
蓝膜对西南杨幼苗光合参数及苗木生长的影响
2024-08-04 00:00:00陈智华刘金萍马少杰张晓琳王静敏奎梦漪何承忠纵丹
云南农业大学学报(自然科学) 2024年3期







摘要: 【目的】阐明蓝膜下光质成分对西南杨(Populus schneideri) 幼苗生长的影响。【方法】选取1 年生西南杨扦插幼苗置于蓝色、白色薄膜制成的拱棚下培育60 d,测定其生长指标、叶片解剖结构、光合参数、光合色素及糖含量。【结果】蓝膜能够促进西南杨幼苗的苗高生长及叶长、叶宽的增加,其苗高、叶长和叶宽分别比白膜增加36.36%、418.52% 和495.24%,且两者间差异极显著。与白膜相比,蓝膜处理的叶片上表皮显著变薄;而叶片、栅栏组织、海绵组织和下表皮的厚度仅略微增加。蓝膜处理下,光合气体交换参数蒸腾速率(Tr)、净光合速率(Pn)、胞间CO2 浓度(Ci) 和气孔导度(Gs) 均显著或极显著增加,表明蓝膜处理能够提高西南杨幼苗的光合效率。蓝膜处理的表观量子效率、最大净光合速率、羧化效率和光呼吸速率均极显著高于白膜,而CO2 饱和点和CO2 补偿点均显著或极显著小于白膜,表明蓝膜处理使得西南杨幼苗对弱光有较好的适应性,但对高浓度CO2 环境适应能力较弱。蓝膜处理的叶绿素a、叶绿素b、淀粉和还原性糖含量分别比白膜高22.69%、41.18%、65.85% 和54.26%,而蔗糖含量显著低于白膜。【结论】西南杨幼苗对高比例蓝光有较强的适应性,蓝膜可促进西南杨幼苗的苗高生长,并通过增加叶片Gs、叶绿素含量、RubisCO 活性及含量等促进植株的光合作用。研究结果为解析西南杨适应高比例短波光的机制提供了研究基础,也为西南杨引种栽培光质条件的选择与优化提供了科学依据。
关键词: 蓝膜;光合作用;西南杨;叶片解剖结构;红光;蓝光
中图分类号: S792.113.01 文献标志码: A 文章编号: 1004–390X (2024) 03−0152−11
光是调控植物生长发育的重要环境因子之一,其中光质在植物生长发育、气孔开度、光合色素含量、光合作用、基因表达等方面具有重要的调控作用[1-2]。红光和蓝光是植物生长和发育所必需的光质[3],红光可促进叶面积增加、茎的伸长和干物质积累;蓝光则主要参与叶绿素生物合成、气孔开闭和植物光形态建成[4],但抑制叶面积增加和茎的伸长,使植物株型更加矮小紧凑[5]。人工改良环境中的光质比例技术(如有色薄膜覆盖、LEDs 电照等) 可有效改善植物生长发育及形态建成,具有良好的实用性和可操作性。有色薄膜覆盖技术是一种比LEDs 电照技术更经济实惠的方法,常用于花卉、蔬菜及部分林木植物培育,但在林木育种中的运用研究相对较少[6]。前人研究表明:有色薄膜覆盖技术能显著改善心里美萝卜、甜瓜、青花菜、越橘、春美桃等作物的产量和品质[7-11],但同一光质对作物的影响结果不完全相同甚至有很大差异,说明有色薄膜覆盖技术的培育效果在不同环境及物种中可能存在较大差异。因此,有色薄膜覆盖技术的效果还需要进一步根据植物种类、生长环境及处理方式进行具体探究。
杨树(Populus spp.) 是杨柳科(Salicaceae) 杨属(Populus) 树种的统称,其生长速度快、成材时间短,被广泛用于绿化造林、工业用材等。西南杨(P. schneideri) 为杨属青杨派(Tacamahaca) 树种,是分布于中国西南地区海拔3 000 m 以上的乡土树种,具有良好的速生性和抗逆性,对西南地区复杂多样的气候和地理环境有较强的适应性[12]。近年来,杨树作为主要的经济兼绿化树种在云南省昆明市大规模引进种植。为丰富昆明的杨树资源、实现杨树的科学引种和推广栽培,课题组在前期研究中将西南杨引种至昆明,并对比分析证明不同杨树树种间生长量及光合特性存在较大差异[13]。以昆明本土杨树滇杨(P. yunnanensis) 为对照,发现西南杨生长速度低于滇杨。由于西南杨原生地海拔高于昆明,随着海拔的降低,自然光照中蓝光比例减少可能是导致西南杨生长速度减慢的关键原因之一。为模拟高海拔地区高蓝光比例,进而提高昆明地区西南杨的生长速度,本研究采用蓝膜覆盖技术对西南杨幼苗进行培育,以白膜为对照,分析不同颜色覆膜下西南杨幼苗的生长指标、叶片解剖结构、光合参数、光合色素及产物含量,研究结果可为解析西南杨适应高比例短波光的机制提供研究基础,也可为西南杨引种栽培光质条件的选择与优化提供科学依据。
1 材料与方法
1.1 试验材料
选取1 年生健康无病虫害的枝条为材料,截取长为25 cm 的枝条作为插穗,经12 h 浸水处理3 个无性系后,各挑选15 根插穗分别扦插至育苗盆中(口径为42 cm),每盆扦插1 根插穗,并置于智能温室大棚内培育,成活后仅保留1 根主枝,修剪其余枝条。于2020 年6 月下旬选择长势一致且生长状况良好的苗木移至覆有蓝膜和白膜的拱棚内。
1.2 试验方法
试验拱棚位于西南林业大学树木园内(25°03′74″N,102°45′03″E,海拔1 954 m),由东向西排列,覆盖蓝膜和白膜的拱棚各搭建1 个,拱棚长×宽×高为3 m×3 m×2 m,间距2 m,覆盖白膜的拱棚为对照。每个棚内放置3 个无性系,1 个无性系为1 个生物学重复,每个无性系放置3 盆插穗,每盆相距20 cm,试验期间定期浇水、除草、抹除插穗萌芽。培育60 d 后测量其生长及光合参数等指标。
1.2.1 薄膜透射光谱测定
在 8: 0 0—18: 0 0 间每隔 2 h 用 HiPoint HR-550 植物智慧光谱仪对拱棚内的透射光谱成分进行测定。每个时间点测量3 次,每个拱棚取其日均值作为透射光谱比率的代表值。
1.2.2 生长指标测定
于2020 年7 月和9 月,采用卷尺和游标卡尺分别测定蓝膜、白膜下西南杨幼苗的生长指标,以主枝长度作为苗高、主枝基部2 cm 处的直径作为地径。每种处理下每个无性系测定3 株苗木,取其平均值作为1 次重复,共测定3 个生物学重复,计算苗高及地径的绝对生长量。
1.2.3 叶片组织结构测定
参考周乃富等[14]的方法。每个无性系选取1 株,每个处理共选择3 株,选其主枝自上而下的第5 片叶,切取2 cm×2 cm 组织制作成厚度为10 μm 的切片,经番红—固绿染色后用OlympusBH-2 型光学显微镜观察。用Motic DigiL abll 软件测定叶片厚度(leaf thickness,LT)、栅栏组织厚度(palisade tissue thickness,PT)、海绵组织厚度(spongy tissue thickness,ST)、上表皮厚度(upperepidermis thickness,UE)、下表皮厚度(lowerepidermis thickness,LE) 等叶片组织结构参数。每个样本观测10 个视野,取其平均值,并根据公式计算叶片组织结构紧密度 (cell tense ratio,CTR)、叶片组织疏松度 (sponge ratio,SR) 以及栅栏组织和海绵组织厚度的比值(R),计算公式分别为:CTR=(PT/LT)×100%;SR=(ST/LT)×100%;R=PT/ST。
1.2.4 光合参数测定
光合参数采用 LI-6 800 便携式光合测定仪测定。光合日变化的测定于2020 年9 月中下旬无云晴天进行,选取苗木主枝从上至下的第5~7 片叶,在 8: 0 0—18: 0 0 间每隔 2 h 分别测定蓝膜、白膜拱棚下苗木的光合气体交换参数及环境因子参数。每株测定3 片叶,每个无性系测定3 株作为3 次生物学重复。
在晴天 9:0 0—11:0 0 对西南杨幼苗从上至下的第5 片叶的Pn-PAR 响应曲线和 Pn-CO2 响应曲线进行测定。每个无性系测定1 株作为3 次生物学重复。测定 Pn-PAR 响应曲线时,叶室 CO2 设定为 400 μmol/mol,使用内置红蓝光将光合有效辐射(photosynthetically available radiation,PAR)依次设置为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、300、200、150、100、50、30 和0 μmol/(m2·s)[15];测定 Pn-CO2 响应曲线时,PAR 设置为1 800 μmol/(m2·s),CO2 梯度设置为400、300、200、100、50、20、10、400、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 和2 200 μmol/mol[15]。
1.2.5 光合色素及糖类含量测定
采集自上向下第 5~7 片新鲜叶片,采用乙醇研磨提取法提取光合色素,利用可见光分光光度仪分别在波长 665、649 和470 nm 下测定吸光度值 (A665、A649 和 A470),每个无性系采集1 株作为1 个生物学重复,每个样品测定3 次并取其平均值作为1 次重复,每个处理设置3 次生物学重复。根据公式计算叶绿素 a (Chla)、叶绿素 b (Chlb)和类胡萝卜素(Car) 的含量[16]:Chla=[(13.95A665−6.88A649)×提取液总量 ]/(样品鲜质量×1 000);Chlb=[(24.96A649−7.32A665)×提取液总量 ]/(样品鲜质量×1 000);Car={[1 000A470+3.27×(13.95A665−6.88A649)+104×(24.96A649−7.32A665)]/299×提取液总量}/(样品鲜质量×1 000)。
采用蒽酮—H2SO4 法和3,5-二硝基水杨酸法[17]分别制备可溶性总糖、还原糖及淀粉提取液;利用可见光分光光度计测定吸光度值,根据公式计算叶片的糖类含量[17]:
还原糖含量= (C×Vt)=(W×Vs)×100%;
可溶性糖含量= (C×Vt×n)=(W×Vs×1000)×100%;
淀粉含量= (C×Vt×0:9)=(W×Vs×1000)×100%;
蔗糖含量= (可溶性糖含量-还原糖含量)×0:95。
式中:C 为利用葡萄糖标准曲线计算出样品中葡萄糖的含量,mg;Vt 为样品提取液总体积,mL;W 为样品干质量,mg;Vs 为测定时所用样品体积,mL;n 为稀释倍数;1 000 为换算系数;0.9为葡萄糖换算成淀粉的系数。每个样品测定3 次,取平均值作为1 次重复,每个处理设置3 个生物学重复。
1.3 数据统计与分析
利用Excel 2019 和 SPSS 21.0 整理并分析数据,差异性分析选用独立样本t 检验;利用GraphPadPrism 8.0 绘图。选用非直角双曲线模型、直角双曲线模型分别对 Pn-PAR 响应曲线、Pn-CO2响应曲线测定的数据进行拟合[18-20]。相关性分析以2 种膜下各测量指标的均值作为代表值计算指标间的Person 相关系数。
2 结果与分析
2.1 薄膜透射光谱
由图1 可知:2 种膜对光质的透过程度不同,光质成分存在显著或极显著差异。白膜下,紫外光比率(2.13%) 显著高于蓝膜(1.60%),绿光(24.28%) 和红光比率(28.19%) 极显著高于蓝膜(19.66%、10.51%),蓝光(15.68%) 和远红光比率(29.72%) 极显著低于蓝膜(33.76%、34.47%)。
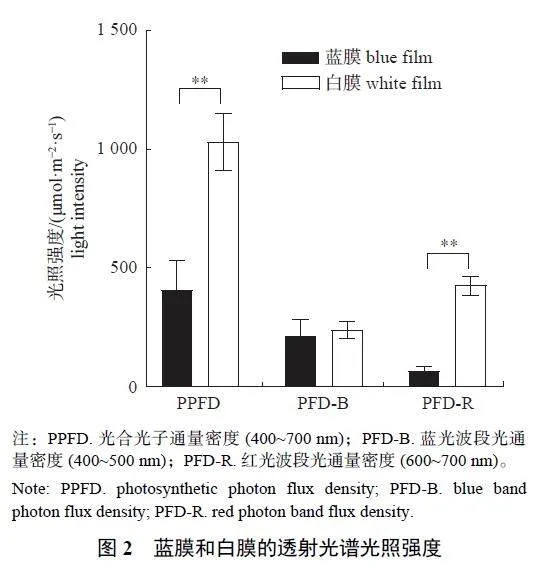
由图2 可知:与白膜相比,蓝膜的光合光子通量密度(photosynthetic photon flux density,PPFD)和红光波段光通量密度(red photon band fluxdensity,PFD-R) 均极显著降低,但两者蓝光波段光通量密度(blue band photon flux density,PFDB)无显著差异。表明2 种膜在蓝光范围的光照强度基本一致,但蓝膜红光范围的光照强度远低于白膜,即蓝膜能够过滤部分红光,使膜内蓝光[PFD-B 215.35 μmol/(m2·s)] 与红光[PFD-R66.35 μmol/(m2·s)] 的光照强度比例约为3∶1。
2.2 蓝膜对西南杨幼苗生长的影响
由表1 可知:蓝膜处理下西南杨幼苗的苗高绝对生长量极显著高于白膜处理,但两者的地径绝对生长量无显著差异;蓝膜处理下西南杨幼苗叶长和叶宽均极显著高于白膜,但两者的叶柄长无显著差异。说明蓝膜处理能够促进西南杨幼苗的苗高生长与叶长、叶宽的增长。
2.3 蓝膜对西南杨幼苗叶片结构的影响
由表2 可知:蓝膜处理下西南杨幼苗叶片上表皮厚度显著低于白膜处理,但2 种处理的其他叶片解剖结构指标无显著差异。说明蓝膜处理能使叶片上表皮变薄,但对叶片下表皮厚度及组织结构构成无显著影响。
2.4 蓝膜对西南杨幼苗光合参数的影响
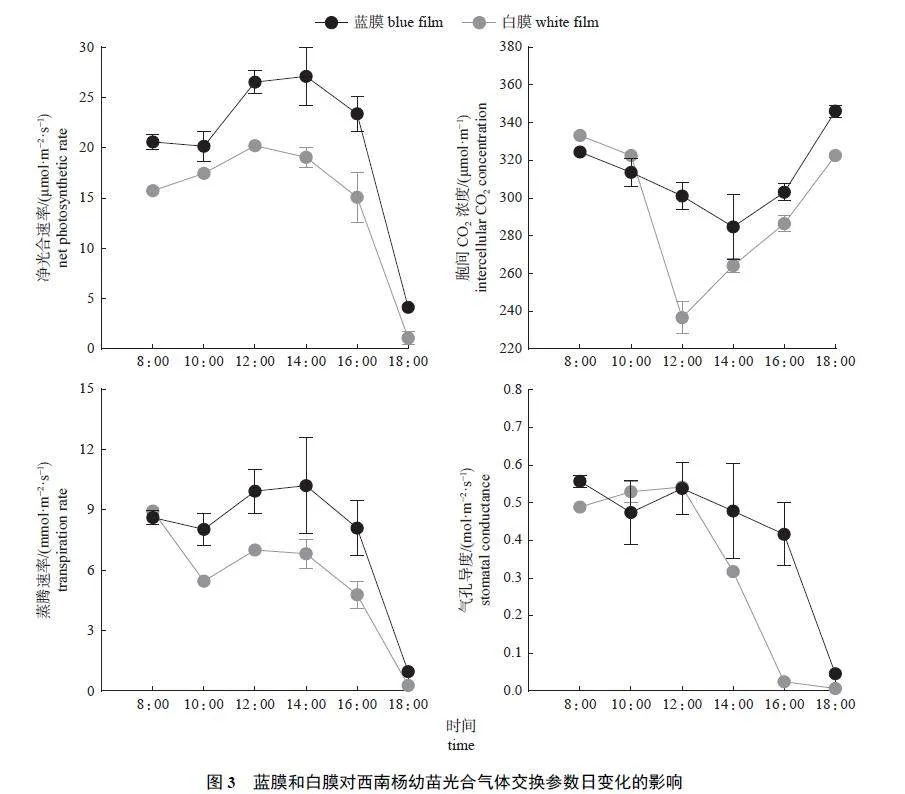
由图3 可知:2 种颜色薄膜处理下,西南杨幼苗的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci) 和气孔导度(Gs) 的变化趋势基本一致。蓝膜处理的 Pn、Tr 峰值出现在 14:0 0,白膜处理的 Pn 峰值出现在 12:0 0;2 种处理的 Gs 峰值均出现在 12:0 0;蓝膜处理的 Ci 在 14:0 0 出现峰谷,白膜处理的 Ci 在 12:0 0 出现峰谷。此外,日均值比较结果(表3) 显示:蓝膜处理下Pn、Tr、Ci 的日均值极显著高于白膜,Gs 显著高于白膜。
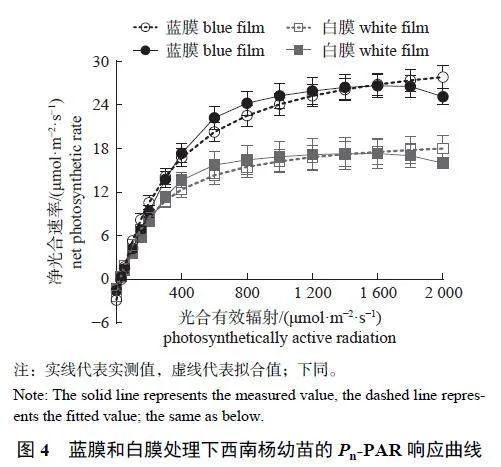
Pn-PAR 曲线(图4) 显示:蓝膜(R2=0.997) 与白膜(R2=0.998) 的曲线拟合效果均较好,当PAR小于200 μmol/(m2·s) 时,Pn 随着PAR 值的增加呈线性增加,达到光饱和点后,上升趋势逐渐平缓。不同处理下西南杨幼苗Pn-PAR 曲线特征参数比较结果(表4) 显示:蓝膜下的表观量子效率(apparent quantum efficiency,AQE) 和光响应最大净光合速率(LPnmax) 极显著高于白膜;而两者的暗呼吸速率(Rd)、光饱和点(light saturation point,LSP) 和光补偿点(light compensation point,LCP)无显著差异。
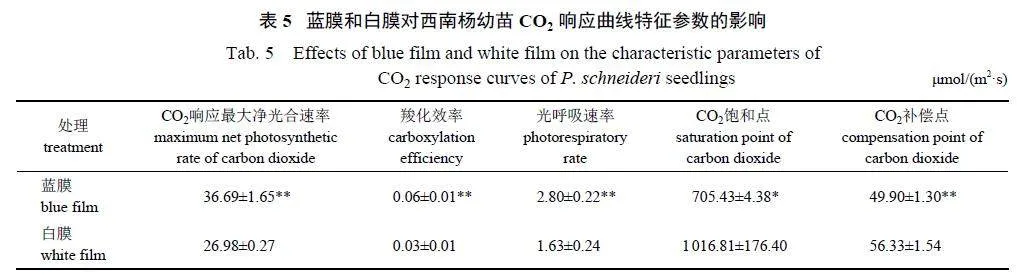
Pn-CO2 曲线(图5) 显示:蓝膜(R2=0.963) 的曲线拟合程度不及白膜(R2=0.992)。Pn 随着CO2浓度的增加而呈线性升高,出现CO2 饱和点后,Pn 的变化曲线逐渐趋于平缓。蓝膜处理下,西南杨幼苗叶片的CO2 最大净光合速率(CPnmax)、羧化效率(α) 和光呼吸速率(Rp) 均极显著大于白膜处理,CO2 饱和点(saturation point of CO2,CSP)显著小于白膜处理,CO2 补偿点(compensation pointof CO2,CCP) 极显著小于白膜处理(表5)。说明蓝膜处理能够通过促进西南杨幼苗的气体交换速率及光辐射的利用能力,提高其光合作用速率,且蓝膜处理能够使西南杨具有较强的弱光适应性,但对高浓度CO2 的适应能力较弱。
2.5 蓝膜对西南杨幼苗叶片光合色素和糖含量的影响
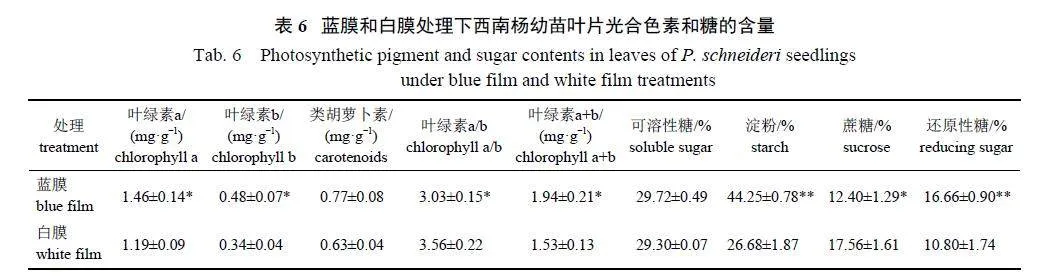
由表6 可知:蓝膜处理下,西南杨幼苗叶片叶绿素a、叶绿素b 和叶绿素a+b 含量显著高于白膜处理,叶绿素a/b 显著低于白膜处理,而类胡萝卜素含量在两者间无显著差异;蓝膜处理下,淀粉和还原性糖含量显著高于白膜处理,而蔗糖含量显著低于白膜处理。说明蓝膜处理能提高西南杨幼苗叶片中叶绿素a 和叶绿素b 的含量,并且能促进淀粉和还原性糖的积累。
2.6 西南杨幼苗各指标与不同光质的相关性
由表7 可知:蓝光和远红光与苗高、叶长、叶宽呈极显著正相关,而紫外光、绿光和红光与上述指标呈极显著负相关;紫外光与地径呈显著正相关,远红光与地径呈显著负相关。蓝光和远红光与上表皮厚度呈显著负相关,紫外光、绿光和红光与其呈显著正相关。蓝光和远红光与AQE、LPnmax、CPnmax、α、Rp 呈极显著正相关,而紫外光、绿光和红光与上述指标呈极显著负相关;蓝光和远红光与CSP 呈显著负相关,而紫外光、绿光和红光与之呈显著正相关;蓝光和远红光与CCP 呈极显著负相关,而紫外光、绿光和红光与之呈极显著正相关。蓝光和远红光与可溶性糖、淀粉、蔗糖呈极显著正相关,紫外光、绿光和红光与上述指标呈极显著负相关。
3 讨论
3.1 光质对西南杨生长的影响
光质对植物的生长、发育具有重要意义,红光和蓝光是植物进行光合作用最主要的光质,不同红、蓝光比例对植物的株高、地径及叶片大小会有不同影响。张亚如等[21]利用不同的红蓝光比例处理桉树组培苗时发现蓝光处理的桉树组培苗的株高更高;史文景[22]用不同红蓝光处理隋珠草莓发现:随着蓝光比例的增加其茎粗增加。本研究中,蓝膜处理下西南杨幼苗的苗高极显著高于白膜,而地径无显著差异。推测光质比例对不同植物的生长影响可能有所不同,特别是草本植物与木本植物之间可能存在较大差异。
3.2 光质对西南杨叶片的影响
叶片大小综合反映了叶片的功能,是植物适应环境的重要性状,在高海拔地区生长的植物叶片小于低海拔地区生长的植物叶片[23]。张悦[24]研究证明:红光、蓝光和绿光都能够促进阔叶苦苣和细叶苦苣叶片的生长,其中高比例的红光更有助于叶长和叶宽增加;杨金钰[25]的研究也证明:红、蓝光为4∶1 是大麦生长的最适光质,能够显著提高其叶面积和叶长。本研究中,西南杨原生地为高海拔地区,其短波光比例高、光照强度更强,因此,西南杨对高短波光比例及高光照强度有较好的适应性;而蓝膜下的光照强度低于白膜,其叶长和叶宽极显著高于白膜,推测西南杨幼苗叶片形态受光照强度的影响比光质的影响更大。环境因子的改变能显著影响植物的光合特性,光质作为光的重要属性,不仅通过影响叶绿素吸收光的方式调控植物对光能的利用,还通过影响植物叶片的组织结构进而引起光合功能的改变[26-27]。利用不同光质处理小白菜的研究发现:蓝光有利于其叶片发育,其栅栏组织和海绵组织的厚度增加[28]。本研究中,蓝膜处理下西南杨幼苗叶片的上表皮厚度显著低于白膜,而叶片、下表皮、栅栏组织和海绵组织的厚度均略高于白膜。推测由于西南杨生长于高海拔地区,环境中短波光比例较高,其幼苗对蓝膜下的光质环境有较好的适应性,因此蓝膜处理不会对其叶片结构造成较大影响。
3.3 光质对西南杨光合效能的影响
影响植物光合作用最主要的2 个因素是CO2浓度和光照强度。气孔是CO2 进入植物体内的通道,已有研究表明:蓝光有利于气孔发育,使其气孔导度更大,进而影响植物的光合作用[29]。本研究表明:蓝膜下西南杨幼苗的Pn、Ci、Gs 值均高于白膜,说明蓝膜处理能够增加叶片的气孔导度,大气中的CO2 能够更多地进入叶片,使得Ci 增加,叶片的Pn 得以提高,促进植株生长。此外,研究植物的光及CO2 响应曲线有助于解释环境因子与植物光合产物积累的关系[30]。LSP 可表征植物利用强光的能力,其值越高表明植株能较好地适应强光照环境;LCP、Rd 和AQE 可表征植物利用弱光的能力,LCP 越低表明植株利用弱光的能力越强;Rd 越低则表明在弱光条件下植物呼吸作用较弱、生理活性较低[31];AQE 反映了弱光环境下植物的光能转化效率,自然环境下植物的AQE 通常为 0.03~0.07 μmol/(m²·s)[32],AQE越高则光能转化效率越高。在本研究中,蓝膜处理下西南杨的LSP、LCP 和Rd 无显著差异,而AQE 显著高于白膜,说明蓝膜处理下西南杨幼苗在弱光环境下的光能转化效率更高,其对弱光的适应性比白膜好,这可能与叶绿素含量变化有关。叶绿素吸收光能用于光合作用,本研究显示:蓝膜处理下西南杨幼苗叶片中的叶绿素a 和叶绿素b 含量均高于白膜,因此蓝膜下西南杨幼苗叶片吸收的光能更多,光合作用更强。LPnmax反映了植物叶片的最大光合潜能[33],蓝膜处理下西南杨幼苗的最大光合潜能高于白膜,说明蓝膜能提升西南杨的光合效能。CCP 和CSP 常用于判断植株对不同浓度CO2 的利用能力,CSP 表征植物利用高浓度CO2 的能力,CCP 表征植物对低浓度CO2 的利用能力[34]。一般认为,具有较高CSP 和较低CCP 的植物对高浓度CO2 环境的适应能力较强,其净光合速率高、生长较快[35]。王静敏等[36]研究表明:蓝膜处理可显著提高滇杨扦插苗对强光的适应能力,绿膜处理可显著提高滇杨扦插苗对弱光的适应能力。本研究中,西南杨幼苗在蓝膜处理下的CSP 和CCP 均低于白膜,表明蓝膜下的苗木对低浓度CO2 的利用能力更强,对高浓度CO2 的利用能力较弱,因此西南杨幼苗在2 种处理下表现出CPnmax 高于LPnmax,表明西南杨幼苗在强光下的Pn 更大程度上受CO2 浓度的限制[35]。α 反映了植株叶片RubisCO的活性和含量,其值越高表明植株对CO2 的利用越充分。在本研究中,蓝膜处理下的α 值极显著高于白膜,表明其叶片RubisCO 的活性更强、含量更高,Pn 也更高。
3.4 光质对西南杨光合产物含量的影响
叶片作为光合作用的主要器官,是植物碳水化合物的重要来源,为植物和其他器官的生长提供养分[37]。植物的能量主要由可溶性糖、淀粉等非结构碳提供。可溶性糖是光合作用的直接产物,是植物体内碳运输及代谢的主要形式,其一般作为植物当前生命活动的能源物质;而淀粉作为植物体内能量储存物质,用于满足植物需求[38]。LEWIS 等[20]研究发现:在蓝光下,黄瓜可溶性糖、蔗糖和淀粉的含量均高于其他光质;张亚如等[21]研究发现:红蓝光组合处理的桉树组培苗,组合光中蓝光比例越低,其茎、叶中可溶性糖含量越少,表明蓝光比例的减少能抑制植株碳代谢及生物量合成。在本研究中,2 种颜色薄膜处理下的西南杨幼苗叶片可溶性糖含量无显著差异,但蓝膜处理的淀粉含量极显著高于白膜,可能是由于蓝膜处理增强了西南杨的光合作用,使富余的产物转化为淀粉存储于植株体内。此外,蓝膜处理下西南杨蔗糖含量显著低于白膜,其原因可能是叶片光呼吸速率加快,使得叶片中糖的分解代谢增强,从而降低叶片蔗糖的积累量。
4 结论
蓝膜处理能够极显著提高西南杨幼苗的苗高,还能够通过增加叶片的气孔导度、叶绿素含量、RubisCO 的含量及活性来增强西南杨的光合作用,最终改善西南杨幼苗的生长状态。研究结果为西南杨引种栽培光质条件的选择与优化提供了依据,也为研究西南杨适应高海拔地区光质的机制提供了基础。
[ 参考文献 ]
[1]LI Y, LIU Z L, SHI Q H, et al. Mixed red and blue lightpromotes tomato seedlings growth by influencing leafanatomy, photosynthesis, CO2 assimilation and endogenoushormones[J]. Scientia Horticulturae, 2021, 290(11):500. DOI: 10.1016/j.scienta.2021.110500.
[2]TARAKANOV I G, TOVSTYKO D A, LOMAKIN MP, et al. Effects of light spectral quality on photosynthet-ic activity, biomass production, and carbon isotope fractionationin lettuce, Lactuca sativa L., plants[J]. Plants,2022, 11(3): 441. DOI: 10.3390/plants11030441.
[3]TARAKANOV I G, KOSOBRYUKHOV A A, TOVSTYKOD A, et al. Effects of light spectral quality on themicropropagated raspberry plants during ex vitro adaptation[J]. Plants, 2021, 10(10): 2071. DOI: 10.3390/plants10102071.
[4]VITALE L, VITALE E, FRANCESCA S, et al. Plantgrowthpromoting microbes change the photosyntheticresponse to light quality in spinach[J]. Plants, 2023,12(5): 1149. DOI: 10.3390/plants12051149.
[5]HAO Y H, ZENG Z X, ZHANG X L, et al. Green meansgo: green light promotes hypocotyl elongation via brassinosteroidsignaling[J]. Plant Cell, 2023, 35(5): 1304.DOI: 10.1093/plcell/koad022.
[6]王春平, 高爽, 薛占军, 等. 有色膜光谱下苗用型大白菜生长及光合活性的基因型响应[J]. 河北农业大学学报,2022, 45(3): 47. DOI: 10.13320/j.cnki.jauh.2022.0042.
[7]付卫民, 王淑芬, 王秀峰, 等. 有色材料覆盖对心里美萝卜品质及产量的影响[J]. 山东农业科学, 2011(6): 39.DOI: 10.14083/j.issn.1001-4942.2011.06.030.
[8]曹明, 杨小锋, 米庆华, 等. 不同颜色棚膜对甜瓜生长及果实品质的影响[J]. 山东农业大学学报(自然科学版),2017, 48(1): 20. DOI: 10.3969/j.issn.1000-2324.2017.01.004.
[9]靳志勇, 刘娜, 艾希珍, 等. 不同棚膜覆盖对秋季青花菜生长及品质的影响[J]. 北方园艺, 2015(3): 56. DOI: 10.11937/bfyy.201503018.
[10]陈丽, 白瑞雯, 汤海昆, 等. 全光照下覆盖不同颜色薄膜对越橘光合特性和果实品质的影响[J]. 植物生理学报,2020, 56(9): 2007. DOI: 10.13592/j.cnki.ppj.2019.0251.
[11]马宏, 孟红志, 李世军, 等. 不同光质对‘春美’桃光合特性和果实品质的影响[J]. 果树学报, 2017, 34(7): 835.DOI: 10.13925/j.cnki.gsxb.20160414.
[12]贾晨, 辜云杰, 何承忠, 等. 川西高原5种乡土杨树生长特性研究[J]. 西北农林科技大学学报(自然科学版),2017, 45(2): 79. DOI: 10.13207/j.cnki.jnwafu.2017.02.012.
[13]纵丹, 王静敏, 张垚, 等. 西南地区9种乡土杨树的秋季光合特性比较[J]. 西北林学院学报, 2022, 37(4): 57.DOI: 10.3969/j.issn.1001-7461.2022.04.08.
[14]周乃富, 张俊佩, 刘昊, 等. 木本植物非均质化组织石蜡切片制作方法[J]. 植物学报, 2018, 53(5): 653. DOI: 10.11983/CBB18051.
[15]段利民, 闫雪, 刘廷玺, 等. 多重情景下半干旱地区人工杨树林光合生理特征[J]. 水土保持学报, 2019, 33(1):213. DOI: 10.13870/j.cnki.stbcxb.2019.01.034.
[16]蔡永萍. 植物生理学实验指导[M]. 北京: 中国农业大学出版社, 2014.
[17]高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006.
[18]THORNLEY J H M. Mathematical models in plantphysiology[M]. London: Academic Press, 1976.
[19]BALY E C C. The kinetics of photosynthesis[J]. Proceedingsof the Roval Society B: Biological Sciences,1935, 117(804): 218. DOI: 10.1098/rspb.1935.0026.
[20]LEWIS J D, OLSZYK D, TINGEY D T. Seasonal patternsof photosynthetic light response in Douglas-fir seedlingssubjected to elevated atmospheric CO2 and temperature[J]. Tree Physiology, 1999, 19(4/5): 243. DOI:10.1093/treephys/19.4-5.243.
[21]张亚如, 陈光彩, 叶春海, 等. LED红蓝光质对桉树组培苗生长及生理特性的影响[J]. 贵州农业科学, 2021, 49(4):97. DOI: 10.3969/j.issn.1001-3601.2021.04.016.
[22]史文景. 不同红蓝光处理对“隋珠”草莓光合特性、产量和果实品质的影响[J]. 中国南方果树, 2021, 50(3):151. DOI: 10.13938/j.issn.1007-1431.20200757.
[23]LIU W S, ZHENG L, QI D H. Variation in leaf traits at differentaltitudes reflects the adaptive strategy of plants toenvironmental changes[J]. Ecology and Evolution, 2020,10(15): 8166. DOI: 10.1002/ece3.6519.
[24]张悦. 不同光质、光照强度及光周期对苦苣生长特性及营养品质的影响[D]. 武汉: 华中农业大学, 2021.
[25]杨金钰. 水培大麦苗生长及饲用营养物质代谢对不同光配方和氮素的响应[D]. 乌鲁木齐: 新疆农业大学,2022.
[26]PANG Y D, LIAO Q H, PENG H G, et al. CO2 mesophyllconductance regulated by light: a review[J]. Planta,2023, 258(1): 11. DOI: 10.1007/s00425-023-04157-5.
[27]王纪, 李飘, 李娜, 等. 青钱柳无性系生长、叶解剖结构和光合色素含量的差异[J]. 西部林业科学, 2023, 52(2):132. DOI: 10.16473/j.cnki.xblykx1972.2023.02.018.
[28]樊小雪, 高文瑞, 孙艳军, 等. 不同光质对小白菜叶片发育和光合作用的影响[J]. 信阳师范学院学报(自然科学版), 2018, 31(4): 562. DOI: 10.3969/j.issn.1003-0972.2018.04.010.
[29]任毛飞, 毛桂玲, 刘善振, 等. 光质对植物生长发育、光合作用和碳氮代谢的影响研究进展[J]. 植物生理学报,2023, 59(7): 1211. DOI: 10.13592/j.cnki.ppj.300151.
[30]周玉霞, 巨天珍, 王引弟, 等. 4种光响应曲线模型对3种高寒草甸植物的实用性分析[J]. 草地学报, 2018, 26(2):488. DOI: 10.11733/j.issn.1007-0435.2018.02.031.
[31]章毅, 蔡建国, 孙欧文, 等. 水淹胁迫下绣球光合响应机制的研究[J]. 核农学报, 2019, 33(4): 808. DOI: 10.11869/j.issn.100-8551.2019.04.0808.
[32]韩晓, 王海波, 王孝娣, 等. 基于4种光响应模型模拟不同砧木对夏黑葡萄耐弱光能力的影响[J]. 应用生态学报, 2017, 28(10): 3323. DOI: 10.13287/j.1001-9332.201710.030.
[33]叶子飘. 光合作用对光和CO2响应模型的研究进展[J].植物生态学报, 2010, 34(6): 727. DOI: 10.3773/j.issn.1005-264x.2010.06.012.
[34]刘杨杨, 李俊, 于强, 等. 甘蔗叶片光合CO2响应参数分析及其品种间差异[J]. 中国农业气象, 2019, 40(10): 637.DOI: 10.3969/j.issn.1000-6362.2019.10.004.
[35]赖帅彬, 潘新雅, 简春霞, 等. 转苜蓿 MsOr 基因烟草光合—光响应和光合—CO2响应曲线特征研究[J]. 草地学报, 2020, 28(1): 20. DOI: 10.11733/j.issn.1007-0435.2020.01.003.
[36]王静敏, 杨青青, 胡治旭, 等. 光质对滇杨叶片结构及光能利用特性的影响[J]. 云南农业大学学报(自然科学),2022, 37(5): 862. DOI: 10.12101/j.issn.1004-390X(n).202203036.
[37]YANG H, LI Y P, QIAO Y Z, et al. Low light stress promotesnew tiller regeneration by changing source-sink relationshipand activating expression of expansin genes inwheat[J]. Plant, Cell amp; Environment, 2023, 46(5): 1562.DOI: 10.1111/pce.14548.
[38]曹宝慧. 不同外源光对三个树种幼苗生长以及碳水化合物和养分积累的影响[D]. 沈阳: 沈阳农业大学,2020.
责任编辑:何承刚
基金项目:云南省应用基础研究项目(202101AU070144);云南省农业基础研究联合专项(202101BD070001-127);云南省“兴滇英才支持计划”青年人才专项(XDYC-QNRC-2022-0232);生物学质量工程项目(503190106)。
猜你喜欢
学与玩(2022年6期)2022-10-28 09:18:52
科学(2022年4期)2022-10-25 02:43:00
——记嘉荫县红光乡燎原村党支部
奋斗(2020年24期)2021-01-16 06:23:36
家庭影院技术(2019年8期)2019-08-27 02:45:04
家庭影院技术(2019年1期)2019-01-21 02:25:26
家庭影院技术(2018年10期)2018-11-02 05:35:38
Coco薇(2016年5期)2016-06-03 09:17:41
考试周刊(2016年6期)2016-03-11 08:14:32
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:45:31
无机化学学报(2014年1期)2014-02-28 17:30:11
