
烟酰胺调控大豆幼苗耐盐碱性的生理机制研究
2024-07-13 20:26:34张婉莹王怡张洋窦可欣邱桂林赵晓菊张琦
安徽农学通报 2024年12期
张婉莹 王怡 张洋 窦可欣 邱桂林 赵晓菊 张琦

摘要 为研究烟酰胺对大豆幼苗耐盐碱性的影响及其生理机制,本研究以大豆品种合丰50为试验材料,通过对盐碱胁迫下大豆幼苗喷施不同浓度梯度的烟酰胺进行处理,分析了不同处理下大豆幼苗的生长指标和生理指标以及烟酰胺酶基因(NIC1)的表达量。结果表明,一定浓度的烟酰胺处理可以提高盐碱胁迫下大豆幼苗的生长指标株高、根长和生物量,同时提高叶片中生理指标渗透调节物质的含量、抗氧化酶活性和光合色素含量,本试验条件下,烟酰胺溶液质量浓度以50 mg/L处理时效果最佳。烟酰胺处理后,大豆叶片中NIC1基因的表达量有所上调,说明烟酰胺酶基因的上调表达能够提高大豆的耐盐碱能力。研究结果为烟酰胺调控大豆幼苗耐盐碱性的生理机制研究提供参考。
关键词 大豆;烟酰胺;盐碱胁迫;光合色素;基因表达
中图分类号 S565.1 文献标识码 A
文章编号 1007-7731(2024)12-0038-07
Physiological mechanism of nicotinamide regulating salt and alkaline
tolerance in soybean seedlings
ZHANG Wanying WANG Yi ZHANG Yang DOU Kexin QIU Guilin ZHAO Xiaoju ZHANG Qi
(Daqing Normal University, Daqing 163712, China)
Abstract In order to explore the effects of nicotinamide on salt and alkaline tolerance of soybean seedlings and its physiological mechanism,this paper used soybean Hefeng 50 as experimental material to analyze the growth indicators and physiological indicators of soybean seedlings under different concentrations of nicotinamide sprayed under salt and alkali stress, as well as the expression level of nicotinamide enzyme genes (NIC1) under different treatments. The results showed that a certain concentration of niacinamide treatment could increase the plant height, root length, and biomass of soybean seedlings under salt alkali stress, while also increasing the content of osmoregulatory substances, antioxidant enzyme activity, and photosynthetic pigment content in leaves. Under the experimental conditions,the best effect was achieved when the mass concentration of niacinamide solution was 50 mg/L. After nicotinamide treatment, the expression level of NIC1 gene in soybean leaves was upregulated, indicating that the upregulation of nicotinamide enzyme gene could improve the salt alkali tolerance of soybean. The research results provided a reference for the physiological mechanism of nicotinamide regulating salt and alkaline tolerance in soybean seedlings.
Keywords soybeans; nicotinamide; salt alkali stress; photosynthetic pigment; gene expression
大豆富含蛋白质、脂肪、碳水化合物以及多种人体所需的氨基酸等,是重要的粮油饲菜兼用作物之一,其籽粒中的蛋白质含量占籽粒干物质的40%左右,部分高达50%,是重要的植物蛋白来源之一[1]。目前,部分耕地土壤盐碱化问题影响着粮食生产[2-3]。大豆在盐碱胁迫土地生长时,幼苗植株的生长可能受到抑制,进而降低了地上部和地下部物质量积累,最终表现为植株矮小等[4]。盐碱胁迫使土壤中无机离子含量增加,无机离子会破坏细胞的生物膜结构,可能使植物的渗透调节系统失调,严重失调可导致植物死亡[5]。植物可以通过酶类抗氧化系统来清除体内在盐碱胁迫下产生的过量活性氧,严媚欣等[6]研究表明,在盐碱胁迫下,罗布麻幼苗叶片中抗氧化酶活性随胁迫处理浓度的增大而上升。植物细胞中的叶绿体是受盐碱胁迫影响较大的细胞器之一,Jia等[7]研究发现,盐碱混合胁迫可降低植株对叶绿素必须金属离子的吸收,从而抑制海棠根系合成叶绿素,导致叶片内叶绿素a和叶绿素b含量下降。
烟酰胺是维生素B3的活性物质,维生素与植物激素有许多相似的作用,均可以促进植物生长[8]。烟酰胺处理可明显提高小麦的生长指标,且其叶片中叶绿素含量和α-淀粉酶活性也有明显提高[9]。利用不同浓度外源烟酰胺处理菜豆种子,50 mg/L烟酰胺浸种的菜豆种子的发芽率明显提高[10]。烟酰胺本身在碱性条件下并不稳定,会水解生成烟酸,呈酸性,具有抗氧化和促进根系发育的效果,有助于大豆在盐碱地的生长。同时,烟酰胺还可以与盐碱地中的碱性盐发生反应,使土壤pH值降低,不仅可以改善大豆的生长条件,还可以提高移栽苗成活率。烟酰胺可通过植物根、茎和叶的吸收来提高植物体内辅酶Ⅰ和辅酶Ⅱ活性,进而促进植株生长和根的形成[11],这在农业生产中十分重要。杨萍[12]利用维生素B3对盐胁迫后的黄瓜进行处理,发现一定浓度的维生素B3能够促进盐碱胁迫下黄瓜种子的萌发和幼苗的生长。植物中烟酰胺酶的活性较高,常被用于合成烟酰胺腺嘌呤二核苷酸(Nicotinamide adenine dinucleotide,NAD)与烟酰胺腺嘌呤二核苷酸磷酸(Nicotinamide adenine dinucleotide phosphate,NADP),并参与植物生长发育的调节过程,可以增强作物的抗逆性,提高作物产量和改善品质。Wang等[13]发现拟南芥烟酰胺酶基因AtNIC1是拟南芥中烟酰胺转化成烟酸主要的酶之一,携带该基因突变的拟南芥植株的NAD和NADP水平较低,并对外源添加的NaCl表现出异常敏感。
幼苗期是大豆生长的关键期,也是对盐碱胁迫比较敏感的时期,提高幼苗期的耐盐碱性对大豆整个生长期具有重要作用[14-15]。本试验以生长至一节期(V1)的大豆幼苗为试验样本,采用盐碱混合胁迫下叶面喷施不同浓度的烟酰胺的方法,通过分析幼苗生长情况和生理指标,以及NIC1基因的表达变化趋势,探究外源施用烟酰胺缓解大豆幼苗盐碱胁迫的生理机制,为利用烟酰胺提高大豆耐盐碱性提供参考。
1 材料与方法
1.1 试验材料
大豆品种为合丰50,由黑龙江省农业科学院佳木斯分院提供。烟酰胺等试剂(分析纯)购自国药试剂公司。试剂盒购自天根生化科技有限公司。
1.2 试验方法
采用塑料盆(长×宽×高为23 cm×20 cm×15 cm)栽植试验,试验土壤基质按栽培土∶腐殖土∶砂土为2∶1∶1比例混合而成。选取籽粒饱满、大小均匀一致的大豆种子,每盆种植10粒。在幼苗长至V1期时进行盐碱处理,采用NaCl和NaHCO3以摩尔质量比1∶1混合配制浓度100 mmol/L的盐碱混合溶液,在大豆生长的V1期和V2期用100 mmol/L盐碱混合溶液各处理一次。
本试验设置7组处理,每组处理设3次重复。CK,清水;SA,100 mmol/L盐碱混合溶液+0 mg/L烟酰胺溶液;N1,100 mmol/L盐碱混合溶液+5 mg/L烟酰胺溶液;N2,100 mmol/L盐碱混合溶液+10 mg/L烟酰胺溶液;N3,100 mmol/L盐碱混合溶液+25 mg/L烟酰胺溶液;N4,100 mmol/L盐碱混合溶液+50 mg/L烟酰胺溶液;N5,100 mmol/L盐碱混合溶液+100 mg/L烟酰胺溶液。
1.3 测定指标及方法
1.3.1 生长指标测定 在叶面喷施烟酰胺溶液后的第1、3、5和7 d分别取样,每处理各取3株生长均匀的大豆幼苗,将子叶节作为分段点,以地上和地下两侧为标准,分割成两个部分,分别测定其株高、根长以及地上和地下部分鲜重,再置于120 ℃温度下杀青10 min,80 ℃烘干,待至恒重后称量地上和地下干物质量。
1.3.2 渗透调节物质含量测定 参照李合生[16]的方法测定各处理叶片中脯氨酸、可溶性糖和可溶性蛋白含量。
1.3.3 抗氧化酶活性测定 参照生林山等[17]的方法测定各处理叶片中超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)活性。
1.3.4 光合色素含量测定 采用分光光度法[16]测定各处理叶片中叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量。
1.3.5 NIC1基因表达水平测定 选取大豆三出复叶0.1 g,使用总 RNA提取试剂盒进行RNA提取,反转录后获得cDNA,稀释10倍后备用,设计荧光定量PCR引物,以大豆Actin为内参基因,按照FP-205试剂盒进行操作,并在实时荧光定量PCR仪(ABI 7500)中进行反应。计算基因的相对表达量。
1.4 数据处理
用Microsoft Excel 2016软件进行数据整理及分析,采用单因素(One-Way ANOVA)和Duncan法进行方差分析和多重比较(α=0.05)。利用Excel 2016软件作图,相关数据以平均值±标准差表示。
2 结果与分析
2.1 生长指标
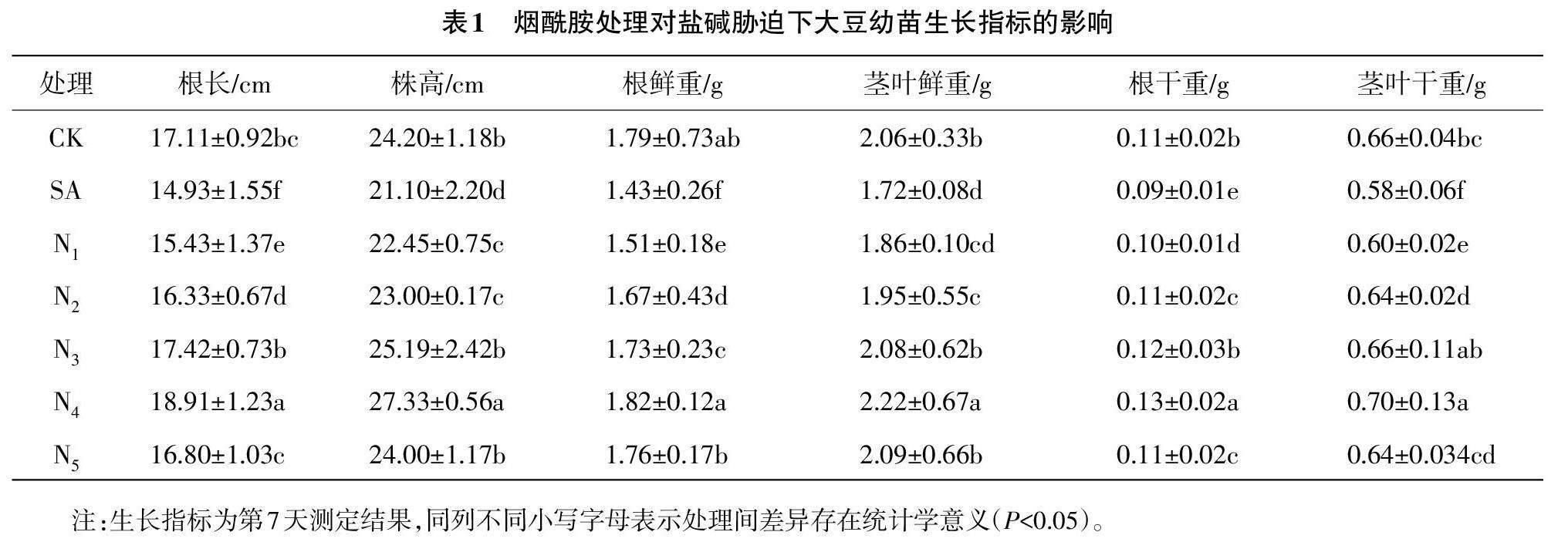
由表1可知,盐碱胁迫处理(SA)的大豆幼苗各项生长指标均低于对照组(CK),随着叶面喷施烟酰胺浓度的增加,大豆幼苗的根长、株高和生物量情况表现为先升高后降低的趋势,以N4处理的各项生长指标表现较好,说明50 mg/L烟酰胺处理(N4)能够有效缓解盐碱胁迫对大豆幼苗生长的抑制作用,且在一定程度上可以促进大豆幼苗生长。与SA处理相比,N4处理的大豆幼苗的根长、株高、根鲜重、茎叶鲜重、根干重和茎叶干重明显增加(P<0.05)。N1、N2处理的大豆幼苗的各项生长指标均明显低于CK(P<0.05),说明叶面喷施低浓度的烟酰胺溶液对盐碱胁迫下大豆幼苗生长的促进作用较弱。
2.2 渗透调节物质
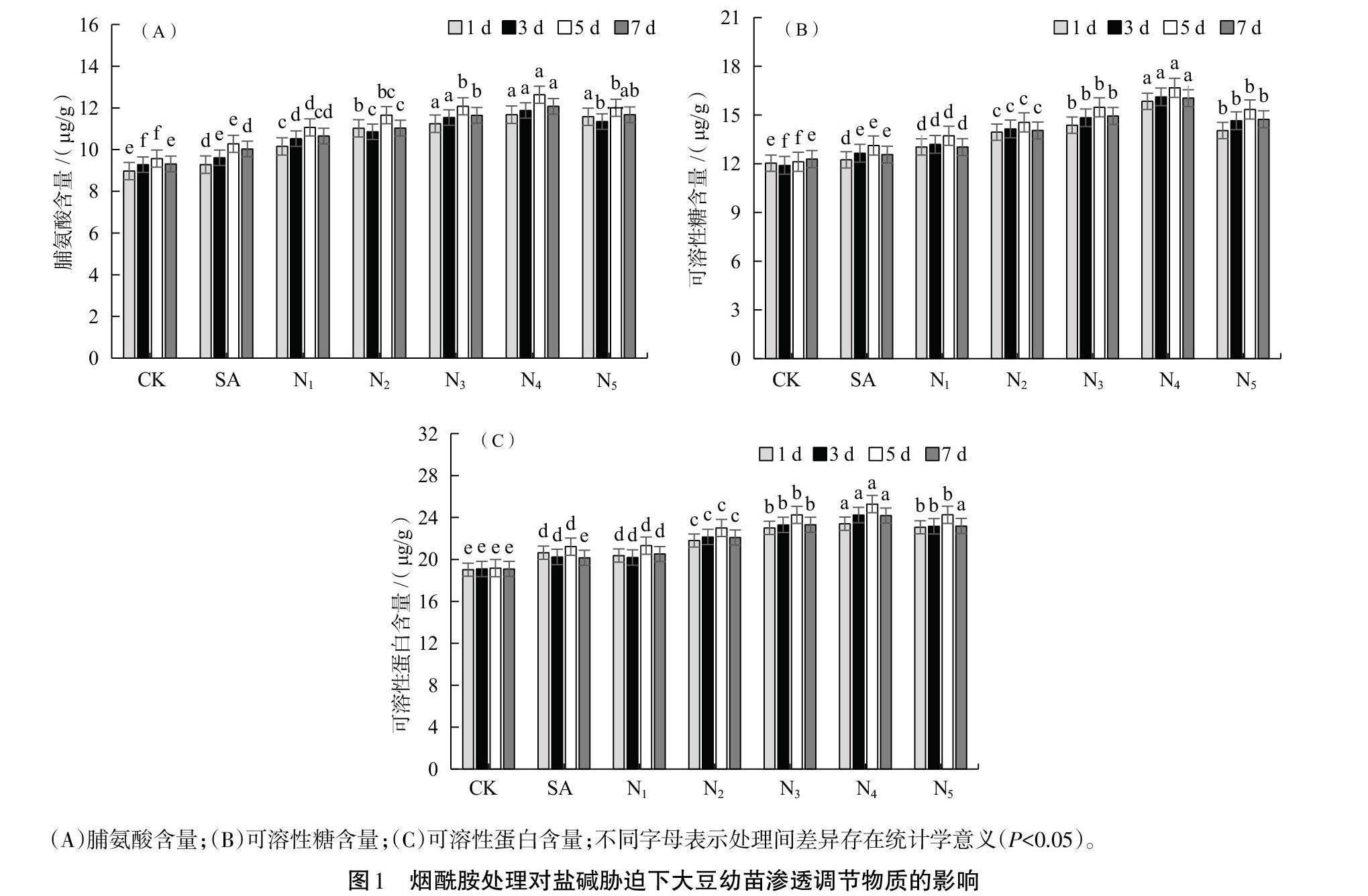
烟酰胺对盐碱胁迫下大豆幼苗渗透调节物质的影响如图1所示。SA处理的大豆幼苗叶片中脯氨酸、可溶性糖和可溶性蛋白含量与CK相比有所增加。随着喷施烟酰胺浓度的增加,大豆幼苗叶片的3种渗透调节物质含量呈现先上升后下降的趋势,且均在N4处理时含量达到最高。从不同取样天数来看,整体表现为在第5天取样时,大豆叶片中积累的3种渗透调节物质的含量较高。即N4处理后第5天取样的大豆叶片中脯氨酸、可溶性糖和可溶性蛋白含量最高,分别为12.633、16.674和25.280 μg/g,较SA处理第5天取样时分别增加了22.92%、21.29%和16.07%。大豆叶片中的渗透调节物质含量在N5处理下均有所降低,表明一定浓度的烟酰胺处理可使大豆幼苗叶片中渗透调节物质含量升高,进而缓解盐碱胁迫对大豆的伤害,但施用烟酰胺浓度过高可能导致缓解效果变差。
2.3 抗氧化酶活性
烟酰胺对盐碱胁迫下大豆幼苗抗氧化酶活性的影响如图2所示。在盐碱胁迫条件下,大豆叶片中的SOD、POD和CAT活性均呈现上升趋势,而APX活性呈现不同程度的下降趋势;随着大豆幼苗叶片喷施不同浓度的烟酰胺,叶片中各种抗氧化物酶活性的变化程度不同。在N4处理下,SOD、POD、CAT和APX的活性最高;在N4处理后第5天,SOD、CAT和APX活性达到最大值,分别比SA处理增加28.17%、26.26%和23.05%;POD活性在N4处理后第3天最大,比SA处理增加22.13%。随着烟酰胺浓度的增加以及取样天数的延长,大豆幼苗叶片中4种抗氧化物酶活性均存在不同程度的降低,这表明过高浓度的烟酰胺处理可能会减弱对大豆盐碱胁迫的缓解作用。
2.4 光合色素含量
烟酰胺处理对盐碱胁迫下大豆幼苗光合色素含量的影响如图3所示。SA处理后的大豆叶片中光合色素的含量与CK相比明显下降(P<0.05),说明盐碱胁迫对大豆的光合作用存在抑制作用。施用不同浓度烟酰胺后,大豆叶片中光合色素的含量较SA处理相比整体呈现上升趋势,即烟酰胺的施用在一定程度上能够提高盐碱胁迫下大豆的光合能力。
整体来看,大豆叶片中叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量在N4处理后第5天最高,分别为3.965、1.327、5.292和1.297 mg/g FW。与SA处理相比分别增加20.00%、20.31%、20.08%和21.10%,差异均存在统计学意义(P<0.05)。
2.5 叶片中NIC1基因表达水平
由图4可知,在盐碱胁迫处理下,NIC1基因相对表达量较CK组明显降低(P<0.05),而喷施烟酰胺处理的大豆幼苗叶片,NIC1基因相对表达量有不同程度的提高。在N4处理时,NIC1基因相对表达量最高,较SA组明显增加(P<0.05)。说明外源烟酰胺的施用能够提高烟酰胺酶的活性,促进烟酰胺的转化与再生。
3 结论与讨论
土壤盐碱化在一定程度上会影响大豆幼苗的生长发育,主要表现为抑制根系的伸长,使植株变得矮小且短粗,极大降低了植株的鲜重、干重和叶面积,并且明显抑制植株生物量的积累[18-19]。烟酰胺是维生素B3的活性物质,研究表明外源施用烟酰胺可以提高水稻[20]、菜豆[10]和小麦[9]等幼苗的株高、根长、植株鲜重和干重。在植物受到非生物胁迫时,提高体内烟酰胺含量可以有效促进植物的生长发育。杨萍[12]利用烟酰胺对盐胁迫后的黄瓜进行浸种处理,发现一定浓度的烟酰胺能够促进盐胁迫下黄瓜种子的萌发以及幼苗的生长。本研究结果与此一致,盐碱胁迫下,大豆幼苗叶片喷施50 mg/L烟酰胺溶液能够缓解盐碱胁迫对大豆幼苗的伤害,幼苗的各项生长指标与盐碱胁迫处理相比均有所增长。
植物细胞中的脯氨酸、可溶性糖和可溶性蛋白是植物体内渗透调节系统的主要组成部分,与植物耐盐碱性密切相关[21-22]。Khurshid等[23]研究表明,干旱胁迫下,在小麦花期土壤中施用0.492 g/L的烟酰胺能够提高叶片中的脯氨酸含量。本研究对盐碱胁迫下大豆幼苗叶片喷施不同浓度的烟酰胺溶液发现,施用50 mg/L的烟酰胺溶液能够有效提高叶片中渗透调节物质的含量,对抵抗盐碱胁迫的效果较好。
NAD的积累能够降低植物体内线粒体呼吸,提高活性氧(ROS)能量利用率,进而提高植物对非生物胁迫的耐受性[24]。当植物在逆境条件下生长时,细胞中ROS的主要生产者是NADPH氧化酶,因此NADPH的增加会抑制ROS的产生,提高抗氧化酶的活性[25]。Farooq等[26]研究表明,在干旱胁迫下施用烟酰胺后,大麦植株的CAT、SOD和POD活性增加;干旱处理下小麦植株的CAT、SOD、POD和APX等抗氧化酶的生物合成和积累明显增加[23]。本研究发现,在盐碱胁迫处理下施用烟酰胺能够使大豆叶片中抗氧化酶活性增强,与上述研究结果相似,说明在非生物胁迫下施用烟酰胺能够提高植物细胞中的抗氧化酶活性,从而抵抗非生物胁迫对细胞产生伤害。
盐碱胁迫可能会破坏植物中叶绿体的结构,打破叶绿素代谢平衡,导致叶绿素含量下降,阻碍光合磷酸化过程,最终减弱植物光合作用[27]。研究表明,烟酰胺可以提高多种作物的光合速率,如小麦[9]、辣椒[28]等,这可能与叶绿素浓度升高、叶片超微结构改变以及1,5-二磷酸核酮糖羧化酶活性增强等因素有关[29]。本研究发现,在盐碱胁迫下施用一定浓度的烟酰胺能够增加大豆叶片中的光合色素含量,表明外源添加烟酰胺可以有效缓解盐碱胁迫对叶绿体造成的损伤。
NIC1基因是烟酰胺合成NAD途径的关键酶基因[30],本研究结果表明,叶面喷施50 mg/L烟酰胺溶液能够增加细胞中的烟酰胺含量,使烟酰胺酶NIC1基因的表达水平上调,从而促进NAD的合成,NAD与NADP可以通过磷酸化作用相互转变。NAD和NADP在细胞代谢中参与糖酵解、磷酸戊糖途径和乙醛酸循环等过程,为细胞积累淀粉、蛋白质和脂类等物质,具有促进大豆生长的作用。
本研究通过对盐碱胁迫下大豆幼苗喷施不同浓度梯度的烟酰胺,分析了不同处理下大豆幼苗的生长指标、生理指标以及烟酰胺酶基因(NIC1)的表达量。结果表明,外源施用一定浓度的烟酰胺可以提高大豆植株的生物量,促进细胞中渗透调节物质的积累,大豆叶片中抗氧化酶活性、光合色素含量提高,大豆叶片中NIC1基因表达水平上调。综合来看,外源施用50 mg/L的烟酰胺溶液能够有效提高大豆幼苗抵抗盐碱胁迫的能力。但烟酰胺在大豆体内的转运和再生途径比较复杂,未来可对大豆体内的烟酰胺循环和再生通路对其他植物激素水平调节在抵抗非生物胁迫方面进行更深入的探讨。
参考文献
[1] 杨文英,于晓波,安建刚,等. 2011—2022年四川省审定春、夏大豆品种主要性状演变趋势分析[J]. 中国油料作物学报,2023,45(5):935-946.
[2] 刘建新,刘瑞瑞,刘秀丽,等. 不同时期喷施NaHS对盐碱胁迫下裸燕麦叶片渗透调节物质和抗氧化活性的影响[J]. 生态学杂志,2021,40(11):3620-3632.
[3] 马晓军,金峰学,杨姗,等. 作物耐盐碱数量性状基因座(QTL)定位[J]. 分子植物育种,2015,13(1):221-227.
[4] 毛恋,芦建国,江海燕. 植物响应盐碱胁迫的机制[J]. 分子植物育种,2020,18(10):3441-3448.
[5] 韩毅强,高亚梅,杜艳丽,等. 大豆耐盐碱种质资源鉴定[J]. 中国油料作物学报,2021,43(6):1016-1024.
[6] 严媚欣,白茹,崔金霞,等. 混合盐碱胁迫对罗布麻生理特性的影响[J]. 农业科技通讯,2021(8):153-157.
[7] JIA X M,WANG H,SVETLA S,et al. Comparative physiological responses and adaptive strategies of apple Malus halliana to salt,alkali and saline-alkali stress[J]. Scientia horticulturae,2019,245:154-162.
[8] 唐瑞,吴瑜. 维生素对小麦生长及生理功能的调节作用研究进展[J]. 应用与环境生物学报,2006,12(6):869-873.
[9] 周浩,金如昌,孟恒君,等. 烟酰胺对小麦萌发及幼苗生长的影响[J]. 安徽化工,2021,47(5):61-62,68.
[10] 李振轮,李鑫强,朱培. 维生素浸种和电场处理对菜豆生长发育初期的影响[J]. 西南大学学报(自然科学版),2013,35(5):145-151.
[11] VON W N,KLAIR S,BANSAL S,et al. Nicotianamine chelates both FeIII and FeII. Implications for metal transport in plants[J]. Plant physiology,1999,119(3):1107-1114.
[12] 杨萍. 外源维生素对盐胁迫下黄瓜种子萌发及植株生理特性的影响[D]. 重庆:西南大学,2010.
[13] WANG G D,PICHERSKY E. Nicotinamidase participates in the salvage pathway of NAD biosynthesis in Arabidopsis[J]. The Plant journal,2007,49(6):1020-1029.
[14] 胡雯恬. 大豆幼苗性状耐盐碱胁迫能力优异等位变异的发掘[D]. 哈尔滨:黑龙江大学,2022.
[15] 侯鹏浩,杨万明,杜维俊,等. 不同程度盐胁迫对大豆苗期生物量及生理指标的影响[J]. 大豆科学,2020,39(3):422-430.
[16] 李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[17] 生林山,韩佳一,盛雨婷,等. 外源(Z)-3-己烯基乙酸酯缓解小麦盐胁迫的作用机制[J]. 植物生理学报,2022,58(7):1263-1274.
[18] 余明龙,黄露,郑殿峰,等. 外源调环酸钙对盐碱胁迫下大豆幼苗生长及生理特性的影响[J]. 生态学杂志,2022,41(4):683-692.
[19] 赵怀玉,林鸿宣. 植物响应盐碱胁迫的分子机制[J]. 土壤与作物,2020,9(2):103-113.
[20] CHEN W,WANG W S,PENG M,et al. Comparative and parallel genome-wide association studies for metabolic and agronomic traits in cereals[J]. Nature communications,2016,7:12767.
[21] CALERO HURTADO A,CHICONATO D A,DE MELLO PRADO R,et al. Different methods of silicon application attenuate salt stress in sorghum and sunflower by modifying the antioxidative defense mechanism[J]. Ecotoxicology and environmental safety,2020,203:110964.
[22] GHOULAM C,FOURSY A,FARES K. Effects of salt stress on growth,inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars[J]. Environmental and experimental botany,2002,47(1):39-50.
[23] KHURSHID N,BUKHARI M A,AHMAD T,et al. Exogenously applied nicotinic acid alleviates drought stress by enhancing morpho-physiological traits and antioxidant defense mechanisms in wheat[J]. Ecotoxicology and environmental safety,2023,263:115350.
[24] GONZALEZ-GUZMAN M,PIZZIO G A,ANTONI R,et al. Arabidopsis PYR/PYL/RCAR receptors play a major role in quantitative regulation of stomatal aperture and transcriptional response to abscisic acid[J]. The Plant cell,2012,24(6):2483-2496.
[25] KAWAI S,MURATA K. Structure and function of NAD kinase and NADP phosphatase:key enzymes that regulate the intracellular balance of NAD(H) and NADP(H)[J]. Bioscience,biotechnology,and biochemistry,2008,72(4):919-930.
[26] FAROOQ T H,BUKHARI M A,IRFAN M S,et al. Effect of exogenous application of nicotinic acid on Morpho-physiological characteristics of Hordeum vulgare L. under water stress[J]. Plants,2022,11(18):2443.
[27] 张景云,缪南生,白雅梅,等. 盐胁迫下二倍体马铃薯叶绿素含量和抗氧化酶活性的变化[J]. 作物杂志,2014(5):59-63.
[28] 刘永军,赵宗芸,杨晓玲,等. 三种维生素对甜椒主要生育和生理生化指标的影响[J]. 河北职业技术师范学院学报,2002,16(2):32-34,38.
[29] SHEMI R,WANG R,GHEITH E S M S,et al. Effects of salicylic acid,zinc and glycine betaine on morpho-physiological growth and yield of maize under drought stress[J]. Scientific reports,2021,11(1):3195.
[30] GAKI?RE B,HAO J F,DE BONT L,et al. NAD+Biosynthesis and signaling in plants[J]. Critical reviews in plant sciences,2018,37(4):259-307.
(责编:李 媛)
猜你喜欢
今日农业(2022年16期)2022-11-09 23:18:44
中国化肥信息(2022年5期)2022-08-30 01:58:26
今日农业(2021年20期)2021-11-26 01:23:56
今日农业(2021年14期)2021-10-14 08:35:34
中国科技博览(2016年24期)2016-12-28 23:53:07
中国实用医药(2016年30期)2016-12-28 16:54:00
中国民族民间医药·上半月(2016年10期)2016-11-19 11:08:04
中国科技博览(2016年22期)2016-11-01 13:58:50
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:18
药学与临床研究(2014年3期)2014-03-06 13:35:35
