
北方根结线虫在党参上的群体动态及世代循环
2024-08-04 00:00:00谢志军石明明吴锦张玲陈京环李惠霞
云南农业大学学报(自然科学) 2024年3期
关键词:党参
摘要: 【目的】明确甘肃省党参上北方根结线虫(Meloidogyne hapla) 的群体动态和世代循环,以期为根结线虫病的综合防治提供科学依据。【方法】2021 年,通过田间定点连续取样对土壤中的北方根结线虫2 龄幼虫和根组织中各虫态进行分离和镜检计数。【结果】在甘肃省,从4 月上旬开始,北方根结线虫在党参上1 年发生4 代,平均每代历期为36 d,完成1 代最短需要28 d,最长为53 d。田间气温和降水量对北方根结线虫发育影响较大,当田间气温为18 ℃、降水量为141~176 mm 时,北方根结线虫发育较快,完成1 代平均为30 d。【结论】本研究明确了北方根结线虫在甘肃省党参上的群体动态及世代循环,对该线虫病的及时防控具有指导意义。
关键词: 党参;北方根结线虫;群体动态;世代循环
中图分类号: S435.672 文献标志码: A 文章编号: 1004–390X (2024) 03−0040−06
根结线虫作为高等植物的重要病原生物之一,广泛分布于世界各地,主要危害蔬菜、粮食作物、果树、观赏植物、中药材等,每年造成的全球经济损失超过1 570 亿美元[1-2]。目前,已报道的根结线虫有100 多种,其中南方根结线虫(Meloidogyne incognita)、花生根结线虫(M. arenaria)、爪哇根结线虫(M. javanica) 和北方根结线虫(M. hapla) 发生最为普遍[3]。北方根结线虫为冷凉地区农作物的重要病原物,寄主范围包括党参、白芷、黄芩、甘草、当归等多种中药材,造成药材须根增多,主根生长不良,严重影响药材产量和品质[4]。
党参[Codonopsis pilosula (Franch.) Nannf.] 为桔梗科(Campanulaceae) 党参属(Codonopsis) 植物,是中国传统药食同源中药材[5-6]。甘肃省为中国党参主产区,有“千年药乡”之称。据统计,党参种植面积约为4.22×104 hm2,总产量达1.025×105 t,居于全国前列[7]。本课题组首次报道了北方根结线虫可引起党参根结线虫病[8],系统调查了该病害在甘肃省的发生分布,发现6 个党参主产县均有根结线虫病发生,部分产区发病严重,且有向全省蔓延的趋势[9]。根结线虫主要以2 龄幼虫或卵在土壤和病残体内存活,一般分布在5~30 cm 土层中。外界条件适宜时,土壤中的卵孵化为2 龄幼虫,侵入植物根系,导致根部膨大,产生根结。2 龄幼虫建立取食位点摄取营养物质,经3 次蜕皮发育为成虫,雌雄虫交配后,雄虫死亡,雌虫产卵。卵囊中的卵孵化为2 龄幼虫,散落到土壤中再侵染或越冬。根结线虫的生长发育受寄主、温度、湿度等诸多因素的影响,一般根结线虫完成1 个生活史需要30~60 d。目前,关于北方根结线虫侵染特性研究较少,尚缺乏该线虫在田间的群体动态和世代发生研究。
鉴于此,明确北方根结线虫在党参上的田间侵染循环,对该线虫病害防治时期的选择乃至综合防控具有重要实践意义。本研究于2021 年4—10 月,通过田间定点连续取样,对党参根组织进行染色,观测北方根结线虫在党参上的群体动态变化,结合当地气象数据,分析气温、降水量等对北方根结线虫世代发生的影响,以期明确党参根结线虫病的田间侵染循环。
1 材料与方法
1.1 试验设置
试验地位于甘肃省定西市临洮县窑店镇武家村(35°19′42″N, 104°2′24″E),属于温带大陆性气候,年均气温10.3 ℃,无霜期80~190 d,年降水量317~760 mm,降雨多集中在7—9 月。试验地为砂壤土,连续多年种植党参。经鉴定,病原线虫种类为北方根结线虫,且在田间分布较均匀。调查取样期间进行正常田间管理[10]。
1.2 样品采集
2021 年4 月7 日—10 月17 日,每隔7 d 取样1 次。采用多点随机取样法,在每个样点采集党参植株和根际土样,混合为1 个样本,装入自封袋中,立即运回实验室[11]。
1.3 土壤中线虫的分离与计数
参考刘维志[12]的方法,将每次采集的试样混匀后,随机称取100 g,用改良贝尔曼漏斗法收集土壤中的2 龄幼虫。在漏斗内放置1 个直径4 cm 的硬塑料网作支撑,其上铺1 层面巾纸,将土壤轻放于纸上,漏斗下端用橡胶管连接1 个指形管,在漏斗内加适量清水,静置24 h 后收集指形管内线虫悬浮液,弃上清液,在体式显微镜(OLYMPUS SZX16 体视显微镜,日本奥林巴斯公司) 下观察分离到的2 龄幼虫并计数。每次取样至少重复3 次。
1.4 党参根组织中的线虫染色及计数
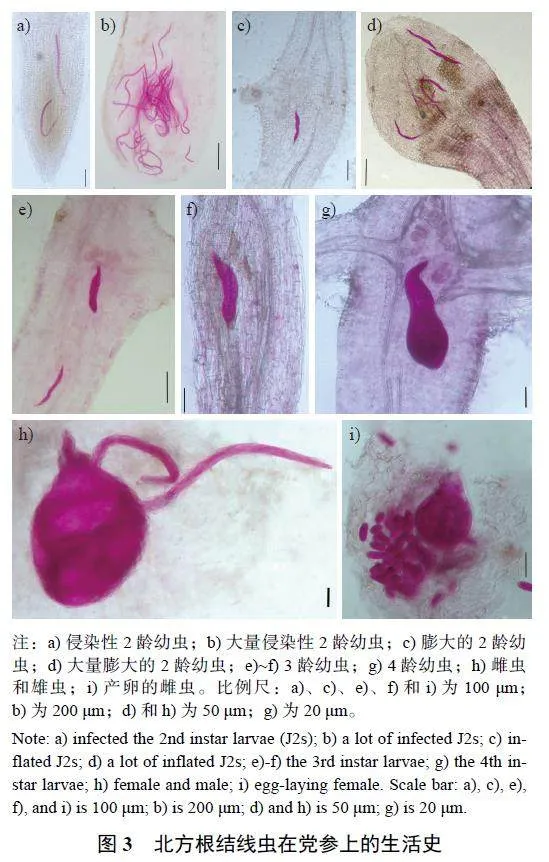
剪取党参根系样品的全部须根,置于80 目筛网中清洗干净,用吸水纸吸取表面明水后称量;采用次氯酸钠—酸性品红染色法[13]对党参根组织进行染色,在体式显微镜下观察并统计根组织中各虫态及数量,最后置于生物显微镜(Axio Lab.A1 显微镜,德国Carl Zeiss 公司) 下拍照。每个样品重复3 次。
1.5 根结线虫世代划分标准
党参根结线虫完成世代数和每代历期的界定,主要依据土壤中2 龄幼虫的数量及根组织内雌虫产卵量的峰值,其次考虑根内2 龄幼虫侵入量的峰值以及3 龄幼虫、4 龄幼虫和雌虫形成的峰值等信息[14]。根据临洮县当年气象数据,分析世代发生与平均气温和平均降水量的关系。采用Excel 2019 和SPSS 19.0 进行数据分析。
2 结果与分析
2.1 土壤中2 龄幼虫的动态变化
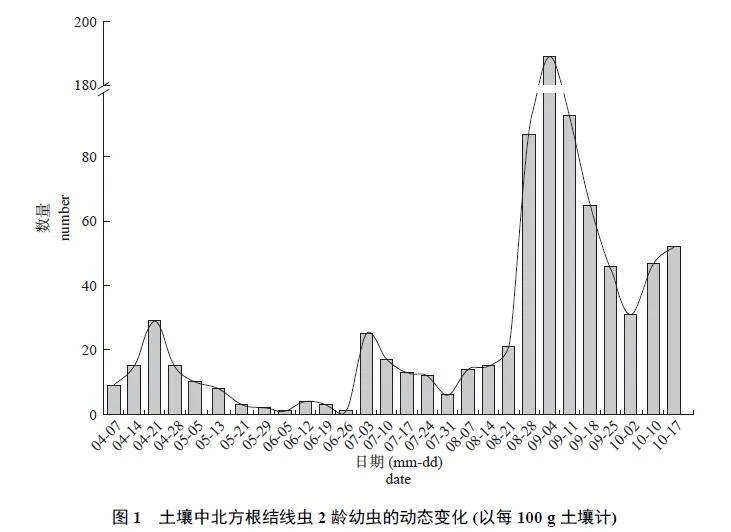
从4 月7 日—10 月17 日,党参根际土壤中均检测到根结线虫2 龄幼虫,共出现3 个比较明显的峰值,分别为4 月21 日、7 月3 日和9 月4 日(图1),表明北方根结线虫在党参上1 年最少发生3 代。其中,9 月4 日土壤中2 龄幼虫密度为全年最高,每100 g 土壤中有189 条;6 月5 日和6 月26 日2 龄幼虫极少,每100 g 土壤中仅有1 条,表明在党参生长期间北方根结线虫2 龄幼虫反复侵染根部,临近收获期侵染最为严重。
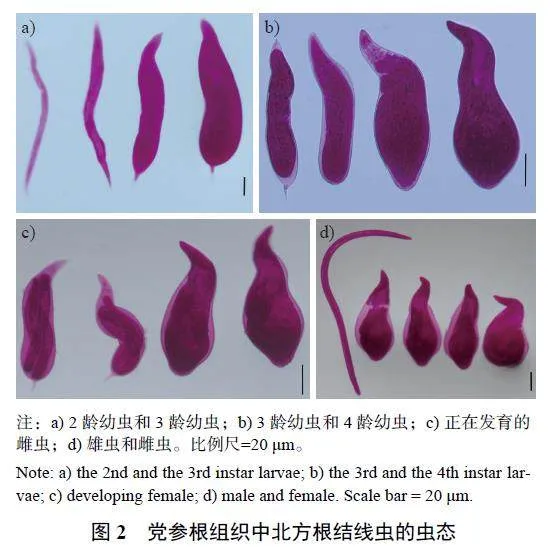
2.2 党参根组织中各虫态的群体动态
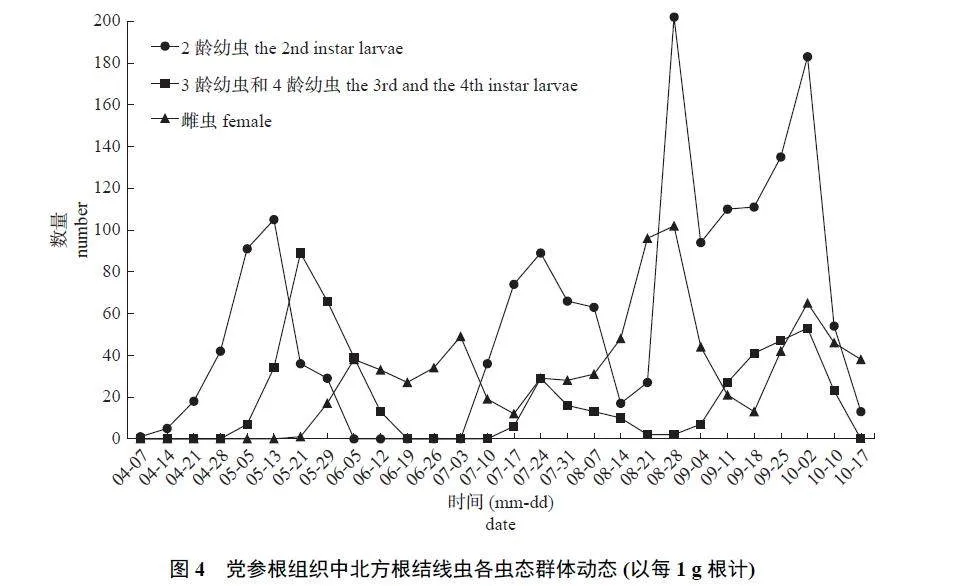
观察党参根组织内北方根结线各虫态形态(图2) 及其生活史(图3),可知党参根组织中各虫态的群体动态(图4)。从苗期至收获期,在党参根组织中均检测到2 龄幼虫,其密度有4 个明显的峰值, 分别为5 月13 日、7 月24 日、8 月28 日和10 月2 日,其中8 月28 日的密度在全年中最高,每1 g 根中有202 条。结合土壤中2 龄幼虫的动态变化可知:土壤中2 龄幼虫形成高峰会导致根组织中2 龄幼虫的高峰,表明土壤中2 龄幼虫大部分能侵染党参根组织,若不对其加以控制,对党参根部具有巨大的危害。此外,5 月13 日在党参根组织首次检测到根结线虫3 龄幼虫和4 龄幼虫,且5 月13 日—6 月12 日、7 月17 日—10 月10 日均检测到3 龄幼虫或4 龄幼虫,其密度有3 个明显的峰值,分别为5 月21日、7 月24 日和10 月2 日,其中,5 月21 日密度全年最高,每1 g 根中有89 条。5 月21 日—10 月17 日均能检测到根组织内的根结线虫雌虫,其密度有4 个明显的峰值,分别为6 月5 日、7 月3 日、8 月28 日和10 月2 日,并在7 月24 日出现1 个不明显的峰值,其中8 月28 日根组织内雌虫最多,每1 g 根中有102 条。
2.3 北方根结线虫的世代循环
党参根组织中2 龄幼虫侵入有4 个明显的高峰,每个高峰发生时或发生后会伴随着3 龄幼虫、4 龄幼虫或雌虫形成的高峰。可见,北方根结线虫在党参上1 年发生4 代,各代历期长度不一。
第1 代从4 月14 日开始,持续到6 月5 日,历期约为53 d。土壤中卵孵化和2 龄幼虫侵入速度较慢,根组织内各龄期幼虫数量多,但发育缓慢,世代历期最长。第2 代为7 月3 日—7 月31 日,历期28 d。土壤中2 龄幼虫侵入和各个虫态发育速度较快,从7 月3 日开始下降,7 月31 日达到最小值,随后又开始上升,世代历期最短。第3 代为7 月31 日—9 月4 日,历期35 d。该代发生特点是历期较长,根组织内各虫态发育较迟缓,土壤中2 龄幼虫持续增加,9 月4 日达到最大值,随后下降;根组织中3 龄幼虫、4 龄幼虫和雌虫也持续增加,到8 月28 日达到最大值,随后下降。第4 代从9 月4 日开始发生,到10 月2 日结束,历期28 d。在此期间,土壤中2 龄幼虫平均密度全年最高,根组织中2 龄幼虫、3 龄幼虫和4 龄幼虫的数量不断增加,雌虫数量先减少后增加,9 月18 日有1 个明显的低峰。
2.4 气象因素对北方根结线虫发生的影响
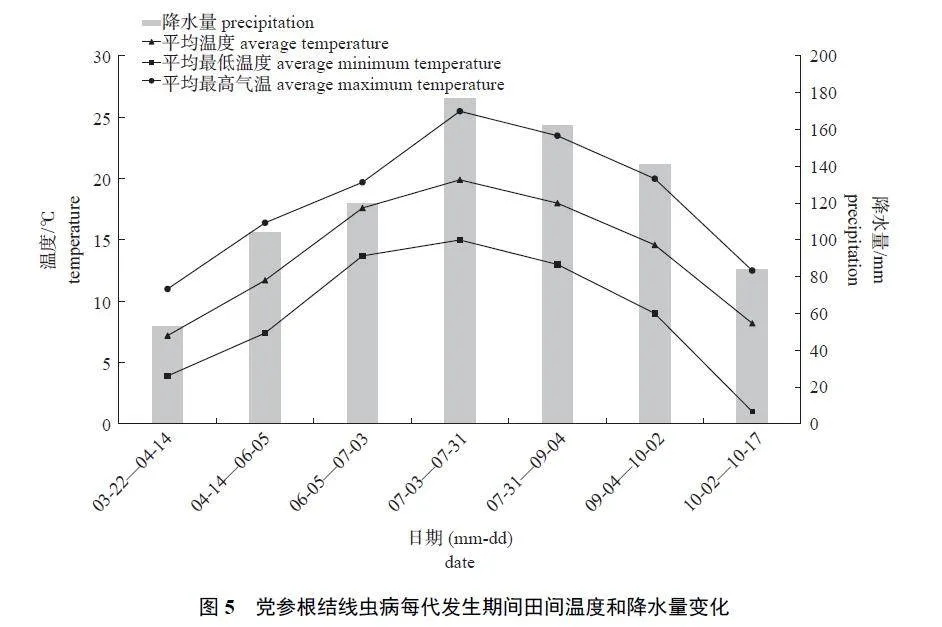
该试验田在党参生长期间无人工灌溉,故田间气温和降水量是影响党参根结线虫发生的主要因素。北方根结线虫侵染党参的第1 代发生期间,田间平均气温11.7 ℃,平均最低气温7.4 ℃,平均最高气温16.4 ℃,田间降水量122.4 mm。在4 代中第1 代发生期间的平均气温和降水量均最低,历期最长。随着田间气温和降水量增加,北方根结线虫完成1 代所需时间缩短(图5)。第2 代至第4 代期间,田间平均气温和降水量均高于第1 代,历期较短,为28~35 d。上述结果表明:田间气温和降水量是影响北方根结线虫完成1 代的主要因素。当田间平均气温为18 ℃、降水量为141~176 mm 时,北方根结线虫在党参上完成1 代的历期较短。
3 讨论
本研究表明:北方根结线虫在甘肃省党参上1 年发生4 代,平均历期为36 d,当外界条件适宜时,完成1 个生活周期仅需28 d。而在山东烟台地区,北方根结线虫在花生上1 年发生3 代,完成1 代时间约为48 d, 完成1 代最短需要39 d[15],与本研究结果差异较大,可能是寄主或地理差异所致。不同种的根结线虫侵染寄主后,完成1 代所需时间也不一致。南方根结线虫引起的黄瓜根结线虫病1 年发生5~6 代,完成1 个生活史需25 d[16];而香蕉根结线虫病在广西南宁1 年可发生9 代,平均30~45 d 完成1 代[17];在孟加拉国部分地区,拟禾本科根结线虫(M. graminicola)在水稻上只需19 d 即可完成1 代[18];禾谷孢囊线虫(Heterodera avenae) 在北京小麦田1 年发生2 代[19],在江苏省1 年仅发生1 代[20]。可见,不同病原线虫在不同寄主上的世代发生差异较大,地理差异可能是其主要影响因素。
根结线虫从卵孵化到雌虫产卵完成1 个周期还受田间温度、降水量、土壤质地、pH 等因素的影响。本研究表明:当田间平均气温为18 ℃、降水量为141~176 mm 时,北方根结线虫发育最快,完成1 代约为30 d,低于第1 代历期的53 d。该结果与其他根结线虫报道类似。崔江宽等[21]发现:烟草根结线虫(M. incognita 和 M. hapla) 在20~28 ℃ 条件下20 d 可完成1 代,但当温度低于10 ℃ 时,根结线虫停止发育,不能完成生活史。为害河南园林苗木的北方根结线虫在23~27 ℃时,完成1 代约为60 d;但温度低于12 ℃ 时,2龄幼虫活动能力减弱,不能侵入根系[22]。SINGH等[23]也发现:拟禾本科根结线虫在湿度32%、温度22~29 ℃ 的环境下,2 龄幼虫的侵入和发育较快。但温、湿度如何影响北方根结线虫侵染和发育,还需进一步调查和研究。
近年来,党参根结线虫病已成为甘肃省党参生产的一类重要病害,但目前相关研究较少,且该病害未受到管理部门和种植户重视,这可能会导致党参根结线虫快速蔓延,对党参产业造成巨大的影响。党参为多年生植物,种植方式主要为移栽幼苗,而前期调查发现部分育苗地已发生根结线虫病。因此,种苗调运时应检测幼苗是否携带病原线虫,并在移栽前应进行土壤处理,以降低线虫数量。本研究表明:在第1 代发生期间,土壤中卵的孵化所需时间较长,且幼虫侵染和发育速度较慢,因此,在生长期应以第1 代为主进行防治。该病害田间变化复杂,且受环境条件影响,对其侵染特性及田间的精准防控还有待进一步研究。
4 结论
不同地区、不同寄主对北方根结线虫发育影响较大,当外界条件适宜时,北方根结线虫在甘肃省党参上能较快完成1 代。在生产中,依据田间侵染规律选取最佳防治时期是保证甘肃省党参产业高质量发展的关键。
[ 参考文献 ]
[1]ABAD P, GOUZY J, AURY J M, et al. Genome sequenceof the metazoan plant-parasitic nematode Meloidogyneincognita[J]. Nature Biotechnology, 2008, 26(8):909. DOI: 10.1038/nbt.1482.
[2]PERRY R N, MOENS M, STARR J L. Root-knot nematodes[M]. USA: CABI, 2013.
[3]金娜, 陈永攀, 刘倩, 等. 我国蔬菜根结线虫发生、致害和绿色防控研究进展[J]. 植物保护学报, 2022, 49(1):424. DOI: 10.13802/j.cnki.zwbhxb.2022.2022828.
[4]PERRY R N, MOENS M. 植物线虫学[M]. 简恒, 译. 北京: 中国农业大学出版社, 2011.
[5]谢琦, 程雪梅, 胡芳弟, 等. 党参化学成分、药理作用及质量控制研究进展[J]. 上海中医药杂志, 2020, 54(8):94. DOI: 10.16305/j.1007-1334.2020.08.013.
[6]金俊奇, 陈代文, 毛湘冰, 等. 党参多糖的生物学功能及其在畜牧生产中的应用[J]. 动物营养学报, 2021, 33(5):2527. DOI: 10.3969/j.issn.1006-267x.2021.05.013.
[7]李成义, 刘书斌, 李硕, 等. 甘肃党参栽培现状调查分析[J]. 中国现代中药, 2016, 18(1): 102. DOI: 10.13313/j.issn.1673-4890.2016.1.014.
[8]LI W H, LI H X, LIU Y G, et al. First report of northroot-knot nematode Meloidogyne hapla on Codonopsispilosula in China[J]. Plant Disease, 2020, 104(8): 2295.DOI: 10.1094/pdis-02-20-0295-pdn.
[9]石明明, 李惠霞, 刘永刚, 等. 甘肃省党参根结线虫病发生、分布及病原种类鉴定[J]. 植物保护, 2022, 48(3):181. DOI: 10.16688/j.zwbh.2021203.
[10]王振华, 时立波, 吴海燕, 等. 大豆根内胞囊线虫发育进程及分布[J]. 中国农业科学, 2009, 42(9): 3147. DOI: 10.3864/j.issn.0578-1752.2009.09.016.
[11]赵洪海, 程子超, 王凤龙, 等. 山东省烟草孢囊线虫的群体动态和世代发生特点[J]. 植物保护学报, 2013, 40(6):529. DOI: 10.13802/j.cnki.zwbhxb.2013.06.008.
[12] 刘维志. 植物线虫志[M]. 北京: 中国农业出版社, 2004.
[13] 段玉玺. 植物线虫学[M]. 1版. 北京: 科学出版社, 2011.
[14]刘国坤, 肖顺, 张绍升, 等. 拟禾本科根结线虫对水稻根系的侵染特性及其生活史[J]. 热带作物学报, 2011,32(4): 743. DOI: 10.3969/j.issn.1000-2561.2011.04.034.
[15]宋协松. 花生根结线虫(Meloidogyne hapla Chitwood)的生活史及温湿度影响[J]. 植物病理学报, 1992,22(4): 59.
[16]陈志杰, 张淑莲, 张锋. 设施蔬菜根结线虫防治基础与技术[M]. 北京: 科学出版社, 2013.
[17]李树庆. 香蕉根结线虫生物学特性及防治研究[D]. 南宁: 广西大学, 2002.
[18]RAO Y S, ISRAEL P. Life history and bionomics of Meloidogynegraminicola, the rice root-knot nematode[J].Indian Phytopathology, 1973, 26(2): 333.
[19]LIU S S, ZHAO J L, AHMED S, et al. Population dynamicsand infection of cereal cyst nematode (Heteroderaavenae) in wheat in Beijing[J]. 云南农业大学学报(自然科学), 2017, 32(1): 1. DOI: 10.16211/j.issn.1004-390X(n).2017.01.001.
[20]向桂林, 宋志强, 梁旭东, 等. 禾谷孢囊线虫的田间侵染规律及垂直分布研究[J]. 麦类作物学报, 2013, 33(4):789. DOI: 10.7606/jissn.1009-1041.2013.04.29.
[21]崔江宽, 任豪豪, 孟颢光, 等. 我国烟草根结线虫病发生与防治研究进展[J]. 植物病理学报, 2021, 51(5): 663.DOI: 10.13926/j.cnki.apps.000487.
[22]周思源. 根结线虫病的发生与防治[J]. 现代农村科技,2021(11): 32.
[23]SINGH I, GAUR H S, BRIAR S S. Role of wheat in sustainingMeloidogyne graminicola in rice-wheat croppingsystem[J]. International Journal of Nematology, 2003,1(13): 79.
责任编辑:何承刚
基金项目:国家自然科学基金项目(31760507);国家重点研发计划子课题(2018YFC1706301-5)。
猜你喜欢
中国民族民间医药(2023年1期)2023-02-16 05:53:42
Natural Products and Bioprospecting(2021年2期)2021-04-10 01:42:56
中成药(2018年5期)2018-06-06 03:12:14
中成药(2018年3期)2018-05-07 13:34:21
中成药(2018年4期)2018-04-26 07:12:58
中成药(2018年1期)2018-02-02 07:20:31
中成药(2017年10期)2017-11-16 00:50:06
中成药(2017年6期)2017-06-13 07:30:35
中国中医药信息杂志(2016年5期)2016-12-01 06:07:16
农家科技中旬版(2016年9期)2016-11-02 04:33:50
