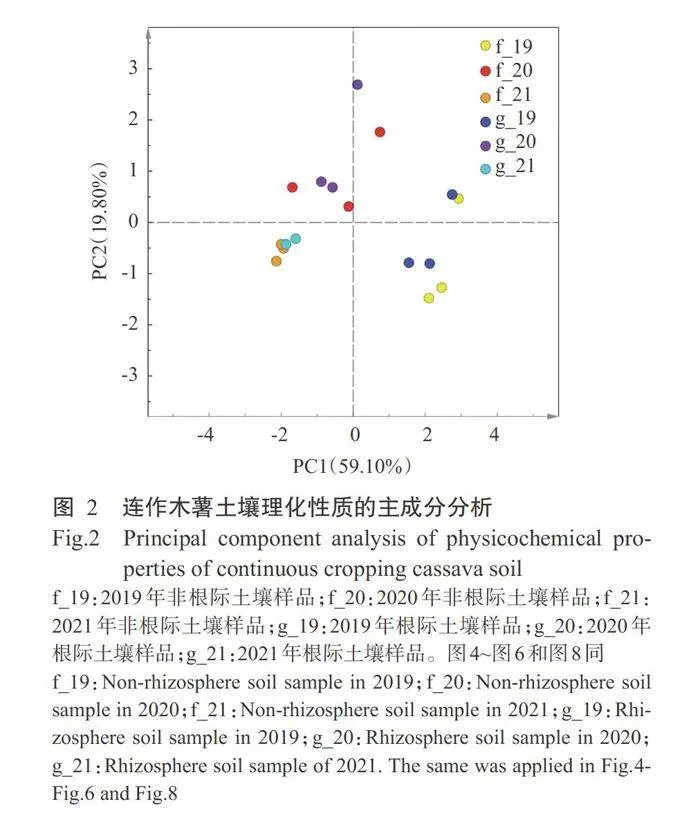
由图2可看出,第一主成分(PC1)解释了总方差 的59.10%,按年份将根际土壤与非根际土壤大致分 成3个模块,说明连作年限是引起木薯根际土壤和 非根际土壤养分含量及脲酶活性差异的主要原因, 连作会引起木薯土壤理化性质劣化。
2.2连作木薯对非根际和根际土壤细菌群落多样性及结构的影响
2.2.1Alpha多样性分析表2为不同处理细菌群
落多样性指数(Shannon指数和Simpson指数)和群落丰富度指数(Sobs指数、ACE指数和Chaol指数)。各处理的OTU文库覆盖率均超过95.00%。随连作年限增加,木薯非根际土壤的细菌Shannon指数呈 下降趋势,连作1年与连作3年间差异显著,而根际土壤的细菌Shannon指数无显著变化;非根际和根 际土壤的细菌Simpson指数在年际间均无显著差 异。随连作年限增加,非根际和根际土壤的细菌 Sobs指数均呈下降趋势,ACE指数先上升后下降; 非根际土壤细菌Chao1指数随连作年限增加而逐年 下降,根际土壤细菌Chao1指数则先上升后下降。由此可知,连作木薯根际与非根际土壤细菌群落的 Alpha多样性逐年降低,但根际土壤细菌群落的Alpha多样性降低速度较非根际土壤慢。
2.2.2Beta多样性分析
主成分分析结果(图3)表明,PC1将非根际土壤与根际土壤连作1年和2年的 样本与连作3年的样本分为2个群体,分别解释总方 差的17.18%和17.87%,故推测连作可引起细菌群落 Beta多样性变化,连作3年差异达显著水平。由表3可知,基于细菌群落半度量距离(Bray-Curtis)和未加权Unifrac距离,使用置换多因素方差分析方法分 别计算连作年限与土壤类型对细菌群落变化的贡 献,再次验证了连作年限是影响土壤细菌群落分类 和系统发育Beta多样性差异的主要因素,而土壤类型对Beta多样性差异的影响不显著。
2.2.3不同处理细菌群落组成分析
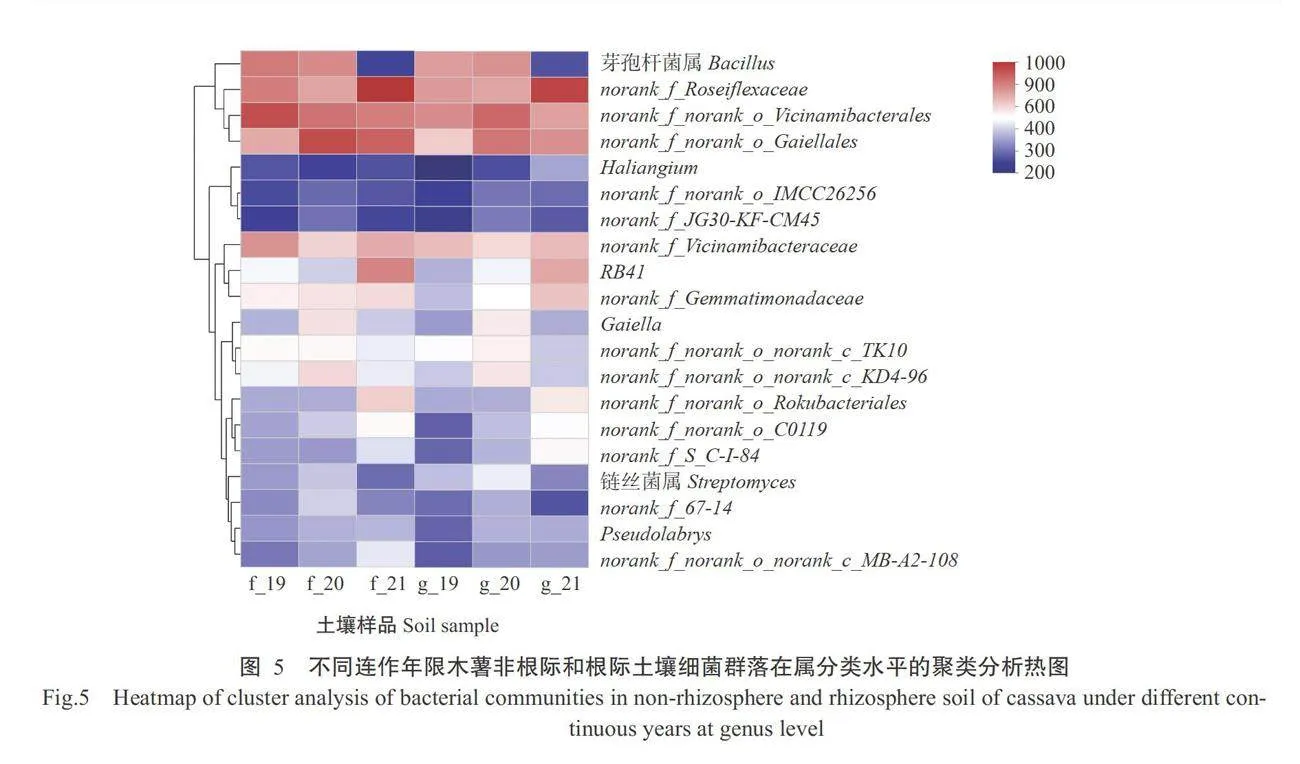
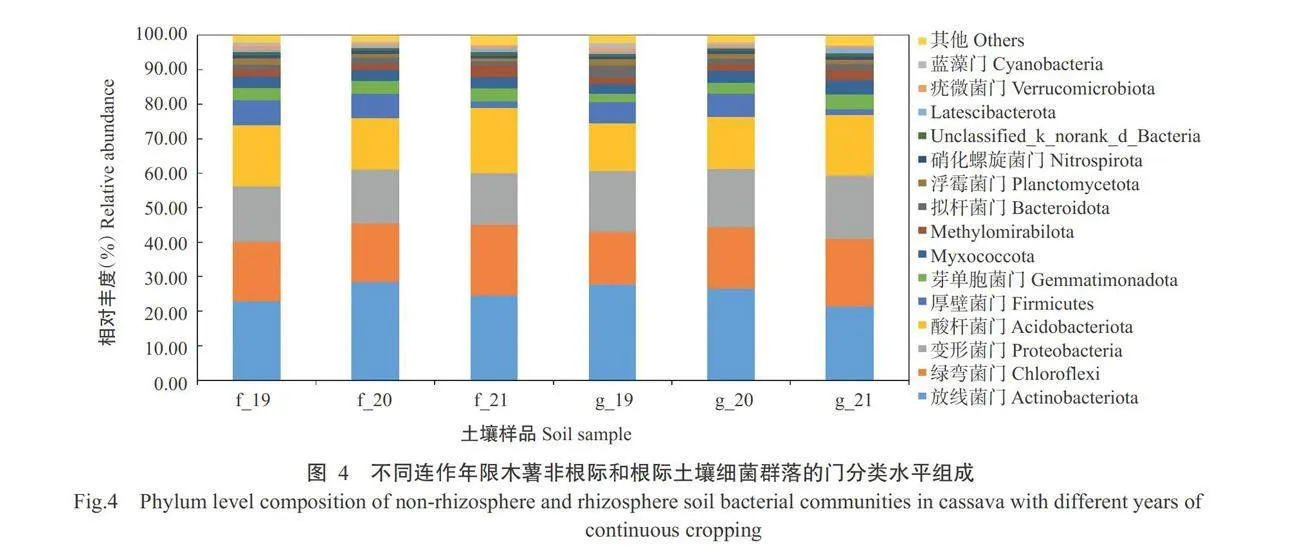
从图4可看 出,不同连作年限木薯的非根际和根际土壤细菌门 水平的群落结构组成基本相同,主要有放线菌门(Actinobacteriota)、绿弯菌门(Chloroflexi)、变形菌 门(Proteobacteria)、酸杆菌门(Acidobacteriota)、厚壁 菌门(Firmicutes)、芽单胞菌门(Gemmatimonadota)、 Myxococcota、Methylomirabilota、拟杆菌门(Bacte- roidota)、浮霉菌门(Planctomycetota)、Latescibacte- rota、蓝藻门(Cyanobacteria)和疣微菌门(Verrucomi- crobiota)等。其中,占主导地位的菌门为放线菌门 (占比19.18%~32.74%)、绿弯菌门(占比13.09%~21.56%)、变形菌门(占比14.08%~20.73%)、酸杆菌门(占比10.66%~22.83%)和厚壁菌门(占比1.16%~8.72%),占总相对丰度的77.14%~85.34%。从图5可看出,在属分类水平中,芽孢杆菌属(Bacillus)、norank f_ Roseiflexaceae、 norank f_norank_o_Vicina-mibacterales 、norank_f_norank_o_Gaiellales为土壤细菌的主要优势菌群。其中,根际土壤中芽孢杆菌属的相对丰度随着连作年限的增加呈先增加后下降变化趋势,非根际土壤中则是呈逐年下降趋势。norank f_Roseiflexaceae的相对丰度在根际土壤中呈先升高后降低趋势,在非根际土壤中呈连续下降趋势。非根际土壤中norank f_norank_o_Vicina-mibacterales的相对丰度随着连作年限增加呈下降趋势,根际土壤中则先升高后降低。norankfnorank o Gaiellales的相对丰度在非根际土壤与根际土壤中均先升高后降低。综上所述,不同连作年限木薯非根际和根际土壤细菌在门分类水平和属分类水平的群落组成基本相同,但其细菌群落相对丰度随连作年限增加而呈不同变化趋势。
2.2.4细菌群落组成差异分析
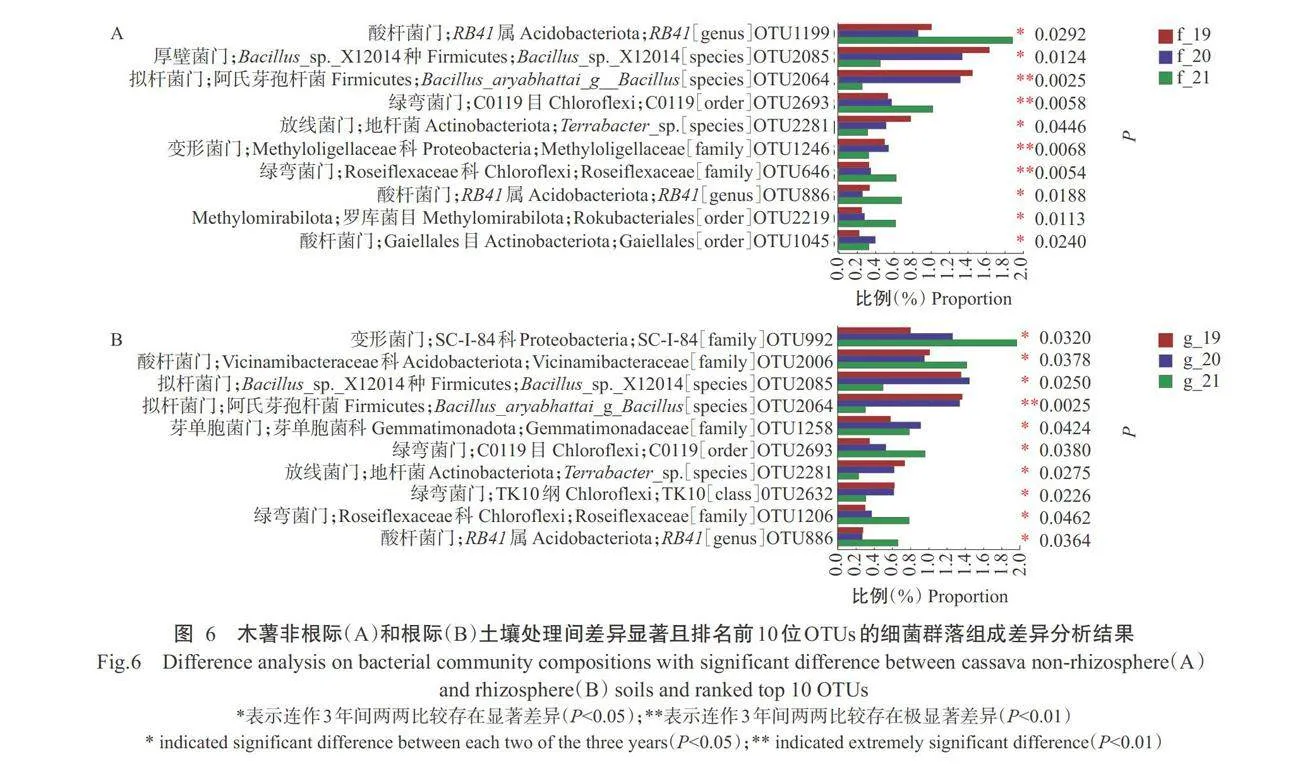
从所有样品的细菌群落中选取相对丰度均存在显著或极显著(Plt;0.01,下同)差异且排名前10位的 OTUs进行细菌群落组成差异分析,结果(图 6)表明,连作木薯非根际土壤细菌群落的前 10 位 OTUs 被注释为RB41属(OTU1199、OTU886)、Bacillus_sp._X12014种(OTU2085)、阿氏芽孢杆菌(OTU2064)、C0119目(OTU2693)、地杆菌(OTU2281)、Methyloligellaceae科(OTU1246)、Roseiflexaceae科(OTU646)、罗库菌目(OTU2219)和Gaiellales目(OTU1045),分别属于酸杆菌门、拟杆菌门、绿弯菌门、放线菌门和Methy-lomirabilota(图 6-A)。其中 OTU1199、OTU2693、OTU646、OTU886 和OTU2219的相对丰度均在连作第年升高,且与前2年相比存在显著或极显著差异,OTU2085、OTU2064、OTU2281和OTU1246 的相对丰度均在连作第3年降低,且与前2年相比存在显著或极显著差异。
从图6-B可看出,连作木薯根际土壤细菌群落的前10位OTUs被注释为SC-I-84科(OTU922)、Vic-inamibacteraceae科(OTU2006)、Bacillus_sp._X12014种(OTU2085)、阿氏芽孢杆菌(OTU2064)、芽单胞菌科(OTU1258)、C0119目(OTU2693)、地杆菌(OTU2281)、TK10纲(OTU2632)、Roseiflexaceae科(OTU1206)和RB41属(OTU646),分别属于变形菌门、酸杆菌门、拟杆菌门、芽单胞菌门和绿弯菌门。其中,OTU922、OTU2006、OTU2693、OTU1206和OTU886的相对丰度均在连作第3年升高,且与前2年相比存在显著差异,OTU2085、OTU2064、OTU2281和OTU2632的相对丰度均在连作第3年降低,且与前2年相比存在显著或极显著差异。可见,连作木薯非根际和根际土壤细菌群落中相对丰度均存在显著或极显著差异且排名前10位的0TUs大多在连作第3年出现显著或极显著变化。
2.3连作木薯对土壤养分与细菌群落相关关系的影响
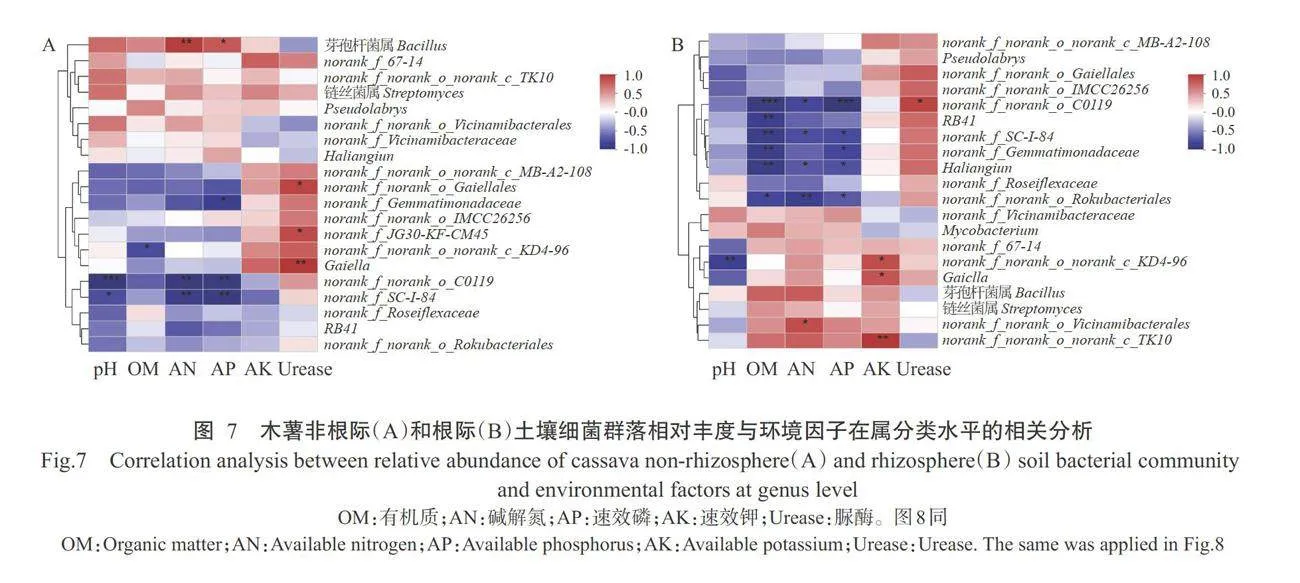
在非根际土中(图7-A),芽孢杆菌属(Bacillus)与碱解氮和速效磷含量,norank_f_norank_o_Gaiel-lales、norank f JG30-KF-CM45、Gaiella与脲酶活性,分别呈显著或极显著正相关。norank_f_Gemmati-monadaceae与速效磷含量,norank f_norank_onorank c KD4-96与有机质含量,norank f noranko_C0119和norank f SC-I-84与pH、碱解氮和速效磷含量,呈显著或极显著负相关;在根际土中(图7-B),norank f_norank o C0119与脲酶活性,norank fnorank_o_norank_c_KD4-96、Gaiella 和norank_fnorank o norank c TK10与速效钾含量,norank fnorank o Vicinamibacterales与碱解氮含量,呈显著或极显著正相关;RB41与有机质含量,norankf_Gemmatimonadaceae与有机质和速效磷含量,norank_fnorank_o_norank_c_KD4-96与pH,norank f_norank_o_C0119、norank_f_SC-I-84、Haliangium 和 norank_f_norank_o_Rokubacteriales 与有机质、碱解 氮和速效磷含量,呈显著或极显著负相关。可见,土 壤细菌群落组成同时受土壤理化性质和酶活性的影 响,在根际与非根际土环境中的影响程度存在差异。
从图8可看出,细菌群落丰度值最高的前5个细 菌门分别为变形菌门、酸杆菌门、绿弯菌门、厚壁菌 门和放线菌门,第一轴(RDA1)和第二轴(RDA2)的 解释变量分别为40.31%和9.99%,2个变量在坐标 对角线上把根际与非根际土壤样本按年际分成3个 群体,其中第3年样本与第1和第2年样本距离较远,说明木薯连作第3年开始对土壤理化性质及微生物群落产生明显影响。土壤pH、碱解氮含量、有机质含量、速效磷含量和脲酶活性是对细菌群落结构影响较大的环境因子,其中,脲酶活性与其他土壤理化性质指标均呈负相关。
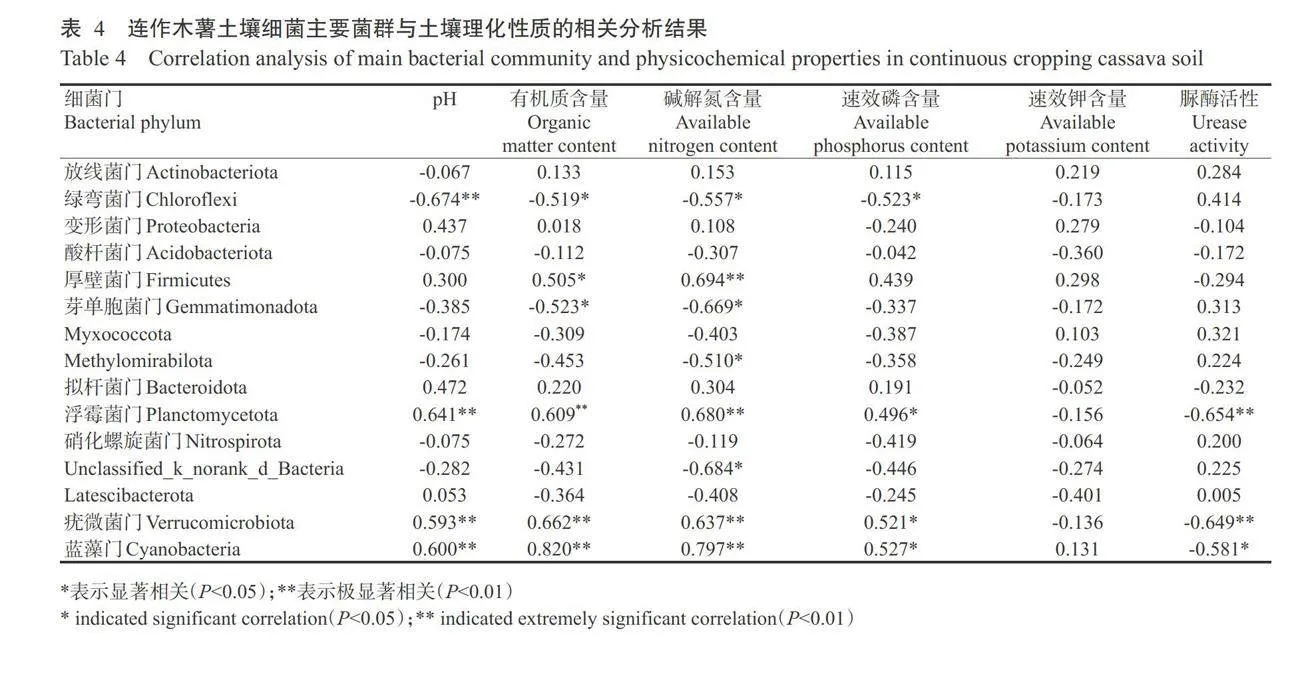
进一步选择门分类水平相对丰度大于0.10%的土壤细菌群落与理化性质进行相关分析,结果(表4)表明,有9个细菌门与土壤理化性质有显著或极显著相关性。其中绿弯菌门与土壤pH呈极显著负相关,与有机质、碱解氮和速效磷含量呈显著负相关;厚壁菌门与有机质含量呈显著正相关,与碱解氮含量呈极显著正相关;芽单胞菌门与有机质和碱解氮含量呈显著负相关;Methylomirabilota与碱解氮含量呈显著负相关;浮霉菌门与 pH.有机质和碱解氮含量呈极显著正相关,与速效磷含量呈显著正相关,与脲酶活性呈极显著负相关;Unclassified_k_norankd_Bacteria与碱解氮含量呈显著负相关;疣微菌门和蓝藻门与 pH、有机质和碱解氮含量呈极显著正相关,与速效磷含量呈显著正相关,与脲酶活性呈极显著或显著负相关。
综上所述,土壤的细菌群落分布与土壤环境因 子相互影响,对细菌群落影响较大的环境因子排序 为碱解氮含量gt;有机质含量gt;pHgt;速效磷含量gt;脲酶 活性。连作木薯会导致土壤pH及碱解氮、有机质和 速效磷含量降低,进而影响微生物群落演替,且连作 障碍在第3年开始显现。
3讨论
土壤理化性质是评价土壤优劣的重要指标。本 研究中,与头茬木薯相比,连作木薯的非根际土壤和根际土壤pH及有机质、碱解氮和速效磷含量均降低,其中,第2年非根际和根际土壤有机质含量急剧下降。前人研究表明,当自然生态系统受到人为干扰转换为农业生态系统后,由于大量地面裸露,导致土壤有机碳库在温带和热带地区分别降低60%和75%以上(陈虎等,2012;李娟等,2022)。本研究中,试验地位于亚热带地区,在种植木薯前为撂荒地,近似于自然生态系统。第1年种植木薯时,因前期除草效果较差,生育前期(4-6月)大量杂草覆盖地表,减少了地面裸露引起的土壤有机质消耗;块根膨大及成熟期(9—12月)遭遇长期干旱,导致木薯生长缓慢,块根平均产量低,木薯生长对土壤养分的消耗较小,且前期杂草、间苗秸秆及后期落叶还田对土壤有机质碳库有一定补充;收获期块根及茎秆生物量低,碳移除量较少,故第1年种植木薯后,土壤有机质还保留在10~20g/kg的水平(张童瑶等,2020;张叶叶等,2021)。而种植木薯的第2年,地块杂草相对较少,加上前期防控措施得当,自3月整地后,杂草更少,一直呈裸露状态,至7月后才开始封行,长期的土地裸露导致有机质下降(陈虎等,2012);块根膨大和成熟期雨量分布相对均匀,木薯整个生育期长势较好,块根平均产量提高,木薯生长对土壤养分消耗巨大,木薯和土壤微生物对土壤养分的竞争剧烈,进而促进微生物矿化分解有机质释放养分(渠晨晨等,2022);此外,第2年杂草较少,还田叶片也远少于移除的茎秆和块根,因此土壤有机质碳库一直处于亏损状态。另外,张叶叶等(2021)的研究表明,秸秆还田所引起的碳激发效应也可能造成土壤有机质下降;碳激发效应与土壤有机碳含量存在一定关系,当土壤有机碳含量在10~20g/kg时激发强度最高,高于或低于该阈值时激发变弱;长期单一作物秸秆还田不利于构建多样的微生物环境,易造成同一营养型微生物富集,对土壤有机碳造成长期损耗。本研究中,第1年收获木薯后的土壤有机质含量刚好处于最高激发强度区间,且第2年木薯长势较好,相对强大的根系所产生的根系分泌物也较多,加上连作还田的落叶,可能导致同一营养型微生物富集,进而对土壤有机碳造成损耗。此外,连作第2年土壤速效钾含量较前、后2年偏高,可能是因为第2年6月降雨量较大(为第1年和第3年6月降雨量的2倍),此时为木薯块根形成初期,根系活动旺盛,受 涝害胁迫会分泌大量有机酸和有机碳,在有机酸的 酸化作用下土壤速效钾含量缓慢提高,但因为作用 时间短且作用面积小,速效钾含量变化未达显著水 平,该结果与Henry等(2007)、何冰等(2015)的研究 结果一致。综上所述,土壤理化性质在连作木薯第 3年明显劣化,建议木薯连作不超过3年。该结果 也与梁海波等(2017)基于多年生产实际的预测结果 一致。
在健康的土壤系统中,植物会分泌可供微生物 利用的营养物质,同时有益微生物也会以各种方式 协助植物吸收土壤养分,如改变土壤养分有效性、形 成从枝菌根等,且有益微生物能同时协助植物抵御 病虫害和适应逆境(孙子欣和蔡柏岩,2022)。而连 作引起的土壤理化性质劣化和土壤微生物群落从 “有益菌”向“有害菌”改变是连作障碍形成的重要原 因(戴传超和张伟,2022)。在连作马铃薯(谢奎忠, 2021)、三七(刘雨艳等,2022)、切花菊(王坦,2022) 等作物中均发现根际分泌物会改变微生物群落多样 性。本研究结果表明,连作木薯根际与非根际土壤 细菌群落的Alpha多样性会逐年降低,但根际土壤细菌群落的Alpha多样性降低速度比非根际土壤慢。这可能是因为连作导致土壤养分下降,增强细菌群落之间的拮抗作用,进而导致群落稳定性下降;木薯根际分泌物中丰富的糖类和氨基酸能提供丰富的碳源和氮源,缓解根际土壤细菌群落对养分的竞争,故而保持较高的丰富度,从而导致非根际与根际土壤细菌群落结构的多样性和丰富度差异。
本研究中,不同连作年限木薯非根际与根际土壤中的优势菌门均包含放线菌门、变形菌门、绿弯菌门、拟杆菌门和酸杆菌门,与刘诗蓉等(2022)对半夏连作根际土细菌群落结构的研究结果相似。基于不同连作年限间有显著差异且最丰富的前 10个OTUs对比研究发现,非根际与根际土壤中共有的优势菌群为 Bacillus_sp._X12014 种、阿氏芽孢杆菌、地杆菌、C0119 目、RB41属和 Roseiflexaceae 科。Bacil-lus_sp._X12014种与阿氏芽孢杆菌均属于芽孢杆菌属,该菌属中有许多能降解土壤有毒物质的益生菌,能增强植物对外界环境的适应和抵抗能力,其中,阿氏芽孢杆菌具有促进植物吸收锌离子、防治植物病害、促进植物生长等作用(熊乙等,2018)。本研究发现,非根际与根际土壤的 Bacillus_sp._X12014种与阿氏芽孢杆菌的相对丰度均在连作第3 年大幅下降,导致土壤降解有毒物质的能力下降,这可能是引起木薯连作障碍的重要原因之一。地杆菌的相对丰度也在连作第3年显著下降,其在厌氧条件下的亚硝酸盐和硝酸盐转化为 NH的过程中扮演重要角色(田丽丽和王鑫,2020),这可能是引起土壤碱解氮含量随连作年限增加而下降的原因之一。C0119目与土壤CO,的相互作用可促进植物生长(Miao et al.,2021),而RB41被认为是土壤中参与碳循环的关键菌属(Stone et al.,2021),两者的相对丰度均在连作木薯第3年升高,但由于其相对丰度较小(lt;1.00%),有可能是由于相对丰度较高的芽孢杆菌属大幅下降引起的。此外,木薯根际土壤中特有的优势菌群有SC-I-84科、Vicinamibacteraceae科、芽单胞菌科和TK10纲。其中,SC-I-84被证实为作物根际土壤特有优势菌群或根系内生菌(庞师婵等,2020;肖健等,2022),其相对丰度随木薯连作年限逐年上升,这可能与某些木薯根系分泌物的逐年积累相关;而Vici-namibacteraceae科属酸杆菌门,其相对丰度随连作年限呈先降后升趋势,可能是由于连作第2年6月降雨过多使耕地受渍,甲烷菌大量繁殖,进而导致第2年Vicinamibacteraceae科相对丰度下降。综上所述,木薯非根际与根际土壤中相对丰度较高的细菌OTUs均在连作第3年出现显著的相对丰度变化,其中Bacillus_sp._X12014、阿氏芽孢杆菌和地杆菌等有益菌丰度大幅下降,这可能是木薯连作障碍在连续种植第3年后开始显现的重要原因之一,也直接印证了梁海波等(2017)的模型预测结果。此外,本研究还发现,在连作木薯非根际与根际土壤中,pH、有机质含量、碱解氮含量、速效磷含量、速效钾含量和脲酶活性对土壤细菌群落结构有显著影响,但不 同环境因子对土壤细菌群落的影响也不同,表明在 不同生境中,土壤细菌对环境因子的响应存在差异。
4结论
连作会导致木薯非根际和根际土壤理化性质、 细菌群落多样性和丰富度发生变化,土壤理化性质 和细菌群落结构均在连作第3年出现明显劣化。土 壤理化性质变化与根际土壤细菌群落结构的差异存 在一定相关性,有益菌丰度降低可能是木薯连作造 成土壤细菌群落劣化演替的重要原因之一。
参考文献(References):
陈福慧,申乃坤,姜明国,王一兵.2022.作物重茬连作障碍中 自毒物质的研究进展[J].中国农业科技导报,24(10): 125-132. [Chen F H, Shen N K, Jiang M G, Wang Y B. 2022. Research progress of autotoxic secretions in crops replant successive cropping obstacles [J]. Journal of Agri- cultural Science and Technology, 24 (10) : 125-132.] doi: 10.13304/j.nykjdb.2021.0889.
陈虎,王嘉学,胡灿灿,马照伦.2012.滇东喀斯特地区石漠化 裸露红土的表层有机质含量分析[J].中国岩溶,31(4): 423-425. [Chen H, Wang J X, Hu C C, Ma Z L. 2012. Organic matter content of the bare lateritic surface-layer in east Yunnan karst rocky deserts [J]. Carsologica Sinica,31(4):423-425.] doi:10.3969/j.issn.1001-4810.2012.04.011.
戴传超,张伟.2022.有益微生物缓解花生连作障碍机理研究 进展[J].微生物学杂志,42(1):1-8.[DaiCC,Zhang W.
2022. Research progress on the beneficial microbe-mediated alleviation of peanut continuous cropping obstacle[J]. Journal of Microbiology,42(1):1-8.] doi:10.3969/j.issn.1005-7021.2022.01.001.
何冰,薛刚,张小全,许晓敬,姚健,杨铁钊.2015.有机酸对土 壤钾素活化过程的化学分析[J].土壤,47(1):74-79.[He B, Xue G, Zhang X Q, Xu X J, Yao J, Yang T Z. 2015. Analysis on chemical mechanism of potassium release pro- cess from soil as influenced by organic acids[J]. Soils,47(1):74-79.] doi:10.13758/j.cnki.tr.2015.01.012.
侯乾,王万兴,李广存,熊兴耀.2019.马铃薯连作障碍研究进 展[J].作物杂志,(6):1-7.[Hou Q,Wang WX,Li G C, Xiong X Y. 2019. Advances in the research on potato con- tinuous cropping obstacles [J]. Crops, (6) : 1-7.] doi: 10.16035/j.issn.1001-7283.2019.06.001.
李娟,魏甲彬,杨宁.2022.土地利用变化对土壤有机质影响 的研究进展及展望[J].湖南生态科学学报,9(3):106-111.李军,魏建波,杨。土地利用变化对土壤有机质影响的研究进展与展望[J].湖南生态学报,9 (3) : 106-111。」doi:10.3969/j . ISSN . 2095-7300 . 2022 . 03 . 014。
梁海波,黄洁,魏云霞,陆昆典,刘翠娟.2017.基于农户尺度 的木薯产量差及生产限制因素分析[J].云南农业大学学报(自然科学),32(6):975-984.[Liang H B,HuangJ,Wei Y X, Lu K D,Liu C J. 2017. Analysis of yield gap andlimiting factors for cassava on the farmland[J]. Journal ofYunnan Agricultural University,32(6) : 975-984.] doi: 10.16211/j.issn.1004-390X(n).2017.06.003.
刘珊廷.2020.木薯连作与轮作对土壤理化性状及微生物群落和产量的影响[D].南宁:广西大学.[LiuST.2020.Causes and mechanism on crop continuous monocultureproblems and its control strategy [D]. Nanning: GuangxiUniversity.] doi:10.27034/d.cnki.ggxiu.2020.000417.
刘诗蓉,王红兰,孙辉,杨萍,蒋舜媛,蒋桂华.2022.半夏连作对根际土壤微生物群落的影响研究[J].中草药,53(4):1148-1155. [Liu S R, Wang H L, Sun H, Yang P, Jiang SY, Jiang G H. 2022. Effects of continuous cropping ofPinellia ternata on rhizospheric microbial community [J].Chinese Traditional and Herbal Drugs, 53 (4) : 1148-1155.]doi: 10.7501/j.issn.0253-2670.2022.04.023.
刘雨艳,保丽美,魏云林,字富庭,谭勇.2022.连作三七根际土壤自毒物质与微生物互作研究进展[J].中药材,45(1):242-247.[Liu Y Y,Bao L M,Wei Y L,Zi F T,Tan Y.2022. Research progress on interaction between autotoxicsubstances and microorganisms in rhizosphere soil ofPanax notoginseng in continuous cropping[J]. Journal ofChinese Medicinal Materials , 45 (1) : 242-247. ] doi : 10.13863/j.issn1001-4454.2022.01.043.
刘株秀,刘俊杰,徐艳霞,张武,米刚,姚钦,王光华.2019.不 同大豆连作年限对黑土细菌群落结构的影响[J].生态学报,39(12):4337-4346.[Liu ZX,Liu JJ,XuYX,Zhang W, Mi G, Yao Q, Wang G H. 2019. Effects of continuous cropping years of soybean on the bacterial community structure inblack soil [J]. Acta Ecologica Sinica, 39 (12) :
4337-4346.] doi:10.5846/stxb201801270212.
卢孟召,刘梅,陈光,王雪峰.2022.灵芝连作对土壤理化性质 及线虫群落的影响[J].吉林农业大学学报,44(5):586- 594. [Lu M Z, Liu M, Chen G, Wang X F. 2022. Effect of biochar-based fertilizer on soil bacteria and fungi quantity and community structure in acidified tea garden [J]. Jour- nal of Jilin Agricultural University, 44 (5) : 586-594.] doi: 10.13327/j.jjlau.2022.1487.
庞师婵,郭霜,任奎瑜,王帅帅,杨尚东.2020.番茄/茄子嫁接 对其根际土壤生物学性状及细菌群落结构的影响[J].园艺学报,47(2):253-263.[Pang S C,Guo S,Ren KY, Wang S S, Yang S D. 2020. Impact of grafting on soil microbial properties and bacterial community structure in tomato rhizosphere [J]. Acta Horticulturae Sinica, 47 (2) :253-263.] doi: 10.16420/j.issn.0513-353x.2019-0444.
覃锋燕,杨慰贤,彭晓辉,黎亮武,阳太亿,周佳,韦茂贵,申章佑.2022.粉垄耕作木薯根际与非根际土壤的细菌群落结构多样性差异[J].西南农业学报,35(4):729-739.[QinF Y,Yang W X,Peng X H,Li L W,Yang T Y,Zhou J,WeiM G, Shen Z Y. 2022. Difference in the diversity of bacte-rial community structure in rhizosphere and non-rhizospheresoil of cassava in Fenlong tillage[J]. Southwest China Jour-nal of Agricultural Sciences, 35 (4) : 729-739.] doi: 10.16213/j.cnki.scjas.2022.4.001.
渠晨晨,任稳燕,李秀秀,蔡鹏,陈雯莉,黄巧云.2022.重新认识土壤有机质[J].科学通报,67(10):913-923.[QuCC, Ren W Y, Li X X, Cai P, Chen W L, Huang Q Y. 2022. Revisit soil organic matter[J]. Chinese Science Bulletin,67(10):913-923.] doi:10.1360/TB-2021-0704.
宋秀丽,吴舒婷,李锦辉,杨锡朗,周芮,况丹妮,李警督,于海 涛,杨德光.2022.土壤微生物群落对连作种植制度的响 应[J].玉米科学,30(1):172-181.[Song XL,Wu S T,Li J H,Yang XL,Zhou R,Kuang D N,Li J D,Yu H T,Yang D G. 2022. Response of soil microbial community to con- tinuous cropping system[J]. Journal of Maize Sciences,30(1) : 172-181. ] doi : 10.13597/j.cnki.maize.science.20220124.
孙子欣,蔡柏岩.2022.连作对土壤微生物菌群影响及修复研 究进展[J].作物杂志,(6):7-13.[SunZX,Cai B Y.2022.Research advances on the effects of continuous cropping on soil microbial community and restoration techniques [J]. Crops, (6):7-13.] doi:10.16035/j.issn.1001-7283.2022.06.002.
田丽丽,王鑫.2020.地杆菌属微生物在厌氧污水处理中的应 用潜力[J].给水排水,56(S2):201-206.[Tian LL,Wang X. 2020. Potential applications of Geobacterin anaerobic wastewater treatment[J]. Water amp; Wastewater Engineering, 56 (S2) : 201-206. ] doi : 10.13789/j.cnki. wwe1964.2020.S2.030
王坦.2022.连作下切花菊根际微生物群落组成和功能的动 态变化[D].武汉:华中农业大学.[Wang T.2022.Dynamic changes of rhizosphere microbial community composition and function in cut chrysanthemum under continuous crop- ping[D]. Wuhan: Huazhong Agricultural University.] doi: 10.27158/d.cnki.ghznu.2022.000706.
王晓琪,姜伟涛,姚媛媛,尹承苗,陈学森,毛志泉.2020.苹果 连作障碍土壤微生物的研究进展[J].园艺学报,47(11): 2223-2237. [Wang X Q, Jiang W T, Yao Y Y, Yin C M, Chen X S, Mao Z Q. 2020. Research advance of apple replant disease based on soil microorganism[J]. Acta Hor- ticulturae Sinica,47(11):2223-2237.] doi: 10.16420/j.issn.0513-353x.2019-0941.
王战.2016.木薯磷营养与连作障碍的关系研究[D].南宁:广西大学.[Wang Z.2016.Study on the relationship be- tween phosphorus nutrition and continuous cropping obstacle in cassava [D]. Nanning:Guangxi University.]
翁佩莹,郑红艳.2020.作物连作障碍的成因与机制及其消减策略[J].亚热带植物科学,49(2):157-162.[Weng PY,Zheng H Y. 2020. Causes and mechanism on crop conti-nuous monoculture problems and its control strategy [J].Subtropical Plant Science, 49 (2) : 157-162.] doi: 10.3969/j.issn.2095-1191.2021.06.005.
肖健,梁阗,杨尚东,谭宏伟.2022.甘蔗植株茎部内生细菌群落组成对不同配方缓释肥的响应[J].热带作物学报,44(3):614-627. [Xiao J, Liang T, Yang S D, Tan H W. 2022.
Response of endophytic bacterial community compositions in stems of sugarcanes under different slow-release fertilizers applications[J]. Chinese Journal of Tropical Crops,44
(3):614-627.] doi:10.3969/j.issn.1000-2561.2023.03.019.
谢奎忠.2021.连作马铃薯根系分泌物介导的枯萎病发生机 制及根际互作[D].兰州:甘肃农业大学.[XieKZ.2021.
Mechanism of Fusarium wilt and rhizosphere interaction mediated by root exudates in continuous cropping potato [D]. Lanzhou: Gansu Agricultural University.] doi: 10.27025/d.cnki.ggsnu.2021.000041.
熊乙,欧翔,贾蓉,玉柱,许庆方.2018.阿氏芽孢杆菌应用研 究进展[J].生物技术,28(3):302-306.[Xiong Y,Ou X, Jia R, Yu Z, Xu Q F. 2018. Research progress in applica- tion of Bacillus aryabhattai [J]. Biotechnology, 28 (3) :302-306.] doi:10.16519/j.cnki.1004-311x.2018.03.0052.
杨浩娜,周成言,邬腊梅,王立峰,李祖任.2022.植物化感物 质的作用机理研究进展[J].湖南农业科学,(3):108-112.[Yang H N,Zhou C Y,Wu L M,Wang L F,Li Z R. 2022. Research progress of plant allelochemicals mechanism[J].Hunan Agricultural Sciences, (3): 108-112.] doi:10.16498/ j.cnki.hnnykx.2022.003.025.
张童瑶,胡月明,任向宁,陈飞香,冯雪珂.2020.弃耕行为对 亚热带农田土壤有机质时空变化作用机理研究[J].农业资源与环境学报,37(6):805-817.[ZhangTY,Hu Y M, Ren X N,Chen F X,Feng X K. 2020. Study on the spatio- temporal variation of soil organic matter induced by aban- doned tillage behavior[J]. Journal of Agricultural Resour- ces and Environment, 37 (6) : 805-817.] doi: 10.13254/j.jare.2020.0480.
张叶叶,莫非,韩娟,温晓霞,廖允成.2021.秸秆还田下土壤 有机质激发效应研究进展[J].土壤学报,58(6):1381- 1392. [Zhang Y Y, Mo F, Han J, Wen X X, Liao Y C.2021. Research progress on the native soil carbon priming after straw addition [J]. Acta Pedologica Sinica, 58 (6) :1381-1392.] doi:10.11766/trxb202006260259.
周贵靖.2017.木薯根际土壤理化性状与连作障碍关系研究[D].南宁:广西大学.[Zhou G J.2017.The relationship between the continous cropping obstacle and physical and chemical properties of cassava rhizosphere soil abstract[D]. Nanning:Guangxi University.]
Henry A, Doucette W, Norton J, Bugbee B. 2007. Changes in crested wheatgrass root exudation caused by flood,drought, and nutrient stress[J]. Journal of Environmental Quality,36(3):904-912. doi: 10.2134/jeq2006.0425sc.
Hontoria C, Garcia-Gonzalez I, Quemada M, Roldan A, Alguacil M M. 2019. The cover crop determines the AMF community composition in soil and in roots of maize after a ten-year continuous crop rotation [J]. Science of the Total Environment, 660: 913-922. doi: 10.1016/j.scitotenv.2019.01.095.
Miao S J, Zhao H F, Pan S Q, Zhang N, Qiao Y F. 2021. Elevated CO2 weakens the shift in bacterial community structure in response to 8-year soybean straw return in the same experiment [J]. International Journal of Phytoreme-diation, 23 (5): 505-510. doi: 10.1080/15226514.2020.1825332.
Stone B W, Li J H,Koch B J, Blazewicz S J,Dijkstra P, Hayer M, Hofmockel K S, Liu X A, Mau R L, Morrissey E M, Pett-Ridge J, Schwartz E, Hungate B A. 2021. Nutrients cause consolidation of soil carbon flux to small proportion of bacterial community[J]. Nature Communications, 12
(1):3381. doi:10.1038/s41467-021-23676-x.
(责任编辑王晖)
猜你喜欢
传奇·传记文学选刊(2022年8期)2022-05-30 10:48:04
世界热带农业信息(2019年7期)2019-08-30 01:02:52
世界热带农业信息(2018年11期)2018-01-17 10:08:19
广东第二课堂·小学(2017年5期)2017-05-27 20:09:13
江苏农业科学(2016年12期)2017-04-05 14:33:56
现代农业科技(2016年20期)2016-12-20 08:54:23
江苏农业科学(2016年7期)2016-10-20 15:04:28
热带农业科学(2015年12期)2016-01-15 05:02:36
江苏农业科学(2015年2期)2015-03-12 18:18:10
世界热带农业信息(2014年8期)2014-09-23 18:09:39
