
产红青霉菌对脲醛树脂的降解机制
2024-07-15 00:00:00郭仕伟向阳仝迎芳刘亚青郭建峰白玉洁高焕秦旭巍王芳
山西农业大学学报(自然科学版) 2024年3期





摘要:[目的]每年有30 多万吨脲醛用做树脂、粘合剂和绝缘材料,由于其降解持久性,此类物质废弃后对环境造成广泛的损害。本研究以产红青霉菌(Penicillium rubens 23229)为对象,研究其对脲醛树脂(UF)的降解过程和降解酶的性质。[方法]用X⁃射线衍射(XRD)和失重率评价P. rubens 23229 及其酶的降解能力,通过液相-质谱联用(LCMS)分析降解过程中间代谢产物,应用凝胶色谱柱对降解酶进行纯化。[结果]P. rubens 23229 作用于UF,30 d 后,材料失重率(降解率)为35%,结晶度由26. 21% 提升至29. 44%,经计算得出,降解的35% 包括UF 结晶区7%,非结晶区28%;从菌种和脲醛共培养液中得到P. rubens 23229 脲醛降解粗酶液,其最适作用温度为50 ℃,最适pH 为4. 0,LC-MS 结果表明该粗酶液在降解过程中,首先将高分子UF 主链降解,形成四甲基五脲、三甲基四脲等短链中间代谢物,然后进一步分解中间代谢产物为二氧化碳以及甲醛等终产物,降解作用位点包括脲醛主链中亚甲基与尿素间的C-N、酰胺键等。进一步对比了P. rubens 23229 粗酶液与商业脲酶、蛋白酶对UF 的降解能力,发现蛋白酶几乎无降解效果,脲酶有一定降解能力,但显著低于P. rubens 23229 粗酶液。粗酶经纯化后降解效率提高28 倍,凝胶渗透色谱(GPC)显示出分子量为6. 89 kD 和220. 83 kD 的2 个峰,对应前面降解过程分析,说明降解过程包含主链降解酶和中间代谢物降解酶至少2 种酶的复合体系共同发挥作用。[结论]P. rubens 23229 脲醛降解酶由主链降解酶和中间代谢物降解酶共同组成,其中包含脲酶作用类似的酶,但降解主链的酶与已有蛋白酶作用方式不同,其具体的组成与性质有待于进一步研究。本研究对UF 的生物降解具有一定的参考价值。
关键词:脲醛树脂; P. rubens 23229; 脲醛降解酶; 生物降解
中图分类号:Q89;Q939.96 文献标识码:A 文章编号:1671-8151(2024)03-0099-09
脲醛树脂(UF)是由尿素和甲醛聚合而成的高分子材料,其主链连接键为酰胺键以及C-N[1-3],其中酰胺键结构类似于蛋白质中的肽键。甲醛与尿素的比例不同,聚合反应后会形成三维交联和线性2 种结构的过渡结构,甲醛比例高更倾向于交联结构,而尿素比例高更倾向于线性结构,导致UF 在硬度、绝缘性、结晶度等性能方面存在一定的差异[4-5],从而使得UF 具有广泛的用途。每年有30 多万吨脲醛用做树脂、粘合剂和绝缘材料[6]。由于其降解持久性,这些树脂产品废弃后对环境造成广泛的损害。2016 年的《国家危险废物名录》已明确将废UF 等树脂类废物列为第13 类危险废弃物[7]。
随着UF 用量的增加,其降解性能逐渐成为研究热点[8]。焚烧是世界范围内经常使用的有机废物处理技术,但运营成本高,且会产生大量空气污染物[9]。热解可以将UF 转化为高价值的燃料或化学产品,但在废气中通常存在有毒含氮气体,如一氧化氮、异氰酸等,且热解要求材料干燥,处理成本较高[10-12]。水解UF 的过程中需要盐酸或过氧化氢等化学试剂的参与,且降解后会留下含有甲醛的废液[7]。因此迫切需要开发环境友好的回收利用和降解脲醛的方式,利用微生物降解UF 作为一种清洁技术,具有环境安全、成本效益和不产生二级废物的优点,因而越来越受到重视[13-15]。
作为人工合成的有机高分子聚合物,UF 的降解过程可以参考聚对苯二甲酸乙二醇酯等塑料制品。有机高分子聚合物的降解过程,通常分为几个阶段,首先,材料通过机械、光、热、酸碱等理化作用降解成微粒及较低分子量的聚合物,然后进一步由理化作用或微生物参与彻底代谢为二氧化碳、水及生物质[16],如果微生物无法彻底代谢或代谢速度远远低于产生速度,塑料微粒则经由海洋、土壤等环境在生物链中逐步累积,造成巨大危害[17]。筛选能降解塑料微粒或有机大分子化合物的微生物成为解决问题的关键,其生物降解过程及作用机制也是研究热点。
通常随着UF 分子量增大,交联度和结晶度增高,UF 抗生物降解性增加。目前对UF 生物降解的研究较少。Jahns 等[18]采用从Rhodococcus、Burkholderia 等细菌中纯化到能降解亚甲基脲(MDU)、异丁烯二脲(IDU)等脲醛小分子中间代谢物的降解酶,提出了亚甲基脲的降解途径,该过程包含1 个MDUase(EC3. 5. 1. 21),其分子量为189 kD,可以将亚甲基脲水解逐步脱氨基、脱羧,代谢为尿素和甲醛。本实验室[19]设计了1 种以含磷、钾的脲醛肥料为唯一碳源和氮源的筛选培养基,首次筛选到降解脲醛缓释肥料的真菌产红青霉菌(Penicillium rubens 23229)。
热解是脲醛主链降解为小分子聚合物的1 个途径,为了研究脲醛主链的生物降解,实验对脲醛材料进行热处理后得到相对耐热的脲醛材料,发现P. rubens 23229 可以针对耐热脲醛进行降解,说明P. rubens 23229 可以降解脲醛缓释肥的主链。但是其降解过程和降解酶性质尚不明确[19]。此外,P. rubens 23229 筛选中用到的脲醛肥料含磷酸根基团,易成为主链降解的突破点。废弃UF通常不含磷酸根基团,且结晶度和硬度相对较高,其主链尤其不易被生物降解。P. rubens 23229 对不含磷酸根基团的UF 是否有降解能力,能否在废弃UF 的生物降解中发挥作用,仍不得而知。针对以上问题,本文通过分析降解效率及中间代谢产物,探究P. rubens 23229 对结晶度相对较高的UF的降解过程及降解酶性质。
1 材料与方法
1. 1 试剂与仪器
试剂:脲醛树脂(UF)以尿素和甲醛(1. 5∶1)为原料酸催化制备,山西省高分子复合材料工程技术研究中心制备;葡聚糖凝胶G-50,索莱宝科技有限公司;脲酶(1. 7 eU·mg-1),上海迈坤化工有限公司;胃蛋白酶(1200 U·g-1),国药集团化学试剂有限公司。
仪器:立式压力蒸汽灭菌器,BXM-30R(上海博迅实业有限公司医疗设备厂,上海);净化工作台,SW-CJ-2FD(上海博迅实业有限公司医疗设备厂,上海);X-射线衍射仪,MiniFlex600(日本理学公司,日本);高效液相色谱-质谱联用,LCMS-8050(岛津实验器材有限公司,日本);凝胶渗透色谱,Agilent 1260(安捷伦科技有限公司,美国)。
1. 2 耐热UF 的制备
参照王芳等[19]方法处理UF,将处理后的UF于40 ℃干燥箱中烘干,备用。
1. 3 P. rubens 23229 的培养及降解酶液的制备
用脲醛培养基[19]培养P. rubens 23229,其中脲醛缓释肥改用UF,按2% 的含量接种P. rubens23229,在28 ℃ 、100 rpm 的水浴摇床中培养。取P. rubens 23229 与UF 共培养液,用1 张中速滤纸过滤,弃去滤渣,在4000 rpm 条件下离心20 min,离心上清液即为P. rubens 23229 降解粗酶液。
1. 4 P. rubens 23229 对UF 的降解分析
脲醛培养基与P. rubens 23229 共培养,每隔3 d 取1 次混匀的培养液样品,加入200 mL 去离子水,在温度为100 ℃的条件下,恒温水浴锅中搅拌30 min,随后用1 张快速滤纸过滤,弃去滤液。加入200 mL 去离子水重悬,用0. 45 μm 的滤膜抽滤,弃去滤液。将剩余的脲醛树脂于40 ℃干燥箱中烘干24 h,计算材料失重率,公式如下:
M =(m1 - m2)/m1
式中,M:失重率;m1:降解前材料质量,g;m2:降解后材料质量,g。
1. 5 X⁃射线衍射分析
利用X - 射线衍射仪(XRD)分别对UF、经100 ℃热处理后的UF 和进一步经P. rubens 23229共培养30 d 后的UF 的晶型进行表征。扫描角度范围为10~80 °,扫描速度为1 °·min-1。材料结晶度由MDI jade 6 和HighScore Plus 分析得出。
1. 6 降解酶性质研究
1. 6. 1 最适温度及温度稳定性
量取10 mL 降解酶液,加入1 g UF,分别在10、20、30、40、50、60、75 ℃条件下,培养36 h,测定甲醛含量及其失重率。
1. 6. 2 最适pH 及pH 稳定性
分别取10 mL 降解酶液,分别调节其pH 为2、3、4、5、6、7、8、9,加入1 g UF,常温条件下培养36 h,测定甲醛含量及其失重率。
1. 7 不同酶降解性能比较
分别取10 mL 去离子水、10 mL P. rubens23229 降解酶液、10 mL 去离子水加50 mg 蛋白酶、10 mL 降解酶液加50 mg 脲酶、10 mL 去离子水加50 mg 脲酶,加入1 g UF,在常温条件下降解24 h,测定甲醛含量及其失重率。
1. 8 降解中间代谢产物的HPLC-MS 分析
P. rubens 23229 降解粗酶液处理UF,15 d 后取上清液,过0. 22 μm 水系过滤膜,取过滤液进行高效液相色谱- 质谱联用(HPLC-MS)分析。HPLC 条件:采用色谱柱(Waters C18 1. 7 μm,2. 1×100 mm)分离溶液,流动相甲醛-水溶液(体积比为9∶1),柱温为40 ℃ ,流速0. 75 mL·min-1。柱的末端引入质谱仪,MS 条件:ESI+离子源,扫描模式为正离子,接口电压为4. 0 kV,接口温度为400 ℃,检测器电压为1. 76 kV,IG 真空度为2. 0×10-3 Pa,PG 真空度为1. 3×102 Pa,CID 气为2. 7×105 Pa,扫描范围为50~400 m·z-1。
1. 9 酶的提取与纯化
使用Sephadex G-50 对降解粗酶液进行凝胶柱色谱层析,流加速度1 mL·min-1,每10 mL 收集1 管,检测其酶活变化,取纯化酶液中活性较高的进行进一步纯化,重复凝胶柱色谱层析。酶活计算如下:
酶的活力:每24 h 每毫克酶降解UF 产生1 mg甲醛为1 个单位(U)
U =m1/m2
式中,U:酶的活力;m1:甲醛生成量,mg;m2:酶的质量,mg。
1. 10 凝胶渗透色谱分析
取纯化后的酶液进行真空冷冻干燥,将干燥后的粉末溶于水,过0. 22 μm 的滤膜,采用色谱柱(PL aquagel-OH 8 μm),检测器为RI 检测器、MLLS 激光光散射检测器,以聚乙二醇、聚环氧乙烷为标准品,采用PTEF 0. 22 μm 针筒过滤头,进样量50 μL,流速1 mL·min-1。
1. 11 统计分析
所有试验均重复3 次,实验结果取平均值并进行方差分析,运用单因素方差分析(ANOVA)中的最小显著性差异(LSD)法进行显著性差异分析。实验数据用Excel 2019 和Origin 2021 软件整理和作图,采用SPSS 20. 0 软件进行统计分析。
2 结果与分析
2. 1 P. rubens 23229 对UF 的降解性能
采用共培养体系研究P. rubens 23229 对热处理后UF 的降解作用。脲醛材料经热处理后,去除了在合成及存储过程中由热引起的易自动降解的短链小分子部分,因此热处理后的材料更便于研究UF 长链大分子的生物降解。从图1 可以看出,随着降解时间的延长,P. rubens 23229 以较为均一的速度对UF 进行降解,30 d 之后,材料失重率达到35%;对照组的无菌体系经过30 d 后,材料失重率为15%。结果表明,UF 在28 ℃实验条件下存在非生物性降解,加入P. rubens 23229 则可以在此基础上叠加生物降解作用,降解效率加倍。
2. 2 降解粗酶液性质研究
2. 2. 1 最适温度及温度稳定性
如图2 所示,在10~50 ℃范围内,降解酶随作用温度升高,降解效率增加,超过50 ℃,降解效率下降。说明该降解粗酶液最适工作温度为50 ℃左右。50 ℃条件下,降解36 h 后UF 失重率为11%。对比图1 中P. rubens 23229 菌与脲醛共培养条件下72 h 失重率只有3% 左右。分析其原因,P. ru⁃bens 23229 为青霉菌,其最适生长温度在28 ℃ 左右,此温度下酶处理UF 的失重率为50 ℃的18%,不能高效发挥其降解作用。由于温度的限制,单纯应用活菌共培养对材料进行降解不能发挥最优效果;然而,从共培养液中提取出来的粗酶液则可以通过提高作用温度达到UF 降解酶的最佳作用温度,提高降解效率,表明从菌培养液中提取降解酶是提高生物降解效率的1 个有效途径。
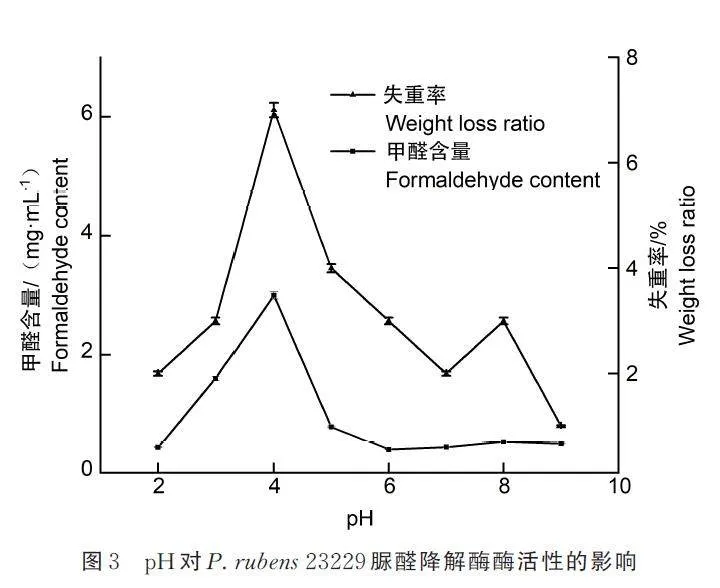
2. 2. 2 最适pH 及pH 稳定性
在初始pH 4. 0 条件下,降解酶液在36 h 降解了7% 的UF,同时产生了2. 998 mg·mL-1 含量的甲醛,其降解效率远高于其它pH 条件。如图3 所示,结果表明,酶工作的最适pH 在4. 0 左右。UF的降解包括将高分子主链降解为中间代谢物,然后彻底降解为甲醛和二氧化碳等小分子物质的过程,因此,从菌和材料共培养液中提取出来脲醛降解粗酶液中可能至少包含主链降解酶和中间代谢物降解酶2 类降解酶。从图3 可以看出,除在pH 4处同时产生失重率和甲醛含量2 个最高值峰外,在pH 8 左右失重率又有1 个小峰,但甲醛含量没有出现相应的小峰,说明此时UF 主链降解为可溶性物质,导致失重率增加,但是没有进一步降解为甲醛。进一步分析,可以得到以下结论:1)粗酶液中的主链降解酶在pH 4 活性最高,此外在pH 8 左右活性也比较高,它可以降解脲醛主链,使高聚合度的大分子不溶物含量下降,形成较多可溶性中间代谢物;2)将中间代谢物进一步降解为小分子物质,如甲醛的中间代谢物降解酶在pH 4 活性最高,在其它pH 值,包括pH 8 时活性较差;3)主链降解酶和中间代谢物降解酶协同作用降解UF,后者的低效率影响脲醛的彻底降解。
2. 3 不同酶降解性能比较
UF 以甲醛与尿素合成,连接键为酰胺键以及C-N,其中酰胺键结构类似于蛋白质中的肽键。从酰胺键及尿素水解的角度考虑,蛋白酶及脲酶也有可能降解UF,为此本实验对比了常用的蛋白酶胰蛋白酶、脲酶与P. rubens 23229 脲醛降解酶的降解性能。如图4 所示,胰蛋白酶处理组与蒸馏水对照组无显著差异,说明胰蛋白酶没有UF 降解活性。分析其原因,蛋白酶作用位点与蛋白质高级结构及氨基酸残基相关,虽然UF 主链连接键酰胺键类似于蛋白质肽键,但缺乏蛋白酶作用的高级结构与氨基酸残基,因此蛋白酶并不能水解UF。因此,P. rubens 23229 脲醛降解酶可能是菌体在脲醛环境中诱导产生的特异性分解酶。与蛋白酶不同,脲酶对UF 产生较好的降解效果,其失重率为P. rubens 23229 脲醛降解酶的20%,甲醛产生量为34%,这一结果说明P. rubens 23229 脲醛降解酶含有类似于脲酶作用方式的中间代谢物降解酶,它可以将主链降解下来的中间代谢物降解为甲醛等小分子产物并促进降解的顺利进行。为了进一步验证脲醛中间代谢物降解酶的作用,在P. rubens 23229 脲醛降解酶中加入脲酶进行UF 的降解,数据结果显示叠加脲酶后,其失重率是单独P. rubens 23229 脲醛降解酶的107%,甲醛产生量是其110%,说明脲酶的加入增强了中间代谢物降解酶的作用,提高了甲醛的产量,同时由于中间代谢物减少,产物抑制减少,主链降解酶的作用也相应增加,体现在失重率也相应增加。
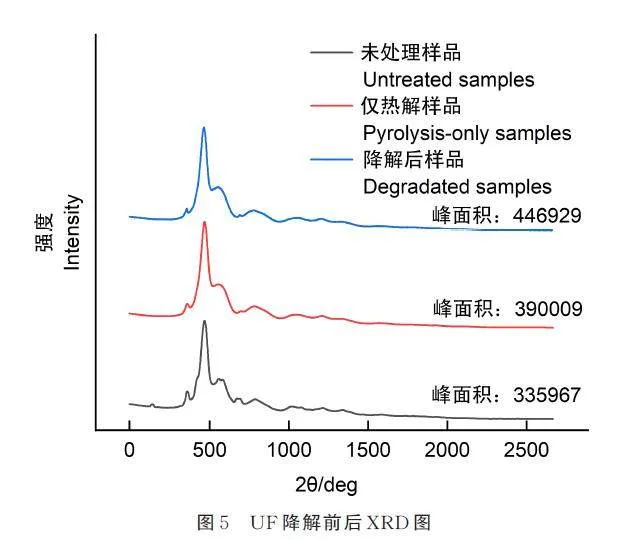
2. 4 降解前后UF 的X-射线衍射
P. rubens 23229 降解前后UF 的X-射线衍射图如图5,结晶度计算结果表明,未进行处理的样品结晶度为25. 18%,经热处理后样品的结晶度为26. 21%,说明100 ℃热处理导致非结晶区的一部分被降解去除,处理后的材料具有一定的耐热降解性。经热处理的UF 经P. rubens 23229 共培养后其结晶度进一步增加为29. 44%,失重率为35%,结合失重率及结晶度提高可以计算得出,UF 35% 的失重率中7% 为UF 结晶区,28% 为非结晶区[10]。非结晶区由于不规则排列,物理和化学性质相对不稳定,较易生物降解,高分子材料降解通常从非结晶区开始。结晶区排列紧密,通常较难生物降解。结果说明P. rubens 23229 降解酶除了降解非结晶区,对结晶区规则排列的脲醛分子也进行了一部分的降解。目前对高分子材料生物降解的报道多为对低结晶区的降解,如2016 年Science 期刊报道了对结晶度1. 6% 的聚对苯二甲酸乙二醇酯薄膜的生物降及降解酶研究[16]。26%以上结晶度UF 的生物降解尚未见报道。
2. 5 HPLC-MS 分析降解过程
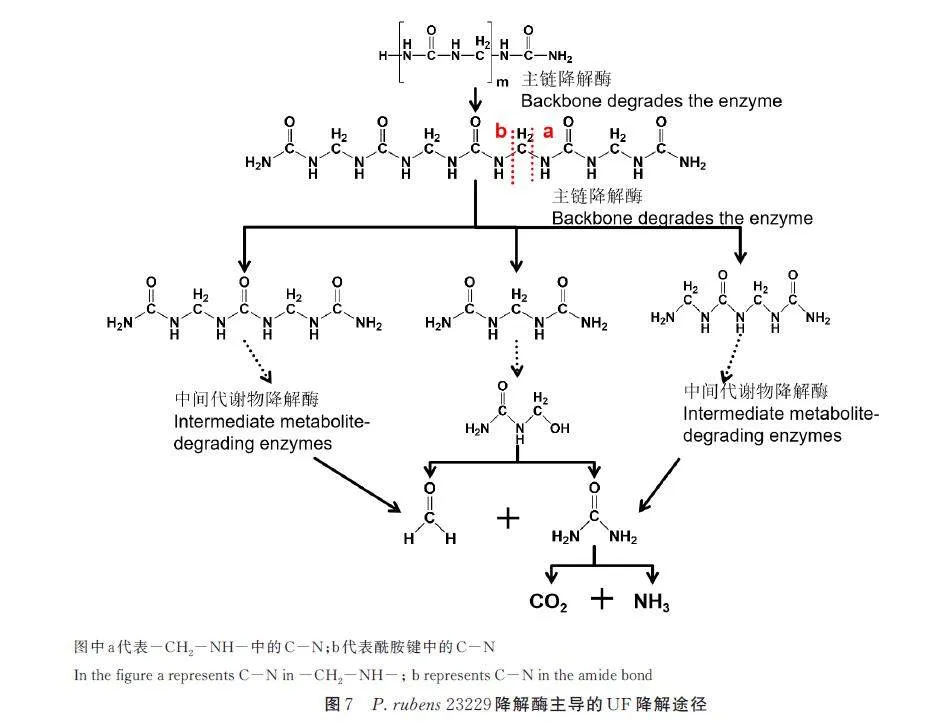
P. rubens 23229 降解粗酶液与UF 共培养后,取上清液进行HPLC-MS 分析,从图6 可以看出,与蒸馏水处理的对照组相比,上清液中出峰数量增加,新出现了③、⑦、⑧、⑨、⑩、⑪对应的峰,说明加入P. rubens 23229 降解粗酶液后可溶性代谢物种类增加,增加的代谢物为P. rubens 23229 酶切产物。此外,与对照组相比,酶处理组① 、② 、④、⑤、⑥对应的峰强度有大幅增加,说明酶的加入促进或叠加了原有UF 的理化降解。基于峰对应的质谱图分析出UF 降解形成的中间代谢产物,其中⑥号峰对应的m/z 值为205,对应的降解中间代谢物为二甲基三脲,⑨ 号峰对应的m/z 值为277,对应的降解中间代谢物为三甲基四脲。其它中间代谢物包括:亚甲基二脲、三甲基二脲、三甲基三脲、四甲基四脲、四甲基五脲等。基于中间代谢物的解析,分析在主链降解过程中降解酶的酶切位点主要涉及-CH2-NH-中C-N 的断裂(a)、酰胺键中C-N 的断裂(b)以及-CH2-NH-中脱氨基反应,基于中间代谢物及降解酶性质分析,结合Jahns 等[18] 报道的MDUase,绘制的P. rubens23229 酶主导的UF 降解过程见图7。
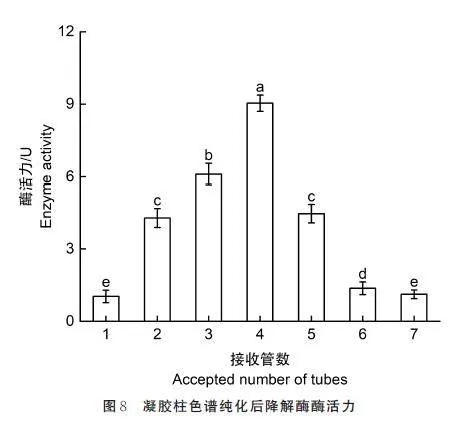
2. 6 酶的纯化
为了进一步研究酶的性质,应用凝胶柱色谱层析对P. rubens 23229 降解粗酶液进行多次纯化,结果如图8 所示。以降解酶液的酶活作为纯化物检测标准,最初粗酶酶活力为1. 02 U,层析纯化4 次后,酶活力达到28. 86 U,第5 次纯化后,酶活力为29. 96 U,与第4 次纯化后无显著差异。经过凝胶层析4 次后,酶活力较为稳定,纯化后酶的降解效率是粗酶的28 倍。
2. 7 凝胶渗透色谱
应用凝胶渗透色谱对纯化后的酶进行分子量的测定,结果见图9。纯化后的酶主要呈现2 个峰,第1 个峰(P1)相对分子量为220. 83 kD,PDI 值为1. 138;第2 个峰(P2)相对分子量为6. 89 kD,PDI值为1. 034。从图形来看,P1 仍然包含1 个以上的物质,可应用相应的凝胶柱进一步纯化。已有分子量分布结果说明在降解过程中,P. rubens 23229酶为至少由2 个酶组成的复合酶。
3 结论
以P. rubens 23229 及其分泌的酶对UF 进行降解,对降解酶及降解过程中中间代谢产物进行分析。凝胶渗透色谱分析显示P. rubens 23229 脲醛降解酶包含分子量为6. 89 kD 和220. 83 kD 的2个峰,表明P. rubens 23229 脲醛降解酶至少为2 个酶蛋白组成的复合酶,从功能可分为主链降解酶和中间代谢物降解酶,主链降解酶首先将脲醛主链降解为四甲基五脲、三甲基四脲等短链中间代谢物,然后由中间代谢物降解酶进一步降解为甲醛、二氧化碳等终产物,酶作用位点包括脲醛主链中亚甲基与尿素间的C-N、酰胺键等。进一步研究表明P. rubens 23229 脲醛降解酶中有1 种酶作用方式与脲酶相似,但降解主链酰胺键的酶与已有的蛋白酶作用方式不同。粗酶作用的最适温度为50 ℃,最适pH 为4,在最适条件下酶的降解效率远高于菌体最适生长条件的降解效率,因此以酶来进行生物降解效率更高。粗酶液经凝胶色谱纯化后酶活力提高了28 倍,其具体的组成与性质有待进一步研究。本研究对UF 的生物降解具有一定的参考价值。
参考文献
[1]郝志显,程艺艺,王乐乐,等.表面活性剂存在条件下脲醛树脂微球的合成反应过程[J].化学学报,2012,70(3):126-133.
Hao Z X,Cheng Y Y,Wang L L,et al. Synthesis of ureaformaldehyderesin microspheres under surfactant conditions[J].Acta Chimica Sinica,2012,70(3):126-133.
[2]向阳.新型有机高分子缓控释肥制备、降解机理以及应用效果研究[D].太原:中北大学,2018.
Xiang Y. Study on preparation,degradation mechanism andapplication effect of novel organic polymeric slow and controlledrelease fertilizers[D]. Taiyuan:North University of China,2018.
[3]Zhang B G,Jiang S Y,Du G B,et al. Polyurea-formaldehyderesin:a novel wood adhesive with high bonding performance andlow formaldehyde emission[J].The Journal of Adhesion,2021,97(5):477-492.
[4]Guo Y L,Shi Y Y,Cui Q X,et al. Synthesis of ureaformaldehydefertilizers and analysis of factors affecting theseprocesses[J].Processes,2023,11(11):3251.
[5]Cao L, Pizzi A, Zhang Q Y, et al. Preparation andcharacterization of a novel environment-friendly urea-glyoxalresin of improved bonding performance[J]. European PolymerJournal,2022,162:110915.
[6]Gao S S,Cheng Z H,Zhou X,et al. Unexpected role ofamphiphilic lignosulfonate to improve the storage stability ofurea formaldehyde resin and its application as adhesives[J].International Journal of Biological Macromolecules,2020,161:755-762.
[7]刘林,罗思义,方玲,等. 废旧脲醛树脂热处理研究进展[J].青岛理工大学学报,2021,42(1):126-133.
Liu L,Luo S Y,Fang L,et al. Research progress of heattreatment of waste urea-formaldehyde resin[J]. Journal ofQingdao University of Technology,2021,42(1):126-133.
[8]Samaržija-Jovanović S, Jovanović V, Petković B, et al.Hydrolytic,thermal,and UV stability of urea-formaldehyderesin/thermally activated montmorillonite nanocomposites[J].Polymer Composites,2020,41(9):3575-3584.
[9]王思佳,何品晶,邵立明,等. 树脂纽扣废物的焚烧污染特征[J].化工学报,2016,67(9):4004-4012.
Wang S J,He P J,Shao L M,et al. Characteristic pollutionsduring incineration of waste resin buttons[J]. CIESC Journal,2016,67(9):4004-4012.
[10]Ren T H,Wang Y,Wu N,et al. Degradation of ureaformaldehyderesin residues by a hydrothermal oxidationmethod into recyclable small molecular organics[J].Journal ofHazardous Materials,2022,426:127783.
[11]Wang G Y,Dai Y J,Yang H P,et al. A review of recentadvances in biomass pyrolysis[J]. Energy amp; Fuels,2020,34(12):15557-15578.
[12]Guo M Y,Yu S,Zhang S,et al. Nitrogen migration andconversion in chars from co-pyrolysis of lignocellulose derivedpyrolysis model compounds and urea-formaldehyde resinadhesive[J].Energies,2022,15(19):7221.
[13]Wang M,Xu S,Li S,et al. Isolation of formaldehydedegradingbacteria and the evaluation of the degradationcharacteristics[J] . Journal of Industrial and EngineeringChemistry,2019,75,224-229.
[14]周丽,Abdelkrim Y,姜志国,等.微塑料:生物效应、分析和降解方法综述[J].化学进展,2022,34(09):1935-1946.
Zhou L,Abdelkrim Y,Jiang Z G,et al. Microplastics:Areview of the biological effects,analysis,and degradationmethods[J]. Progress in Chemistry,2022,34(09):1935-1946.
[15]康宗利,刘爽,杨建,等.四环素降解菌的筛选及降解特性研究[J]. 山西农业大学学报(自然科学版),2022,42(2):79-88.
Kang Z L,Liu S,Yang J,et al. Screening and degradationcharacteristics of tetracycline degrading bacteria[J]. Journal ofShanxi Agricultural University (Natural Science Edition),2022,42(2):79-88.
[16]Yoshida S,Hiraga K,Takehana T,et al. A bacterium thatdegrades and assimilates poly(ethylene terephthalate)[J].Science,2016,351(6278):1196-1199.
[17]A M M A,Ying Y D T,Wayne K C,et al. Micro(nano)plastic pollution:The ecological influence on soil-plant systemand human health[J]. Science of the Total Environment,2021,788,147815-147815.
[18]Jahns T,Ewen H,Kaltwasser H. Biodegradability of ureaaldehydecondensation products[J]. Journal of Polymers andthe Environment,2003,11(4):155-159.
[19]王芳,胡培毅,向阳,等.高分子脲醛缓释肥料降解菌的筛选及降解酶性质初探[J]. 中国农业大学学报,2022,27(8):46-57.
Wang F,Hu P Y,Xiang Y,et al.Screening of polymeric ureaformaldehydeslow-release fertilizer degrading microorganismsand preliminary study on its degrading enzyme properties[J].Journal of China Agricultural University,2022,27(8):46-57.
(编辑:郭玥微)
基金项目:中央引导地方科技发展资金项目(YDZJSX2021B007);山西省重点研发计划项目(202302040201007);纳米功能复合材料山西省重点实验室开放基金(NFCM202003);山西省农业农村领域“六新”项目;山西省高分子缓控释肥料重点研发中心
猜你喜欢
中学化学(2024年1期)2024-05-26 13:20:27
生物技术通报(2020年12期)2020-12-21 09:20:00
腐植酸(2020年1期)2020-11-29 00:15:44
中国计算机报(2019年26期)2019-08-27 08:16:16
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07 11:14:32
中国科技博览(2017年39期)2017-09-07 09:14:31
中学生数理化·高二版(2017年2期)2017-04-19 16:29:54
中学化学(2016年12期)2017-02-05 17:24:23
核技术(2016年4期)2016-08-22 09:05:24
塑料制造(2016年5期)2016-06-15 20:27:39
