
4种夏栎对低温胁迫的生理响应及抗寒性评价
2024-07-18 00:00:00姜恒张志刚杨建军李斌王兴胜张晓红
河北大学学报(自然科学版) 2024年4期


DOI:10.3969/j.issn.10001565.2024.04.009
摘要:为探究低温胁迫下夏栎(Quercus robur)生理指标的响应及抗寒性,以普通夏栎、速生夏栎、裂叶夏栎和垂枝夏栎为实验材料,测定在不同温度下细胞膜通透性、超氧化物歧化酶(SOD)、过氧化物酶(POD)、可溶性蛋白等指标,并利用拟合Logistic方程计算半致死温度,结合隶属函数法对4种夏栎综合评价.结果表明:4种夏栎细胞膜通透性均随着温度降低而增加,SOD和POD活性以及可溶性蛋白、脯氨酸和丙二醛(MDA)含量均逐渐下降;4种夏栎半致死温度为-41.9~-38.2 ℃,其中普通夏栎半致死温度最低,速生夏栎半致死温度最高;4种夏栎抗寒性强弱为普通夏栎gt;垂枝夏栎gt;裂叶夏栎gt;速生夏栎.综上可知,普通夏栎抗寒性最强,在不同地区推广和繁育具有显著优势.
关键词:夏栎;抗寒性;隶属函数法;生理指标;半致死温度
中图分类号:Q945.79文献标志码:A文章编号:10001565(2024)04040608
Physiological response and cold resistance evaluation of four
species of Quercus robur to low temperature stress
JIANG Heng1,2, ZHANG Zhigang3 , YANG Jianjun1,2, LI Bin4, WANG Xingsheng4, ZHANG Xiaohong4
(1. Key Laboratory of Oasis Ecology of Education Ministry, College of Ecology and Environment,
Xinjiang University,Urumqi" 830017,China; 2.Technology Innovation Center for Ecological
Monitoring and Restoration of Desert-Oasis," Ministry of Natural Resources (MNR), Urumqi "830002,China;
3. Key Laboratory of Forest Resources and Utilization in Xinjiang of National Forestry and Grassland
Administration, Institute of Afforestation and Desertification Control, Xinjiang Academy of Forestry,
Urumqi" 830000,China; 4. Yili Prefecture Forest Tree Breeding Experiment Center, Chabchal" 835311, China)
Abstract: In order to explore the response of Quercus robur" physiological indicators and the strength of cold resistance under low temperature stress, the common Q. robur, fast-growing Q. robur, split-leaf Q. robur and weeping Q. robur were used as experimental materials. The cell membrane permeability, SOD activity, POD activity and soluble protein at different temperatures were measured, and the fitted Logistic
收稿日期:20240109;修回日期:20240508
基金项目:新疆三农骨干人才培养项目(2022SNGGNT082);新疆维吾尔自治区2023年度重大科技专项(2023A02008)
第一作者:姜恒(1998—),男,新疆大学在读硕士研究生,主要从事生态修复研究.E-mail:670551763@qq.com
通信作者:杨建军(1978—),男,新疆大学教授,博士生导师,主要从事干旱区水土保持与荒漠化防治研究.
E-mail:yjj@xiu.edu.cn
张晓红(1972—),女,伊犁州林木良种繁育试验中心高级工程师,主要从事林木良种选育与推广研究.
E-mail:1436787731@qq.com
equation was used to calculate the half-lethal temperature. And the function method was used for comprehensive evaluation of the four Q. robur species. The results showed that the cell membrane permeability of the four species of Q. robur showed an increasing trend as the temperature decreased, and the SOD activity, POD activity, soluble protein content, proline content and MDA content showed a decreasing trend as a whole, and the cell membrane permeability was calculated by combining the Logistic equation. The semi-lethal temperature of four species of Q. robur ranged from -41.9 ℃ to -38.2 ℃. The common Q. robur had the highest semi-lethal temperature and the fast-growing Q. robur had the lowest semi-lethal temperature. This is consistent with the evaluation by the membership function method. The four species of Q. robur had strong cold resistance. The cold resistance ranking of the four Q. robur species was common Q. roburgt; weeping Q. roburgt;split-leaf Q. robur gt; fast growing Q. robur. The study show that common Q. robur is the most resistant to cold and has significant advantages for promotion and breeding in different regions.
Key words: Quercus robur; cold resistance; membership function method; physiological index; half-lethal temperature
夏栎(Quercus robur)隶属壳斗科(Fagaceae)栎属(Quercus),落叶乔木,也称夏橡、英国栎,其树体高大通直,成年后树高可达40 m,在中国的新疆、北京和山东均有栽培,具有材质好、寿命长、树冠开阔和树形优美等优点,对生长环境要求较低,生长范围十分广泛,是十分理想的造林树种,在干旱和半干旱区林业造林中具有良好的推广前景[1-2].冬季低温是制约植物能否安全越冬的重要因素[3],受全球气候变化影响,春季气候多变,寒潮、冰雹、低温霜冻等灾害性天气频发,严重阻碍了植物的生长发育[4],因此,研究夏栎在低温胁迫下的生理响应及抗寒性对夏栎推广栽培具有重要意义.
低温胁迫下,植物体内会发生一系列生理生化反应来适应外界低温,单一的指标无法对植物的抗寒性进行评价.李呈呈等[5]对6个彩叶桂品种的研究发现,细胞膜通透性、超氧化物歧化酶(SOD)、过氧化物酶(POD)和可溶性蛋白等指标对植物抵御低温胁迫有重要作用,可作为植物抗寒性评价指标;牟开萍等[6]以20个月季品种为研究对象,测定了相对电导率(REC)、丙二醛(MDA)和可溶性蛋白等,并结合主成分分析、聚类分析和隶属函数法对其进行了综合评价;贾金辉等[7]以8种酿酒葡萄为研究对象,测定了低温胁迫下枝条萌芽率和相关生理指标,发现隶属函数法和半致死温度评价结果的一致性;杨振亚等[8]研究了娜塔栎、沼生栎、舒玛栎、双色栎在低温胁迫下的响应,并通过半致死温度确定了其抗寒性.因此,选择合适的指标和评价方法尤为重要.目前关于夏栎的研究多集中在育苗移栽和白粉病等方面[9-11],而对夏栎抗寒性的研究较少.
本研究选取垂枝夏栎、速生夏栎、裂叶夏栎和普通夏栎1年生枝条为实验材料,在人工模拟低温的条件下,通过测定细胞膜通透性、可溶性蛋白和MDA含量及SOD和POD活性,并拟合Logistic方程计算半致死温度,采用隶属函数法对4种夏栎进行综合评价,筛选出抗寒性强的夏栎品系,为夏栎在不同地区推广和繁育提供理论依据.
1材料与方法
1.1实验材料
实验材料来自伊犁州林木良种繁育试验中心,品系为垂枝夏栎、速生夏栎、裂叶夏栎和普通夏栎.先使用自来水清洗枝条表面灰尘,再用去离子水冲洗3次,用纱布吸干表面水分.枝条清洗完毕后,对其进行剪切(避开芽孢,每段至少留存2个芽孢),每段长10 cm,将剪切好的枝条装入自封袋,并贴好标签,标记好品系名称.
1.2实验方法
实验开始后将全部枝条同时放入低温冰柜内.实验处理设置为6个温度梯度,分别为5、-20、-25、-30、-35、-40 ℃,以5 ℃为对照,温度调整为每8 h下降0.6 ℃,达到对应温度后保持24 h,随后取出样条测定抗寒生理指标并进行水培实验.
将低温胁迫的枝条用去离子水冲洗3次,将枝条下端1 cm剪除,置于三角瓶中水培,每个处理10段枝条,将三角瓶放置于组培架上,阳光培养12 h,每隔2 d换水、观测枝条复苏情况并统计枝条成活率(芽鳞开裂、膨大为成活).根据枝条恢复后存活枝的成活率评价各品系的抗寒性.
枝条成活率=成活枝数/水插枝总数×100%.
1.3指标测定及方法
生理指标测定:SOD活性采用氮蓝四唑还原法测定[12];POD活性采用愈创木酚法测定[12];可溶性蛋白含量采用考马斯亮蓝法测定[12];MDA采用硫代巴比妥酸法测定[12];脯氨酸采用茚三酮显色法测定[13].
细胞膜通透性测定:细胞膜通透性采用电导率法测定[12].对不同温度下各品系夏栎细胞膜通透性进行Logistic拟合,计算出各品系的半致死温度(LT50)[14].
拟合方程为Y=K/(1+ae-bx),(1)
LT50=-(ln a)/b,(2)式中:Y为相对电导率;K为常数,代表电解质渗出的最大量;e为自然底数;a、b均为方程参数;x为处理温度;LT50为曲线的拐点温度(半致死温度).
1.4数据处理及分析
采用隶属函数法对不同品系夏栎进行评价,与抗寒性呈正相关指标(SOD、POD、可溶性蛋白、脯氨酸),计算公式为U(Xij)=(Xij-Xjmin)/(Xjmax-Xjmin),(3)与抗寒性呈负相关指标(相对电导率和丙二醛含量),计算公式为U(Xij)=1-(Xij-Xjmin)/(Xjmax-Xjmin),(4)式中:U(Xij)为各指标的平均值;Xij为第i个品系下第j个指标的隶属度值;Xjmin和Xjmax分别为第j个指标不同温度下最小值和最大值.
对各品系在不同温度下的隶属度值求均值,按照隶属度值排序即为不同品系夏栎的抗寒能力.使用Microsoft Office和Origin 2022进行数据整理,SPSS 27.0进行单因素方差分析和显著性分析(Plt;0.05).
2结果与分析
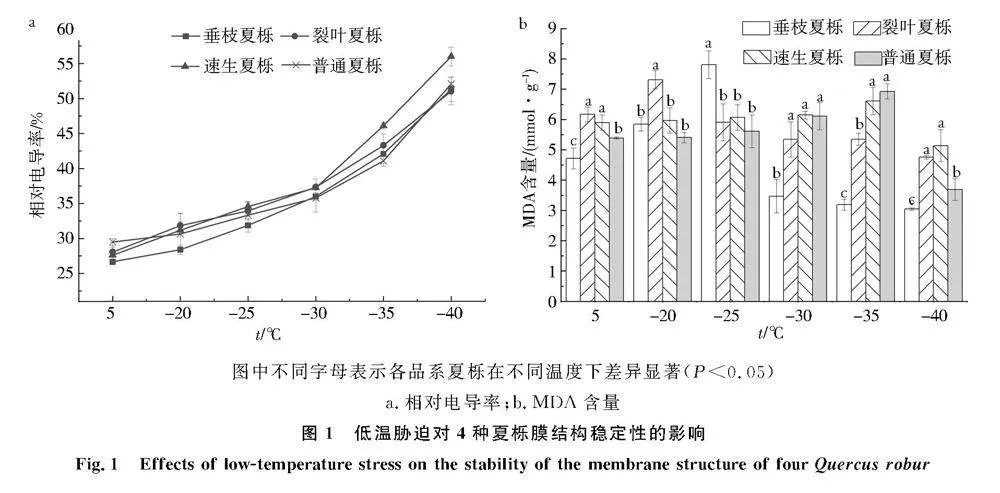
2.1低温胁迫对夏栎细胞膜通透性的影响
低温胁迫下夏栎各品系细胞膜通透性变化如图1a所示,随着温度的降低,各品系细胞膜通透性均呈上升趋势,其中垂枝夏栎和普通夏栎的变化趋势较为一致.对照组4种夏栎枝条相对电导率均保持在28%~33%,并在-40 ℃时均达到最大值.对照组垂枝夏栎细胞膜通透性最小,为28.39%,其次是裂叶夏栎,为30.65%,当温度为-30~-25 ℃时,速生夏栎的细胞膜通透性变化最大,而普通夏栎的变化最小,其余2种均在二者之间,4种夏栎叶片细胞膜通透性达到最大值时,相对电导率依次为速生夏栎gt;裂叶夏栎gt;普通夏栎gt;垂枝夏栎.
2.2低温胁迫对夏栎MDA的影响
低温胁迫下夏栎各品系MDA含量变化如图1b所示,随着温度的降低,各品系的MDA含量均呈现先上升后下降的趋势,其中速生夏栎和普通夏栎MDA含量的变化趋势较为一致.经过-35 ℃处理后,速生夏栎和普通夏栎枝条的MDA含量均达到最大值,分别为6.6 mmol/g和6.9 mmol/g,而裂叶夏栎和垂枝夏栎分别在-20 ℃和-25 ℃处理下达到最大值,分别为7.3 mmol/g和7.8 mmol/g.垂枝夏栎的MDA含量在低温胁迫下均与其他品系存在显著差异.与对照组相比,4种夏栎枝条受到低温胁迫后MDA含量整体表现为速生夏栎gt;裂叶夏栎gt;垂枝夏栎gt;普通夏栎.
图中不同字母表示各品系夏栎在不同温度下差异显著(Plt;0.05)
a.相对电导率;b.MDA含量
2.3Lotistic方程的建立及LT50的确定
低温胁迫下对4种夏栎相对电导率进行方程拟合,结果如表1所示,低温胁迫处理下各品系夏栎的相对电导率和温度之间的关系可以用公式(1)和公式(2)拟合,R2值为0.891~0.979,表明温度和相对电导率有很强的线性关系,拟合结果具有可靠性.4种夏栎的LT50为-41.9~-38.2 ℃,根据LT50大小可得夏栎抗寒性为普通夏栎gt;裂叶夏栎gt;垂枝夏栎gt;速生夏栎.表1低温胁迫下4种夏栎的LT50及拟合方程
Tab.1LT50 and fitting equations of four Quercus robur under low-temperature stress品系拟合方程R2t/℃垂枝夏栎Y=100/(1+6.997e-0.048)0.915-40.5速生夏栎Y=100/(1+5.957e-0.047)0.891-38.2裂叶夏栎Y=100/(1+5.781e-0.043)0.908-41.1普通夏栎Y=100/(1+5.269e-0.04)0.979-41.9
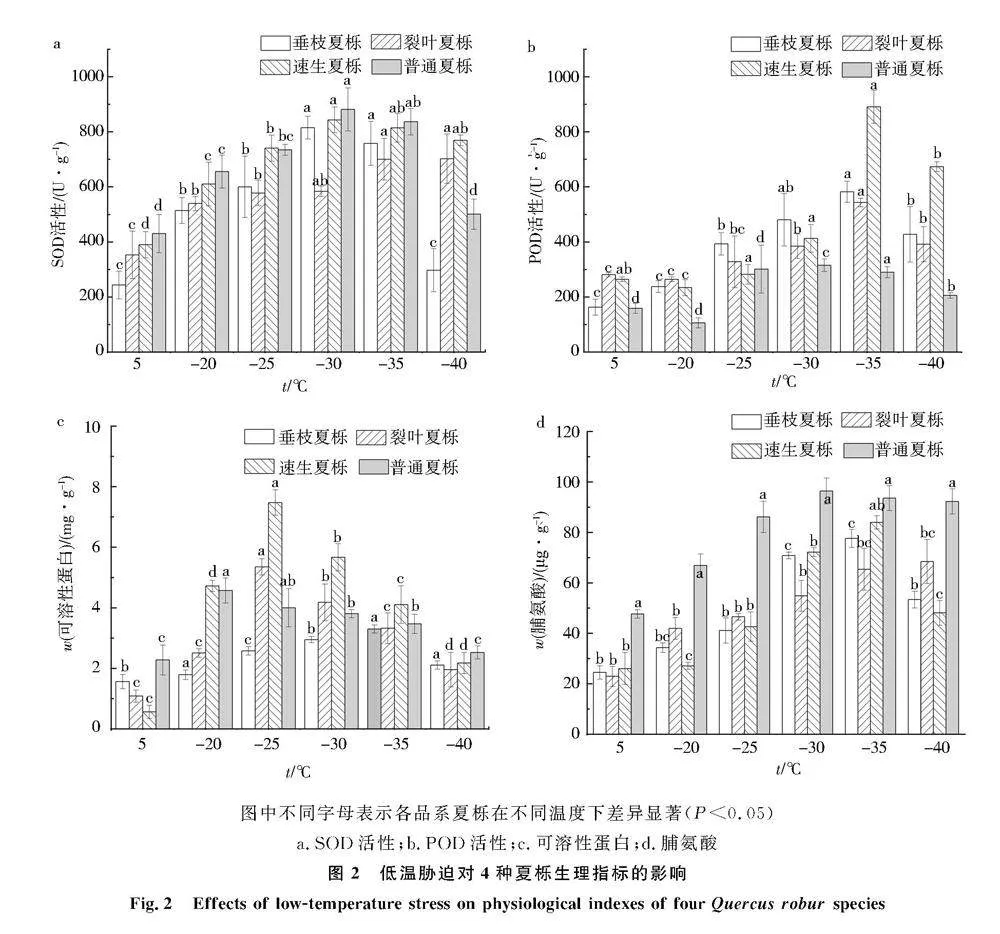
2.4低温胁迫对夏栎SOD的影响
低温胁迫下夏栎SOD酶活性变化如图2a所示,随着温度的降低,4种夏栎1年生枝条SOD酶活性呈现先升高后降低的趋势.4种夏栎SOD酶活性最大值均不相同,其中垂枝夏栎、速生夏栎和普通夏栎均在-30 ℃时枝条的SOD活性达到最大值,分别为815.01、843.28、881.19 U/g,而裂叶夏栎的SOD活性在-35 ℃达到最大值,为702.15 U/g.与对照组相比,垂枝夏栎的SOD含量增加最多,为235%,裂叶夏栎的SOD含量增加最少,为98.55%.4种夏栎枝条受到低温胁迫后SOD活性增加幅度为垂枝夏栎gt;速生夏栎gt;普通夏栎gt;裂叶夏栎.
2.5低温胁迫对夏栎POD的影响
低温胁迫后4种夏栎的POD活性如图2b所示,其中,普通夏栎对照组的POD活性为159 U/g,当经过-20 ℃处理后,POD活性有所下降,但与对照组差异不显著,随着温度的降低,POD活性逐渐增加,到-30 ℃时达到最大值,为315.33 U/g,比对照组增加了98.3%,之后随着温度的降低,POD活性随之降低,但均显著高于对照组.垂枝夏栎、裂叶夏栎和速生夏栎的POD活性整体呈现出先增加后下降的趋势,变化较为相似,在-35 ℃低温胁迫下,垂枝夏栎、裂叶夏栎和速生夏栎的POD活性均达到最大值,分别为582、543、890.67 U/g,且速生夏栎POD活性显著高于其他3种品系,比对照组分别增加了257.1%、93.2%和236.95%,之后随着温度的增加POD活性随之下降.4种夏栎POD活性低温胁迫后整体上升幅度为速生夏栎gt;垂枝夏栎gt;裂叶夏栎gt;普通夏栎.
2.6低温胁迫对夏栎可溶性蛋白含量的影响
低温胁迫下4种夏栎的可溶性蛋白含量(质量分数)如图2c所示,对照组4种夏栎可溶性蛋白含量均处于最小值,其中速生夏栎含量最低,其次为普通夏栎,而裂叶夏栎的可溶性蛋白含量最高,且显著高于其他3种品系.随着温度的降低,各品系夏栎呈现出先增加后下降的趋势,4种夏栎达到最大值的可溶性蛋白含量各不相同,-35 ℃处理后除速生夏栎的可溶性蛋白含量与对照组无显著差异外,其他温度处理下均存在显著差异.普通夏栎的可溶性蛋白含量在-20 ℃处理下达到最大值,相较于对照组增加了321.3%,裂叶夏栎在-25 ℃处理下达到最大值,增加了135.2%,4种夏栎受到低温胁迫后可溶性蛋白含量增加幅度为速生夏栎gt;裂叶夏栎gt;普通夏栎gt;垂枝夏栎.
2.7低温胁迫对夏栎脯氨酸含量的影响
低温胁迫下4种夏栎的脯氨酸含量(质量分数)如图2d所示,随着温度的降低,速生夏栎、垂枝夏栎和普通夏栎的脯氨酸含量呈现先增加后下降的趋势,而裂叶夏栎的脯氨酸含量呈现出持续增大的趋势.在-35 ℃处理下,垂枝夏栎和速生夏栎的脯氨酸含量达到最大值,普通夏栎的脯氨酸含量在-30 ℃处理下达到最大值,相较于对照组增加了102.3%.与对照组相比,4种夏栎受到低温胁迫后脯氨酸含量达到最大值时增加幅度整体为垂枝夏栎gt;速生夏栎gt;裂叶夏栎gt;普通夏栎.图中不同字母表示各品系夏栎在不同温度下差异显著(Plt;0.05)
a.SOD活性;b.POD活性;c.可溶性蛋白;d.脯氨酸
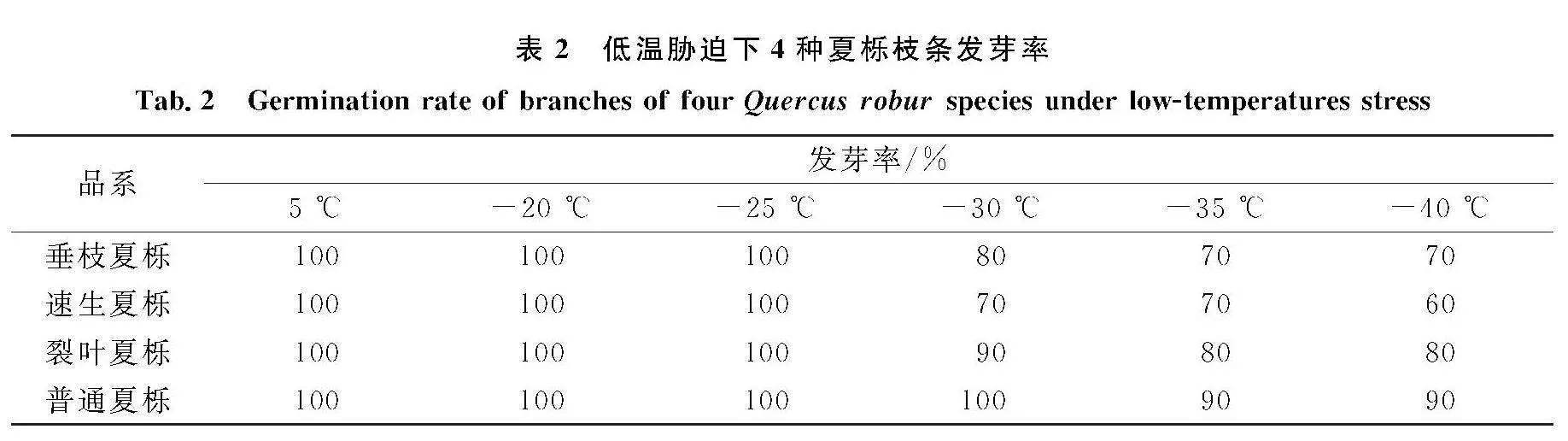
2.8低温胁迫下不同品系夏栎枝条水培萌发情况
水培恢复法是验证植物在抵御低温胁迫后是否遭受冻害最简单有效的方法.低温胁迫后不同品系水培萌芽情况如表2所示,随着温度的降低,不同夏栎发芽率也有所不同,在-25~-20 ℃处理后,4种品系夏栎水培发芽率均达到100%;-30 ℃处理后,垂枝夏栎、裂叶夏栎和速生夏栎的发芽率均有所降低,分别为80%、70%和90%.随着温度的下降,普通夏栎、裂叶夏栎和垂枝夏栎的发芽率较为稳定,经过-40 ℃处理后,速生夏栎枝条的发芽率最低,为60%;普通夏栎的发芽率最高,为90%,表明4种品系夏栎均能在-40 ℃低温下存活,并且普通夏栎抗寒性强于垂枝夏栎、裂叶夏栎和速生夏栎.
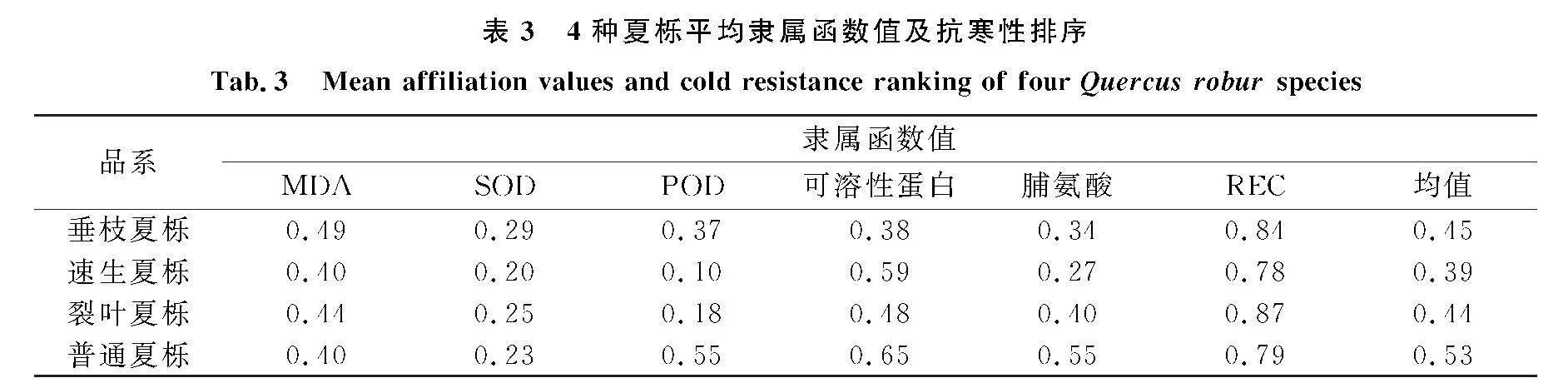
2.9不同品系夏栎的抗寒性分析
对低温胁迫下4种品系夏栎的细胞膜通透性等生理指标采用隶属函数进行综合分析及评价.隶属函数值与抗寒性存在正相关关系,隶属函数值越大,其抗寒性越强,反之抗寒性越弱.
由表3可知,4种夏栎中普通夏栎抗寒性最强,速生夏栎的抗寒性最差,4种夏栎抗寒性由大到小依次为普通夏栎gt;垂枝夏栎gt;裂叶夏栎gt;速生夏栎,这一结果与水培复苏实验的结果基本一致.
3讨论与结论
细胞膜是植物受到低温胁迫后细胞反应最直接的部位,也是容易受到低温胁迫伤害的部位[15-17].低温胁迫下植物细胞膜结构和功能发生改变,细胞内离子和糖类物质渗出,通透性增大,从而导致胞外电导率增大.因此,电导率值可用来表示植物在低温胁迫下受影响的程度,且相对电导率越大,表明植物越容易受到低温胁迫的影响,抗寒性越弱[18].本研究中,4种夏栎相对电导率随着温度的降低呈增加趋势,这与王金鑫等[19]在研究低温胁迫对不同果型翅果油树细胞膜通透性变化表现一致.温度高于-35 ℃时,4种夏栎细胞膜通透性表现相当;温度低于-35 ℃时,速生夏栎的细胞膜通透性迅速增大,表明速生夏栎的抗寒性低于其他3种夏栎.LT50表示植物抗寒能力和耐低温的极限,LT50越低,表明植物的抗寒性越强[20].本研究中,4种夏栎抗寒性从小到大排序为速生夏栎<垂枝夏栎<裂叶夏栎<普通夏栎,LT50依次为-38.2、-40.5、-41.1、-41.9 ℃.
MDA是细胞膜过氧化的最终产物,已经成为植物抗寒评价的重要指标之一.MDA在植物体内的积累量与植物的抗寒性有较强的相关性,MDA含量越高,说明细胞膜过氧化越严重,即抗寒性越弱[21-22].4种夏栎的MDA含量在低温胁迫初期随着温度的降低呈上升趋势,随着低温胁迫的下降,4种夏栎在低温胁迫下MDA含量有所降低,当温度低于-25 ℃时,垂枝夏栎MDA含量显著下降,并显著低于其他品系,表明垂枝夏栎在低温初期抗寒性要弱于其他3种夏栎,这与温日宇等[23]在研究不同低温胁迫下不同藜麦幼苗的MDA含量变化与抗寒性评价结果相一致.随着低温胁迫的加剧,普通夏栎和垂枝夏栎的MDA含量显著低于速生夏栎和裂叶夏栎,即普通夏栎和垂枝夏栎的抗寒性高于裂叶夏栎和速生夏栎,说明低温在胁迫初期导致4种夏栎细胞膜损害加重,但随着低温的加剧细胞膜起到了一定的保护作用,这与LT50计算出的4种夏栎的抗寒性结果相一致.本实验中,4种夏栎在低温胁迫后枝条复苏率与LT50结果基本一致.
低温胁迫下细胞膜脂氧化加重,体内超氧自由基阴离子、过氧化氢、羟基自由基等活性氧含量增加,降低细胞膜的功能,严重时导致植物细胞功能紊乱[24-25].SOD和POD是植物体抗氧化系统中重要的酶,能够清除体内产生过量的活性氧,对维持植物正常的生理功能和抵御低温胁迫具有重要意义,因此在低温胁迫下,SOD和POD活性含量与植物的抗寒性有较强的关系,这与李桂荣等[26]在研究6种山葡萄在低温胁迫下抗氧化酶的结果一致.本研究中4种夏栎的SOD和POD含量在低温胁迫初期有所下降,之后呈上升趋势,这可能是由于低温胁迫初期植物体内保护酶活性降低或分解有关,再次升高可能是由于植物已经适应了低温胁迫从而导致抗氧化酶升高.在温度胁迫下,速生夏栎POD含量在最大值时增加显著高于其他夏栎,相反普通夏栎的变化相对较小,这说明速生夏栎对低温胁迫的变化较为敏感,而普通夏栎对低温胁迫有较强的适应能力.随着温度降低,抗氧化酶活性降低,推测是因为植物细胞膜氧化加剧,活性氧产生超过保护酶的清除能力.
植物细胞渗透调节物质在应对环境胁迫和维持细胞内外水分平衡方面发挥着重要作用[27],低温胁迫条件下,植物细胞会积累可溶性蛋白和脯氨酸,以增加细胞内的渗透浓度,从而缓解低温胁迫对植物造成的损害[28-29].本研究发现抗寒性强的普通夏栎脯氨酸含量高于其他夏栎,即脯氨酸含量与夏栎抗寒性呈正相关,雷梦瑶等[30]对石榴枝条对低温胁迫的响应的结果显示,抗寒性强的千层花和红双喜脯氨酸含量高于另外2种夏栎,与本实验的研究结果一致.从可溶性蛋白含量变化发现,抗寒性差的速生夏栎可溶性蛋白在低温胁迫初期增加最多并显著高于其他3种夏栎,可能是因为速生夏栎对低温变化较为敏感,细胞持水能力下降,需要增加大量的可溶性蛋白来降低细胞冰点,保护植物免受低温损伤.反而抗寒性强的普通夏栎可溶性蛋白含量相较于抗寒性弱的速生夏栎更加稳定,这与李桂荣等[31]的研究结果一致.推测的原因可能是因为普通夏栎体内本身含有较高的可溶性蛋白且本身耐寒性较好,受到低温胁迫后体内可溶性蛋白或仅需小幅度提高即可应对外界胁迫带来的损伤.
评价植物抗寒性的指标有很多,在低温胁迫过程中单一指标无法客观判断植物的抗寒能力[32-33],除了结合多指标对植物的抗寒性进行评价,还需要通过水培条件下不同夏栎的萌芽率来确定植物的抗寒能力.本文采用隶属函数法并结合水培条件下枝条萌芽率对4种夏栎进行综合评价,结果发现隶属函数法与枝条萌芽率结果基本一致.普通夏栎在低温胁迫下表现最好,基本确定普通夏栎可作为在不同地区推广和繁育的品系,4种夏栎抗寒性由强到弱依次为普通夏栎gt;垂枝夏栎gt;裂叶夏栎gt;速生夏栎.
参考文献:
[1]白永强,李永华,朱强,等.夏橡高效育苗技术试验[J].林业科技通讯, 2022(10): 70. DOI:10.13456/j.cnki.lykt.2022.01.21.0001.
[2]石海燕,张志刚,李宏,等.IAA对幼苗期夏橡生长及光合特性的影响[J].河北大学学报(自然科学版), 2022, 42(5): 519-529. DOI:10.3969/j.issn.1000-1565.2022.05.011.
[3]李文明,辛建攀,魏驰宇,等.植物抗寒性研究进展[J].江苏农业科学, 2017, 45(12): 6-11. DOI:10.15889/j.issn.1002-1302.2017.12.002.
[4]HU L, XIAOFENG W, SHUJIANG C, et al. Dynamic analysis and evaluation of Xinjiang forest resources: based on RS and GIS[J]. Journal of Geographical Sciences, 2005, 15(3): 346-352. DOI:10.1007/BF02837522.
[5]李呈呈,吴其超,马燕,等.6个彩叶桂品种对低温胁迫的生理响应及抗寒性评价[J].浙江农林大学学报, 2021, 38(4): 828-836. DOI:10.11833/j.issn.2095-0756.20200606.
[6]牟开萍,李维芳,杨文新,等.20个月季品种的抗寒性综合评价[J].草原与草坪, 2021, 41(6): 58-66. DOI:10.13817/j.cnki.cyycp.2021.06.009.
[7]贾金辉,徐凌,刘慧纯,等.8个酿酒葡萄品种的抗寒性比较[J].中国果树,2021(7):39-44.DOI:10.16626/j.cnki.issn1000-8047.2021.07.008.
[8]杨振亚,台秀国,曹振玉,等.4个引种栎类树种苗期抗寒性初步研究[J].山东大学学报(理学版), 2016, 51(5): 43-47,60. DOI:10.6040/j.issn.1671-9352.0.2015.573.
[9]李素琼,史开奇,张晓红.夏橡白粉病防治[J].农业与技术, 2024, 44(3): 55-57. DOI:10.19754/j.nyyjs.20240215013.
[10]郭英,李开花.珍贵树种夏橡育苗栽培技术[J].农村科技,2016(4): 69-70. DOI:10.3969/j.issn.1002-6193.2016.04.041.
[11]单兴宇.夏橡的育苗和栽培技术[J].现代农机, 2021(3): 109-110. DOI:10.3969/j.issn.1674-5604.2021.03.060.
[12]杨梅花,黄祥,韩彦奇,等.8种睡莲对低温胁迫的生理响应及抗寒性评价[J].江苏农业科学, 2022, 50(23): 123-128. DOI:10.15889/j.issn.1002-1302.2022.23.018.
[13]井俊丽,刘铭潇,魏欣,等.几种苹果中间砧的抗寒性评价[J].果树学报, 2022, 39(6): 970-981. DOI:10.13925/j.cnki.gsxb.20210444.
[14]刘晓东,任伟嘉,何淼.苦水玫瑰和冷香玫瑰抗寒性[J].东北林业大学学报, 2012, 40(11): 28-30,34. DOI:10.3969/j.issn.1000-5382.2012.11.007.
[15]井俊丽,刘铭潇,高美娜,等.不同苹果中间砧枝条在越冬期间的生理特性变化及抗寒性比较[J].河北农业大学学报, 2022, 45(4): 25-31. DOI:10.13320/j.cnki.jauh.2022.0056.
[16]王睿哲,张蓓,郭铁群,等.5个梨品种在低温胁迫下的生理响应及抗寒性评价[J].山东农业科学, 2023, 55(2): 57-63. DOI:10.14083/j.issn.1001-4942.2023.02.008.
[17]王伟强,田新会,杜文华.甘农4号小黑麦苗期抗寒性评价[J].甘肃农业大学学报, 2023, 58(1): 46-54. DOI:10.13432/j.cnki.jgsau.2023.01.006.
[18]赵雪辉,陈双建,成继东,等.3个桃品种抗寒性分析研究[J].果树资源学报, 2020,1(6): 14-19. DOI:10.16010/j.cnki.14-1127/s.2020.06.004.
[19]王金鑫,王延松,明文,等.不同果型翅果油树的抗寒性[J]. 东北林业大学学报, 2023, 51(6): 25-32. DOI:10.13759/j.cnki.dlxb.2023.06.014.
[20]王召元,田启航,常瑞丰,等.桃不同品种对低温胁迫的生理响应及评价[J].中国农业大学学报, 2022, 27(2): 66-77. DOI:10.11841/j.issn.1007-4333.2022.02.07.
[21]刘敏,靳娟,阿布都卡尤木·阿依麦提,等.新疆3个鲜食枣品种的抗寒性评价[J].新疆农业科学, 2023, 60(4): 916-924. DOI:10.6048/j.issn.1001-4330.2023.04.016.
[22]刘在国,赵通,马超,等.7个平欧杂交榛子品种的生理解剖特性及其抗寒性研究[J].干旱地区农业研究, 2023, 41(5): 138-149. DOI:10.7606/j.issn.1000-7601.2023.05.15.
[23]温日宇,刘建霞,李顺,等.低温胁迫对不同藜麦幼苗生理生化特性的影响[J].种子, 2019, 38(5): 53-56. DOI:10.16590/j.cnki.1001-4705.2019.05.053.
[24]曹燕燕,葛昌斌,齐双丽,等.不同冬小麦品种(系)拔节期低温胁迫生理反应及抗寒性评价[J].江苏农业科学, 2022, 50(17): 59-66. DOI:10.15889/j.issn.1002-1302.2022.17.010.
[25]徐功勋,周佳,吕德国,等.4个苹果品种的抗寒性评价[J].果树学报, 2023, 40(4): 669-679. DOI:10.13925/j.cnki.gsxb.20220359.
[26]李桂荣,连艳会,程珊珊,等.低温胁迫对山葡萄等6个葡萄品种抗寒性的影响[J].江苏农业科学, 2019, 47(8): 130-134. DOI:10.15889/j.issn.1002-1302.2019.08.029.
[27]戚建莉,吴文俊,姜成英,等.不同引进希腊品种油橄榄的抗寒性评价[J].经济林研究, 2023, 41(2): 49-57. DOI:10.14067/j.cnki.1003-8981.2023.02.006.
[28]石林曌,倪天虹,裴云霞,等.湖北省主栽油茶品种苗期抗寒性评价[J].经济林研究, 2019, 37(4): 10-19. DOI:10.14067/j.cnki.1003-8981.2019.04.002.
[29]王海鸥,高展,艾克拜尔·毛拉,等.自然越冬条件下17个葡萄砧木品种 几个生理指标变化分析[J].经济林研究, 2019, 37(4): 120-128. DOI:10.14067/j.cnki.1003-8981.2019.04.016.
[30]雷梦瑶,高小峰,白清敏,等.不同品种石榴枝条的抗寒性评价[J].河南农业科学, 2023, 52(6): 120-130. DOI:10.15933/j.cnki.1004-3268.2023.06.013.
[31]李桂荣,连艳会,程珊珊,等.低温胁迫下不同无核葡萄品种抗寒性的分析[J].西南农业学报, 2018, 31(11): 2399-2406. DOI:10.16213/j.cnki.scjas.2018.11.029.
[32]孟诗原,吕桂云,张明忠,等.5种卫矛属植物对低温胁迫的生理响应及抗寒性评价[J].西北植物学报, 2020, 40(4): 624-634.
[33]阳妮,蒋景龙,李丽,等.3个柑橘品种幼苗对低温胁迫的生理响应及其抗寒性评价[J].中国果树, 2020(5): 32-39, 143. DOI:10.16626/j.cnki.issn1000-8047.2020.05.006.
(责任编辑:赵藏赏)
猜你喜欢
热带作物学报(2022年5期)2022-06-01 00:18:44
吉林农业·下半月(2017年1期)2017-02-16 11:31:56
江苏农业科学(2016年8期)2017-02-15 20:29:25
江苏农业科学(2016年8期)2017-02-15 20:14:21
安徽农学通报(2017年1期)2017-02-15 18:23:48
物联网技术(2016年12期)2017-01-21 22:27:52
新疆农垦科技(2016年2期)2016-08-21 13:50:26
中国科技博览(2016年1期)2016-04-25 07:47:28
中国果菜(2016年9期)2016-03-01 01:28:40
中国马铃薯(2015年5期)2016-01-09 06:11:11
