
樱亚属分类学研究进展
2024-06-16 00:00:00伊贤贵李蒙王贤荣
南京林业大学学报(自然科学版) 2024年3期
关键词:分类学



摘要:回顾与整理了樱亚属分类学的研究历史,对各主要樱亚属分类系统进行了概述。总结樱亚属植物实验分类学相关研究,认为形态学标记依然是樱亚属分类的基础;SSR标记研究可以为樱亚属种间及种内关系讨论提供证据;DNA序列标记在樱亚属分类研究中成果显著,基本厘清了樱亚属系统发育关系。整合樱亚属系统发育研究与地理分布最新数据,描绘了樱亚属系统发育主要框架,推测喜马拉雅山区至中国西南部或地中海地区为樱亚属起源地,总结了樱亚属迁移扩散的主要线路。结合文献与标本资料以及野外调查与最新研究结论,列出世界樱亚属物种名录共计76种(含11变种),并对樱亚属植物主要分布区及类群进行了统计;对山樱花复合群与大叶早樱复合群及其他争议类群,提出了分类处理观点。综合分析了樱亚属系统分类学研究现状,认为樱亚属复合体及存疑类群,以及中西部地区资源调查与分类整理等需要更深入的研究;大尺度下的多物种、多群体与多组学的组合探究,是樱亚属系统分类学研究的重要方向。
关键词:樱亚属;分类学;系统发育;复合群
中图分类号:S685.99;S79 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)03-0046-12
A review on the taxonomy study of Prunus subgen. Cerasus (Mill) A. Gray
YI Xiangui, LI Meng, WANG Xianrong*
(Co-Innovation Center for the Sustainable Forestry in Southern China, College of Life Sciences, Cerasus Research Center, Nanjing Forestry University, 210037 Nanjing, China)
Abstract:Reviewed and summarized the research history of the taxonomy of the subgenus Prunus subgen. Cerasus, provided an overview of major classification systems of the subgen. Cerasus. Summarized the advances in experimental taxonomy studies on the subgen. Cerasus, It is found that morphological markers play an important role in the classification of subgen. Cerasus. SSR marker studies can provide evidence for discussions on interspecific and intraspecific relationships within the subgen. Cerasus, DNA sequence markers have made important achievements in the study of subgen. Cerasus classification, essentially clarifying the phylogenetic relationships within the subgen. Cerasus. Integrated the latest data on phylogeny and geographic distribution of the subgen. Cerasus, outlined the main framework of the phylogeny of the subgen. Cerasus, speculation that the origin of this subgenus is from the Himalayan region to southwestern China or the Mediterranean region, summarized the main migration routes of the subgen. Cerasus. By combining literature and specimen data, field surveys and recent research conclusions, a total of 76 species of the subgen. Cerasus (including 11 varieties) were listed in the world species catalog, and statistical analysis was conducted on the main distribution areas and groups of the subgen. Cerasus; proposed classification viewpoints for the P. serrulata complex, P. subhirtella complex, and other controversial groups. A comprehensive analysis of the status of the taxonomy of the subgen. Cerasus, suggesting that more in depth research is needed on the complex and controversial groups within the subgen. Cerasus, as well as resource surveys and classification in the central and western regions; exploring combinations of multiple species, populations, and omics at a large scale is an important direction for the taxonomy research of the subgen. Cerasus.
Keywords:Prunus subgen. Cerasus; taxonomy; phylogeny; comples
樱亚属[Prunus subgen. Cerasus (Mill.) A. Gray],隶属蔷薇科(Rosaceae)李属(Prunus L.),自然分布于北半球温暖地带,集中分布于中国西部、西南部和东部以及日本和朝鲜半岛,是温带与亚热带森林的表征类群。樱亚属植物种类多、分布广且变异幅度大,因其具有极高的观赏价值,为世界著名观赏花木。然而,关于樱亚属的分类地位和物种划分,众多分类学者意见很不一致,樱亚属与樱属(Cerasus Mill.)的分类观点长期并行采用,加之不断有新种/变种的报道,樱亚属分类学研究依然存在较多的疑问与争议。笔者系统梳理樱亚属分类学的研究历史,总结其系统分类学研究进展,以期为未来樱亚属系统分类学研究明晰方向。
1 樱亚属分类学历史
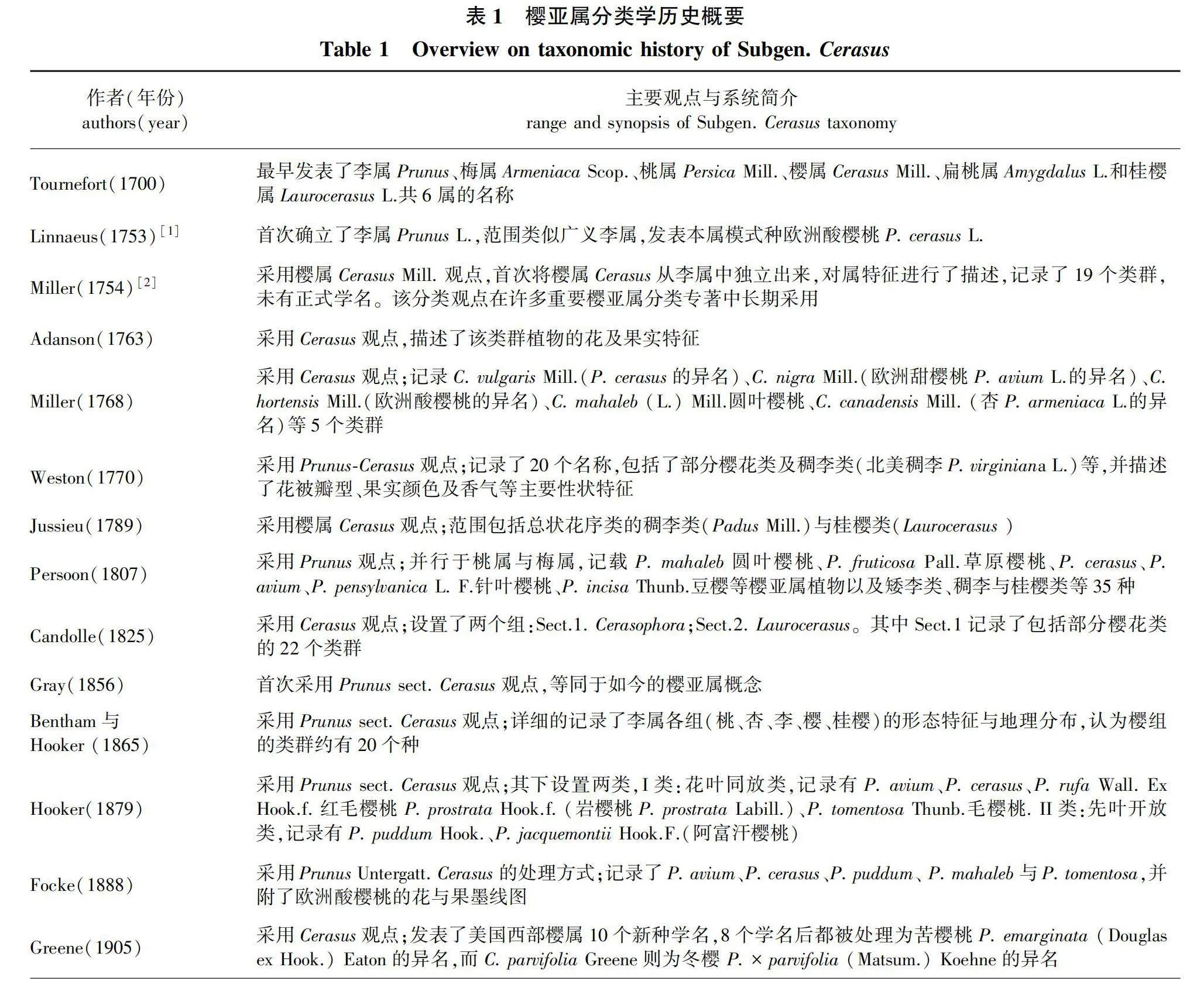
樱亚属分类学历史十分悠久,1753年,Linnaeus[1]在Species Plantarum中确立了李属(Prunus L.),范围与现今广义李属相似,包括了李、桃、杏、梅与樱等。1754年,Miller[2]首次从李属中独立出樱属(Cerasus Mill.),范围包括单芽樱类和3芽樱类。1756年, Gray[3]采用李属樱组(Sect. Cerasus)的观点,分类范畴等同于如今的樱亚属。1912 年, Koehne[4]在专著Plantae Wilsonianae中,采用Prunus subgen. Cerasus 观点, 把其发表的35 个中国产新种分成2 个Grex.,即Grex. I. Typocerasus 和Grex. II. Microcerasus,其概念对应于今天的樱亚属和矮生樱亚属,自此确立了樱亚属的基本范围。樱亚属分类观点长期未得到统一,不同时期的分类系统概览如表1所示[5]。
樱亚属植物具有极高的观花与果用价值,因其在世界范围内广泛分布与栽培,受到植物分类学者的长期关注。到底是采用李属(Prunus)还是采用独立樱属(Cerasus)的观点,传统分类未达成共识。近年来随着分子技术手段的应用,更多的证据表明典型樱亚属(Subgen. Cerasus)为一单系,而矮生樱亚属(Subgen. Microcerasus)与李属的关系更近。笔者借鉴了《中国植物志》与Flora of China关于典型樱亚属 Subgen. Cerasus的观点与范畴[11,14-17],梳理并总结了樱亚属分类历史及其代表性樱亚属分类系统。
2 樱亚属实验分类学研究进展
樱亚属分类研究长期基于形态学手段,随着微形态学、细胞学、生物化学和分子生物学等先进方法的应用,多种实验手段与多学科途径的综合分析,为樱亚属的分类与演化提供了更为坚实和客观的科学证据。
2.1 形态学标记研究进展
形态多样性与变异研究是解开种群分化及物种界定的重要手段。Chang等[18]学者通过蜡叶标本数据,细致分析了山樱花复合体的表型变异,为其分类处理提供了重要参考,将山樱花复合体处理为:C. serrulata var. serrulata. f. serrulata (Lindl.) G. Don、C. serrulata var. serrulata f. spontanea (Maxim.) C. S. Chang、P. serrulata var. pubescens (Makino) Nakai。王贤荣等[19]对野生早樱的不同种群形态变异进行深入探讨,由此支持了宽叶早樱(P. subhirtella var. platyphyllus X. R. Wang, C. P. Xie et X. G. Yi)与小花早樱(C. subhirtella var. miniflora X. R. Wang, C. P. Xie et X. G. Yi)新变种的成立。南程慧[20]详细分析了迎春樱桃不同种群的形态变异情况,并为垂枝迎春樱(P. discoidea var. pendula C. H. Nan, X. R.Wang et G. G. Tang)与大花迎春樱(P. discoidea var. grandiflora C. H. Nan, X. R. Wang et G. G. Tang)两个新变种成立提供了有力的证据。伊贤贵[21]对山樱花复合体的进行了主要形态性状的聚类分析支持P. serrulata var. huanggangensis X.R. Wang et X.G. Yi变种黄岗山樱(P. serrulatita var. huanggangensis X. R. Wang et X. G. Yi)的成立。朱弘[22]对尾叶樱桃种群形态变异的分析,也为长柱尾叶樱桃(P. dielsiana var. longistyla X. R. Wang, X. G. Yi et C. P. Xie)的成立提供了有力的支撑。微形态学标记(morphological markers)是传统形态学标记的重要补充。王贤荣等[23]通过光学显微镜和扫描电镜,对樱亚属植物进行了叶表皮微形态观察,表明樱亚属植物叶表皮的毛被、气孔及外拱盖等特征,在种内保持稳定,而在种间差异显著。叶表皮微形态为樱亚属物种间的分类处理提供了新的视角和证据。花粉形态作为微形态学研究重要内容,同样受到了学者们的关注,王伏雄[24]与蓝盛银等[25]相继对樱桃(P. pseudocerasus)、山樱花(P. serrulta)、东京樱花(P. yedoensis)的花粉形态进行了研究。王贤荣等[26]对樱属11个种和变种的花粉形态进行了细致的扫描电镜观察。周丽华等[27]观察了高盆樱桃(P. cerasoides)的花粉形态。雷海清等[28]对樱属9个种和变种的花粉形态进行扫描电镜观察,伊贤贵等[21]和南程慧等[20]对武夷山、天目山等地区樱亚属植物花粉进行了电镜观察。上述报道均表明樱亚属花粉在极面观上多呈圆形或近三裂圆形,赤道面观则为椭圆形、三孔沟,外壁纹饰多为条纹状。尽管属内特征基本一致,但种间存在着花粉大小和外壁纹饰的差别,花粉微形态为界定樱亚属植物不同类群提供了重要的微形态数据。
樱亚属植物分类研究建立在对其叶、花、果等核心性状的细致观测之中。腺体位置和形状在樱亚属植物种间具有显著差异,叶表皮的微形态特征也有变异,如毛被的有无、密度和长度,以及表皮毛的形态、气孔的外拱盖和内缘纹饰、气孔的形状与大小,以及表皮蜡层的微妙纹饰。这些微形态特征的差异,为樱亚属植物的分类界定提供了重要辅助证据[19-21]。花粉微形态在樱亚属中整体上较为一致,但花粉形态、花粉大小、外壁纹饰等在种间具有一定的变化,可为樱亚属部分种或种下等级分类处理提供证据[26-28]。借助形态与微形态标记手段,分类学者成功报道了多个樱亚属类群,包括鹤峰樱桃(P. hefengensis X. R. Wang amp; C. B. Shang)[29]、武夷红樱(P. campanulata var. wuyiensis X. R. Wang, X. G. Yi amp; C. P. Xie)[30]、雪落樱桃(P. xueluoensis C. H. Nan amp; X. R. Wang)[31]、崂山樱桃(P. laoshanensis D. K. Zang)[32]、沼生矮樱(P. jingningensis Z. H. Chen, G. Y. Li amp; Y. K. Xu)[33]、景宁晚樱(P. paludosa R.L. Liu, W.J. Chen et Z.H. Chen)[34]和熊野樱桃(P. kumanoensis T. Katsuki)[35],其他基于形态与分子标记证据报道的新类群见后文。上述基于形态标记的新类群的不断报道,表明樱亚属物种及其变异丰富,具有很高的物种多样性潜力。因此,形态学标记依然是樱亚属分类研究的前提,即使在分子标记手段十分丰富的分类学新阶段,新类群的发现与报道依然需要形态学数据提供重要支撑;未来,在重视分子标记手段应用的同时,同样应该重视樱亚属植物的野外观测、标本采集以及形态标记等在分类学中的基础作用。
2.2 细胞及化学分类学研究进展
早期樱亚属分类研究往往更多地集中在形态学标记及其应用价值上,较少开展细胞及生物化学方面的研究。然而,这些微观层面的研究对于早期理解樱亚属植物的起源、进化及分类具有重要意义。历史上,Kobel等学者对李属植物的染色体数目进行了详细的研究,最早阐述了P. avium (2n=16)可能为李属的原始种类之一,而P. cerasus (2n=4x=32)则可能是通过种间杂交形成的[5];这也证明了樱亚属植物天然杂交与基因渐渗情况的发生。日本学者在樱属植物(包括园艺品种)和广义李属的部分种类上进行了染色体数目统计和核型研究[36],他们的研究结果表明,樱属植物大部分为二倍体(2n=16),部分园艺品种则表现出多倍体的特征。其中,晚樱(P. serrulata var. lannesiana)种系下的三倍体品种花期明显晚于其原变种,这一结果为理解多倍体在樱亚属植物进化中的作用提供了证据。此外,Mowrey等[37]学者利用同工酶标记技术对34个李属植物(包括Subgen. Cerasus的12种)进行了系统发育研究,他们的研究结果表明樱亚属内的植物在系统发育上能够聚合为一类。同时,他们的研究还推测P. cerasus可能是P. avium与P. fruticosa(2n=4x=32)的杂交种,P. gonduinii可能是P. ceraus与P. avium的杂交种。Granger等[38]等通过对甜樱桃叶内同工酶的分析,探讨了甜樱桃栽培种的分类和遗传多样性方面的问题。这些发现为早期推测樱亚属植物的进化历史提供了理论依据。周春玲等[39]利用酯酶同工酶技术对青岛市的樱花品种进行了亲缘关系鉴定和品种分类研究,他们的研究结果与传统分类学结果一致,也验证了生物化学标记在植物分类学中的有效性。顾宇等[40]则统计了18份樱亚属植物的核型数据,他们的研究结果表明野生类群多为二倍体,而栽培品种则表现出显著的多倍化现象,支持黑果组、芽鳞组的成立。李祯等[41]测定了山樱花基因组的大小,认为其可用于樱亚属植物倍性鉴定。综上,樱亚属的细胞及生物化学分类研究开展较早,同时涉及的类群数量较少,相关证据仅能对少数种系的分类处理提供依据,整体上对樱亚属系统分类贡献较小;随着分子标记方法的持续优化,细胞及生物化学手段在樱亚属分类研究中价值并不高,未来的樱亚属系统分类研究将更多的基于分子标记证据。
2.3 分子标记分类研究进展
在樱亚属分类学领域,分子标记技术已成为揭示遗传多样性的重要工具。它广泛涵盖DNA、RNA和蛋白质标记,但实际应用中常以DNA标记为主。检测遗传多样性的分子方法主要包括两大途径:一是通过识别基因组的特定位点来评估基因多态性;二是直接对特定基因(组)或DNA片段进行测序,检测核苷酸变异,如叶绿体(cpDNA)、线粒体(nrDNA)、核基因(nrDNA)及全基因组(WGS)序列等[5,22]。这些技术的应用,极大推动了樱亚属分类学研究的深入。
2.3.1 位点标记樱亚属分类研究进展
要估量基因组的多态性,通常通过酶切和杂交技术(如RFLPs)以及基于聚合酶链式反应(PCR)的标记方法(如RAPD、AFLP、ISSR、SSR等)。在早期对樱亚属的分子遗传研究中,RFLP、RAPD、AFLP等技术被广泛采用[22]。尽管这些方法能在一定程度上揭示了樱亚属植物类群间的亲缘关系,但总体而言,它们对樱亚属系统分类学的贡献较为有限。
在樱亚属植物的研究领域中,SSR标记(微卫星DNA)因具有分辨率较高且成本较低的优势,受到了广泛的关注和应用。Dirlewanger等[42]、Aranzana等[43]、Canli等[44]、Kacar等[45]从樱属及其相近属种中成功获取引物,并进行扩增实验。这些研究充分展示了SSR引物在广义李属中的良好通用性和丰富的多态性,为遗传多样性分析、指纹图谱绘制、物种鉴定及亲缘关系研究等提供了有力支撑。沈志军[46]的研究深入探讨了樱桃与桃、梅、杏、李在系统发育上的关系,为理解这些植物间的亲缘关系提供了新的视角。李苗苗[47]则对樱亚属10个种的47个种群进行分子亲缘地理学研究,进一步揭示了樱亚属植物的遗传结构和分布特点。南程慧[20]利用SSR标记技术,对迎春樱桃(P. discoidea)的遗传多样性进行了深入的探索。张琼[48]在广义李属的SSR分析中,揭示了典型樱亚属与桃亚属、李亚属之间的亲缘关系。朱淑霞[49]利用SSR技术结合形态学特征对107个樱亚属种及品种进行分类讨论,其分子标记结果与形态标记结果高度一致,进一步验证SSR技术在樱亚属植物分类研究中的可靠性。孙泽硕等[50]则通过SSR指纹图谱的构建,为42个不同种系樱花品种的鉴定和分类提供了有力的技术支持。综上所述,SSR标记以其成熟的引物开发技术、高度的通用性以及良好的标记效果,在樱亚属植物的群体遗传学研究和种内微进化分析中依然发挥着重要作用,可以为樱亚属植物种下及亲缘关系较近的种间关系讨论与分类处理提供有力证据。
2.3.2 序列标记樱亚属分类研究进展
近年来,测序技术的快速发展为樱亚属分类、物种界定与系统进化研究带来了前所未有的机遇。特别是DNA序列标记,在樱亚属分类界定与系统发育研究中取得了显著的成果。利用nrITS标记,Lee等[51]构建了李属的系统发育树,系统建树揭示了典型樱亚属Subgen. Cerasus与稠李亚属Subgen. Padus及桂樱亚属Subgen. laurocerasus之间的紧密亲缘关系,它们共同发育形成了姐妹群。刘艳玲等[52]也基于nrITS序列证实了樱亚属作为单系分支的存在,并揭示了其位于李属系统发育树的基部位置。Shaw等[53]则基于叶绿体非编码区片段rpL16和trnG,构建了系统发育树,结果显示樱亚属与李亚属和桃亚属同样形成了姐妹群关系。Bortiri等[54]结合nrITS,与叶绿体trnL-trnF构建了更为完善的系统发育树,结果表明樱亚属、稠李亚属、桂樱亚属在李属内构成了一大枝系;而矮生樱亚属并非单系,其下多数种类与李亚属和桃亚属更为接近。为了进一步探讨李属内部的系统发育关系,Bortiri等[55]增加了单拷贝核基因s6pdh片段和叶绿体间隔区trnS—trnG的片段,并结合形态学特征进行了深入分析。这项研究显示李属早期分化为两大支,一支由樱亚属、桂樱稠亚属、李亚属组成,另一支则由桃亚属、扁桃亚属(Subgen. Emplectocladus)、李亚属组成;而矮李组与李亚属的关系则相对复杂,它们之间存在相互嵌套的现象,且支持度较低,这一结果为后续的系统发育研究提供了良好的基础。石硕[56-57]基于前人的研究基础,应用新开发的核基因片段SbeI-a及3个cpDNA片段atpH-atpI、petA-psbJ和petN-psbM,通过最大简约法(MP)、最大似然法(ML)和贝叶斯法(Bayes)3种算法完整地重建了李属与樱亚属的系统发育树,该研究结果构建了樱亚属的系统基本框架,并支持矮生樱亚属(矮李组)不属于樱亚属的观点;该研究未能获得高支持率的樱亚属系统发育树。DNA序列不仅为系统发育研究提供了基础数据,也为新种的发现提供了有力证据。例如,近年来报道的磐安樱桃(P. pananensis Z. L. Chen, W. J. Chen amp; X. F. Jin)、凤阳山樱桃(P. fengyangshanica L.X.Ye amp; X.F.Jin)、孙航樱桃(P. sunhangii D. G. Zhang amp; T. Deng)、全州樱桃(P. quanzhouensis Jian L.Li, S.H.Liang amp; R.H.Jiang)以及桐木樱桃(P. tongmuensis X.G.Yi amp; X.R.Wang)等,这些新种的发现与报道丰富了樱亚属的物种多样性,也说明樱亚属资源分类还有许多未完工作[58-62]。
随着测序技术的不断进步和应用范围的扩大,利用全基因组数据进行研究的报道也日益增多。基于叶绿体全基因组数据建树,基本理清了樱亚属的系统发育关系[63-66]。同时,欧洲甜樱桃、东京樱花、染井吉野(P.× yedoensis ‘Somei-Yoshino’)以及山樱花和钟花樱桃等的全基因组测序结果相继完成并公布[67-72],这为开发具有广泛可应用性的遗传标记如SNP、SRAP和低拷贝核基因提供了便利。未来,借助简化基因组、全基因组测序和重测序等方法,为樱亚属系统分类及更小的分类单元的研究提供更加丰富的信息。
3 樱亚属种类与分布
樱亚属分类系统不断调整与完善,矮生樱亚属类群从樱亚属中移出,同时部分樱亚属植物进行独立或合并处理,加之新物种的不断发表,导致樱亚属物种名录存在较大的争议。根据文献资料以及标本查阅整理,并参考“Plants of the World Online”(POWO)数据,目前学名被广泛接受的樱亚属物种约为60种,但部分类群分类地位及处理仍存在争议。结合文献、标本资料以及野外调查情况[5,73-75],可以推测世界范围内樱亚属植物的种类约为80种(含种下等级)。在处理了部分争议类群后,笔者较为全面地列出了世界樱亚属物种名录及其主要地理分布,共计76种(包括11个变种),具体情况如表2所示。经过整理后,可以更清晰地了解各类樱亚属植物的数量及分布范围,有助于今后的研究和保护工作。
樱属植物主要分布于北半球的温带地区,主要集中在东亚地区,包括中国、日本、俄罗斯和朝鲜半岛。中国樱亚属植物种类最为丰富,共计54种(含10变种),其中42种(含9变种)为特有分布。喜马拉雅山区至中国西南地区分布有27种樱亚属植物(含3变种),中国中东部(含东南部)分布有22种樱亚属植物(含6变种)。此外,北美分布有2种,欧洲、地中海至中亚地区分布有3种。总体来看,中国西部、西南部以及日本和朝鲜一线(即喜马拉雅至日本)是世界上樱亚属植物种类最为集中的地区。中国的樱亚属植物种类丰富,分布广泛,是樱属植物的重要分布地之一。与中国相比,日本共有11种樱亚属植物(含1变种),分布相对较少。因此,可以说中国是世界上樱亚属植物种类最为丰富的国家。樱亚属植物的分布情况如表2所示[74-75],樱亚属类群在北半球温暖地区呈现出丰富的物种多样性以及区域性分布的特点。
4 樱亚属系统发育、起源与演化
石硕[57]利用12个叶绿体DNA和3个核基因片段的系统发育分析,构建了广义李属全部10 类77 个代表物种的系统发育关系。将广义李属细分为3个亚属:稠李亚属(Subgen. Padus)、樱亚属(Subgen. Cerasus)和李亚属(Subgen. Prunus)。在对樱亚属的研究中,他确认了矮李(矮生樱亚属)并不属于樱亚属,而将矮生樱亚属的欧李(P. humilis)、郁李(P. japonica)、麦李(P. glandulosa)、天山樱桃(P. tianshanica)的4个种转入李亚属,同时认定了P. mahaleb 和 P. maackii属于樱亚属。然而,樱亚属的系统树支持率不高,这与先前基于形态的分类系统并不一致[56-57]。吴宝欢[78]则采用简化基因组测序技术进行了更广泛的研究,通过使用82 个李属樱亚属的分子样品展开了系统发育分析,发现圆叶樱桃和斑叶稠李并不属于樱亚属。最近的研究表明,基于叶绿体全基因组数据建树可以更好地阐明樱亚属的系统发育关系[63-66]。综合分析结果可以得出一些主要结论:樱亚属是1个单系群,矮生樱类实际上属于李亚属;斑叶稠李并非樱亚属成员;根据形态和地理分布可将樱亚属划分为约10个支系或组;圆叶樱桃是樱亚属的基部类群,而喜马拉雅山脉至中国西南地区的一些类群则构成了樱亚属的重要支系;另一些类群则在中国东部至朝鲜半岛和日本分布,包括山樱花复合群、迎春樱桃、尾叶樱桃、钟花樱桃、太平山樱、山白樱、阿里山樱、熊野樱桃、高岭樱桃、丁字樱桃和豆樱等,它们是樱亚属中最晚分化的支系之一。这些研究成果丰富了对樱亚属植物的分类和系统发育的认识,为进一步研究提供了重要的参考依据。通过综合利用叶绿体和核基因片段的方法,可以更准确地划分类群的关系,对于了解植物进化和物种起源具有重要意义。希望未来的研究能够继续深入,进一步完善对李属樱亚属植物的分类和生物学特征的理解,为保护和利用这一类植物资源提供科学依据。
根据序列片段、叶绿体全基因组以及全基因组建树和分子钟定年结果,可以推测樱亚属植物起源于约5 497万年或3 927万年前的始新世时期;在约1 501万年或1 157万年前,李属开始独立分化出樱亚属,最基部类群为圆叶樱桃[56,63-66]。中国西南地区(横断山—喜马拉雅山)是樱亚属的主要分布区域,拥有较多的特有种,符合樱亚属起源地的条件。根据樱亚属植物现代分布格局以及系统学研究,喜马拉雅地区至中国西南高山地区或地中海地区被认为是本亚属起源地。研究表明,水热条件与生境异质性是影响樱亚属植物分布的主要驱动因素[75]。中国西南部、东南部以及中国东北—朝鲜半岛—日本—俄罗斯远东地区是樱亚属的当代分布中心。中国中东部曾是樱亚属植物的避难所和扩散中心。樱亚属历史演化的主要迁移扩散路线包括:从喜马拉雅山经中亚和西亚扩散;经中国西南高山地区至中国中部和东部地区,然后经东北到达朝鲜半岛和日本,最终通过白令海峡到达北美;部分中国中部和东部类群则从中国东部经台湾岛到达日本和朝鲜半岛。综上所述,樱亚属植物的起源可以追溯至始新世时期,其分布格局受到水热条件和生境异质性的影响。中国西南地区是该亚属主要种类的集中分布区,而樱亚属的起源地可推测为喜马拉雅地区至中国西南高山地区或地中海地区。樱亚属演化的扩散路线经历了多个地区,形成了现代的分布格局。这些研究成果为更深入了解樱亚属植物的起源与演化提供了重要的参考。
5 讨论与展望
5.1 争议类群
1)山樱花复合体(P. serrulata complex)是一个充满着变化和多样性的群体,其分类工作一直备受关注与困扰。自1830年Lindley[76]首次描述山樱花为花复瓣的P. serrulata开始,对该群体的研究就引起了学者们的兴趣。1916年Wilson[6]认为描述山樱花的标本为栽培种,发现花单瓣的野生类群,随后将其归为山樱花变种P. serrulata var. spontanea,并记录了另一种有毛的山樱花(P. serrulata var. pubescens)。1911年,Koidzumi[77 ]记录了红山樱]P. jamasakura (Makino) Siebold ex Koidz.];1912年,Koehne[4]记录了霞樱 (P. leveilleana),其有毛被与毛叶山樱花较相似。在复杂的山樱花复合体中,晚樱[P. serrulata var. lannesiana(P. lannesiana)]被记录主要为栽培品种,其亲本包括大岛樱(P. speciosa)、红山樱(P. jamasakura)和大山樱(P. sargentii)等,种间和种内的杂交现象使得各个类群之间的关系变得模糊不清。这种分类困难也源于山樱花在形态特征上的多样性,不同的花、叶和果实在不同的生长期呈现出明显的变化。通过对文献和标本的查阅以及野外调查[32,78-80],结合部分序列数据,笔者支持将毛叶山樱花、崂山樱(P. laoshanensis)和泰山野樱花(P. serrulata var. taishanensis)归入山樱花的范畴。同时,也支持建立雪落樱桃这一类群,并赞成新变种黄岗山樱(P. serrulata var. huanggangensis)的确立。对于红山樱、大山樱和大岛樱,分别提出了独立为种的分类处理观点。山樱花分布广泛,遍布于中国、韩国和日本等地,不同地区间存在着连续且交叉的形态特征变异。因此,对于山樱花复合体各类群之间关系的研究仍需要进一步的探讨。对于这一复杂群体的准确描述和记录,是未来研究的一个重要课题。
2)大叶早樱复合群(P. subhirtella complex),是一个具有壶形萼筒显著特征的樱花群体。自19世纪以来,该复合群的命名和分类一直备受争议。经过对历史文献、标本和野外调查的综合研究,认为其分类和命名有待进一步的澄清。最早对于大叶早樱复合群进行描述的是Siebold(1836)和Miqul(1865),他们分别记录了P. itosakura和日本常见的1种栽培樱花P. subhirtella Miq. Lavallée ex Ed.,Otto(1879)记录了江户彼岸(P. spachiana),Makino(1893)记录了垂枝大叶早樱(P. pendula var. ascendens),Hayata(1911)记录了雾社樱(P. taiwaniana),Wilson[6](1916)则将大叶早樱野生种作为变种记录为野生早樱(P. subhirtella var. ascendens),Ingram(1949)描述了中国湖北长阳县产的长阳樱桃(P. changyangensis)[5]。然而,在日本的《Flora of Japan》中,P. subhirtella 被限定为仅指栽培种,野生早樱被视为江户彼岸樱的异名。对于江户彼岸樱的学名,Katsuki等[81]认为P. itosakura (Siebold) Masam.amp; Suzuki更为准确。此外,Zhang等[60]学者还发表了1个新种——孙航樱桃(P. sunhangii)。通过对文献、标本和野外调查的详细研究,将杂交起源的长阳樱桃纳入大叶早樱分类记录中,并称之为P. × subhirtella;将野生类群江户彼岸(伊藤樱桃)记录为P. itosakura;将孙航樱桃归入野生早樱记录并称之为P. itosakura var. ascendens;雾社樱桃因其花色白、花期长,处理为变种记录为P. itosakura var. taiwaniana。这一复合群分布在中国、日本、韩国等国家,由于各国学者对其分类的观点不一致以及整体采样不足,因此亲缘地理和物种的确立仍需要进一步的研究。综上,大叶早樱复合群(P. subhirtella complex)的分类学问题仍然存在许多争议,需要更加深入的研究和全面的采样来确定确切的分类和命名。通过对历史文献和标及分子系统学研究的综合分析,可以更好地理解这一群体的起源和演化,为其未来的分类学研究提供重要的参考。
3)其他类群的处理。经过文献研究、标本查阅和野外调查,笔者对于尾叶樱桃、长柱尾叶樱桃等类群的成立提出了支持意见。尾叶樱桃和长柱尾叶樱桃的确存在一定程度的差异,而短梗尾叶樱桃则需要更多的数据来支持其独立性。此外,认为磐安樱桃可以并入尾叶樱桃,而浙闽樱桃可能是尾叶樱桃的1个变异群体,对于其是否成立及其系统地位仍需进一步研究。至于全州樱桃,认为宜将其处理为华中樱桃的1个变种P. conradinae var. quanzhouensis 。对于其他争议类群的处理,笔者也认为需要更多的数据支持才能得出科学结论。
5.2 展望
随着分子技术的不断进步和系统发育研究的深入,樱亚属的系统分类和物种范围已经基本明晰。目前,樱亚属系统发育研究涉及最多的为55个种类,研究采用叶绿体全基因组数据构建系统发育树[63-66]。樱亚属植物以其极高的观赏价值而闻名,一直是植物学研究的热门课题,不断有新的物种被报道。因此,未来的研究需要更广泛地收集物种样本,采用全基因组和重测序等大数据的方法,结合形态学等统计性状,不断完善樱亚属的系统发育树和分类系统。这对于更好地了解和保护这一类群具有重要意义。
传统分类系统与分子分类系统在樱亚属研究中存在着分歧,而受样本采集准确性与完整性影响,关于樱亚属部分类群的争议将长期存在。例如山樱花复合群与大叶早樱复合群由于分布广泛且不同国家主流分类观点不一致,仍然是樱亚属分类研究的争议焦点[74,78]。对于雪落樱桃、沼生矮樱以及维氏樱桃(P. veitchii)等种类之间的关系处理,尚需更多的分子数据支持。细花樱桃与蒙自樱桃,浙闽樱桃与尾叶樱桃,桐木樱桃与景宁晚樱,维西樱桃、姚氏樱桃与毛瓣藏樱等近似种的分类处理同样需要更多的样本数据。近年来,中国东南部地区报道了较多的樱亚属新类群,而喜马拉雅山区至中国西南地区以及华中地区的野外调查难度大,还需要更多的野外调查与观测,不排除未来有更多的新类群被报道。樱亚属植物为落叶树种,花、果期较短,使得野外调查与样本采集更加困难,对于存疑类群的研究需要更长的时间周期。总之,樱亚属植物具有极高的观赏与经济价值,因此其系统分类学研究具有重要意义,未来需要有更多的研究人员开发多区域的樱亚属植物资源调查,进行多物种、多群体、多组学及多分布区相结合的系统研究,以解析其系统发育及进化演化的科学问题,并为樱亚属植物的保护与利用打下基础。
参考文献(reference):
[1]LINNAEUS C. Species plantarum. Vol.I[M]. Holmiæ (Stockholm): Impensis Laurentii Salvii, 1753: 473.
[2]MILLER P. Gardeners dictionary[M].4th ed. London, 1754: 307-314.
[3]GRAY A.Manual of the botany of the northern United States[M].2nd ed. New York: George P Putnam amp; Co, 1856.DOI: 10.5962/bhl.title.50405.
[4]KOEHNE E. Plantae wilsonianae. Part 1[M]. Cambridge: The University Press, 1912: 59, 226-273.
[5]伊贤贵.山樱花种群变异及亲缘地理学研究[D].南京:南京林业大学,2018.DOI: 10.27242/d.cnki.gnjlu.2018.000005.YI X G.The variation and phylogeography of Cerasus serrulata Mill.populations[D].Nanjing:Nanjing Forestry University,2018.
[6]WILSON E H. The Cherries of Japan. No.7[M]. Massachusetts: Arnold Arboretum Publication, 1916.
[7]陈嵘.中国树木分类学[M]. 北京: 科学出版社, 1937: 473-481. CHEN R. Tree Taxonomy in China[M]. Beijing: Science Press, 1937: 473-481.
[8]REHDER A. Manual of cultivated trees amp; shrubs[M]. 2nd ed. Massachusetts: Aronold Abroretum Publication, 1951: 466-477.
[9]SHISHKIN B K, YUZEPCHUK S V. Flora of U.S.S.R. Volume XI[M]. Jerusalem: Israel Program for Scientific Translations, 1971.
[10]郑万钧.中国树木志:第二卷[M].北京:中国林业出版社,1985.
[11]俞德浚,李朝銮.中国植物志:38卷[M].北京:科学出版社, 1986: 41-87. YU D J, LI C L. Flora reipublicae popularis sincae. Vol. 38[M]. Beijing: Science Press, 1986: 41-87.
[12]川崎哲也. 日本の桜[M]. 东京: 山と渓谷社, 1994. KAWASAKI T. Flowering cherries of Japan[M]. Tokyo: Tokyo: Yama-Kei Publisher Co. ltd., 1994.
[13]大场秀章, 川崎哲也, 田中秀明. 新日本の樱[M].东京:山と溪谷社, 2007. OHBA H, KAWASAKI T, TANAKA H. Flowering cherries of Japan[M]. New edition. Tokyo: Yama-Kei Publisher Co.ltd.,2007.
[14]王贤荣.国产樱属分类学研究[D].南京:南京林业大学,1997.WANG X R.Taxonomic study on Sakura from China[D].Nanjing:Nanjing Forestry University,1997.
[15]王贤荣.中国樱花品种图志[M].北京:科学出版社,2014:19-23. WANG X R.An illustrated monograph of cherry cultivars in China[M].Beijing:Science Press,2014:19-23.
[16]IWATSUKI K, BOUFFORD D E, OHBA H. Flora of Japan: Vol. IIb[M]. Tokyo: Kodansha, 2001.
[17]LI C L, BRUCE B. Flora of China: 9[M]. Beijing: Science Press, Missouri:Missouri Botanical Garden Press, 2003: 404-420.
[18]CHANG K S,CHANG C S,PARK T Y,et al.Reconsideration of the Prunus serrulata complex (Rosaceae) and related taxa in eastern Asia[J].Bot J Linn Soc,2007,154(1):35-54.DOI: 10.1111/j.1095-8339.2007.00631.x.
[19]王贤荣,谢春平,伊贤贵,等.不同居群野生早樱形态变异研究[J].植物研究,2007,27(6):746-752.WANG X R,XIE C P,YI X G,et al.Study on the morphological variations of Cerasus subhirtella var.ascendens in different populations[J].Bull Bot Res,2007,27(6):746-752.DOI: 10.3969/j.issn.1673-5102.2007.06.018.
[20]南程慧.迎春樱居群变异与繁殖生物学研究[D].南京:南京林业大学,2012.NAN C H.Study on population variation and reproductive biology of Cerasus discoidea Yü et Li[D].Nanjing:Nanjing Forestry University,2012.
[21]伊贤贵.武夷山樱属资源调查及开发利用研究[D].南京:南京林业大学,2007.YI X G.Studies on investigation and exploitation of the resources of Cerasus Mill.in Wuyi Mountain[D].Nanjing:Nanjing Forestry University,2007.
[22]朱弘.尾叶樱桃(Cerasus dielsiana)系统分类地位与种群生物地理学研究[D].南京:南京林业大学,2020.ZHU H.Phylogenetic position and population biogeography of Cerasus dielsiana(Rosaceae)[D].Nanjing:Nanjing Forestry University,2020.DOI: 10.27242/d.cnki.gnjlu.2020.000023.
[23]王贤荣,向其柏.樱属植物叶腺体形态研究及其在分类中的意义[J].南京林业大学学报,1997,21(4):63-67. WANG X R,XIANG Q B.Morphological study on leaf glands of Sakura and its significance in classification[J].J Nanjing For Univ,1997,21(4):63-67. DOI: 10.3969/j.jssn.1000-2006.1997.04.013.
[24]王伏雄.中国植物花粉形态 [M].2版.北京:科学出版社, 1995. WANG F X. Pollen flora of China. [M]. 2nd ed. Beijing: Science Press, 1995.
[25]蓝盛银,徐珍秀.植物花粉剥离观察扫描电镜图解[M].北京:科学出版社, 1996. LAN S Y, XU Z X. Observation of pollen morphology of plants using scanning electron microscopy[M]. Beijing: Science Press, 1996.DOI: 10.3969/j.jssn.1000-2006.1998.04.012.
[26]王贤荣,向其柏.樱属植物分类订正[J].南京林业大学学报,1998,22(4):60-62.WANG X R,XIANG Q B.Notes on some species of the genus Cerasus Miq[J].J Nanjing For Univ,1998,22(4):60-62. DOI:10.3969/j.jssn.1000-2006.1998.04.012.
[27]周丽华,韦仲新,吴征镒.国产蔷薇科苹果亚科的花粉形态[J].云南植物研究,2000,22(1):47-52,121-123.ZHOU L H,WEI Z X,WU Z Y.Pollen morphology of maloideae of China (Rosaceae)[J].Acta Bot Yunnanica,2000,22(1):47-52,121-123.DOI: 10.3969/j.issn.2095-0845.1999.04.008.
[28]雷海清.樱属花粉形态研究[J].亚热带植物科学,2001,30(4):14-17.LEI H Q.A study on pollen morphology of Cerasus[J].Subtrop Plant Sci,2001,30(4):14-17.DOI: 10.3969/j.issn.1009-7791.2001.04.004.
[29]WANG X R, SHANG C B. Cerasus hefengensis (Rosaceae), a new species from SW Hubei, China[J]. Annales Botanici Fennici, 2007, 44: 151-152.
[30]WANG X R,YI X G,XIE C P.Cerasus campanulata var. wuyiensis,a new variety of Rosaceae in Wuyi Mountain[J].Acta Bot Yunnanica,2007(6):616.
[31]NAN C H,WANG X R,TANG G G,et al.Cerasus Xueluoensis(Rosaceae),a new species from China[J].Ann Bot Fenn,2013,50(1/2):79-82.DOI: 10.5735/085.050.0114.
[32]ZANG D K.Cerasus laoshanensis (Rosaceae),a new species from Shandong,China[J].Ann Bot Fenn,2017,54(1/2/3):135-137.DOI: 10.5735/085.054.0321.
[33]许元科,赵昌高,严邦祥,等.浙江樱属新种:沼生矮樱[J].浙江林业科技,2012,32(4):81-83.XU Y K,ZHAO C G,YAN B X,et al.A new species of Cerasus Mill.from Zhejiang Province[J].J Zhejiang For Sci Technol,2012,32(4):81-83.DOI: 10.3969/j.issn.1001-3776.2012.04.020.
[34]刘日林,张方钢,陈伟杰,等.浙江蔷薇科李亚科植物新资料[J].杭州师范大学学报(自然科学版),2017,16(5):518-521.LIU R L,ZHANG F G,CHEN W J,et al.Noteworthy plants in prunoideae of Rosaceae from Zhejiang[J].J Hangzhou Norm Univ (Nat Sci Ed),2017,16(5):518-521.DOI: 10.3969/j.issn.1674-232X.2017.05.012.
[35]KATSUKI T. A new species, Cerasus kumanoensis from the southern Kii Peninsula, Japan[J]. Acta Phytotax Geobot,2018,69(2):119-133. DOI:10.18942/APG.201801.
[36]OGINUMA K. Karyomorphological studies on Prunus in Japan [J]. Journal of Science of the Hiroshima University, Series B, Division 2 (botany), 1987, 21: 1-66.
[37]MOWREY B D,WERNER D J.Phylogenetic relationships among species of Prunus as inferred by isozyme markers[J].Theor Appl Genet,1990,80(1):129-133.DOI: 10.1007/BF00224026.
[38]GRANGER A R,CLARKE G R,JACKSON J F.Sweet cherry cultivar identification by leaf isozyme polymorphism[J].Theor Appl Genet,1993,86(4):458-464.DOI: 10.1007/BF00838561.
[39]周春玲,陈芳,苗积广,等.青岛市19个樱花品种的酯酶同工酶鉴定[J].西北林学院学报,2008,23(3):40-43.ZHOU C L,CHEN F,MIAO J G,et al.Identification of 19 Cerasus cultivars on esterase isozyme in Qingdao[J].J Northwest For Univ,2008,23(3):40-43.
[40]顾宇,史港影,李祯,等.樱属植物核型参数及种间亲缘关系分析[J].南京林业大学学报(自然科学版),2014,38(S1):25-29.GU Y,SHI G Y,LI Z,et al.Karyotype parameters and interspecific relationship analysis of Sakura[J].J Nanjing For Univ (Nat Sci Ed),2014,38(S1):25-29.
[41]李祯,伊贤贵,顾宇,等.山樱花基因组大小的测定[J].南京林业大学学报(自然科学版),2014,38(S1):17-19.LI Z,YI X G,GU Y,et al.Determination of genome size of Prunus mume[J].J Nanjing For Univ (Nat Sci Ed),2014,38(S1):17-19.DOI: 10.3969/j.issn.1000-2006.2014.S1.004.
[42]DIRLEWANGER E,COSSON P,TAVAUD M,et al.Development of microsatellite markers in peach[Prunus persica (L.) Batsch]and their use in genetic diversity analysis in peach and sweet cherry (Prunus avium L.)[J].Theor Appl Genet,2002,105(1):127-138.DOI: 10.1007/s00122-002-0867-7.
[43]ARANZANA M,CARBÚ J,ARÓS P.Microsatellite variability in peach[Prunus persica (L.) Batsch]:cultivar identification,marker mutation,pedigree inferences and population structure[J].Theor Appl Genet,2003,106(8):1341-1352.DOI: 10.1007/s00122-002-1128-5.
[44]CANLI F A.Development of a second generation genetic linkage map for sour cherry using SSR markers[J].Pak J Biol Sci,2004,7(10):1676-1683.DOI: 10.3923/pjbs.2004.1676.1683.
[45]KACAR Y L Z A,IEZZONI A,CETINER S.Sweet cherry cultivar identification by using SSR markers[J].J Biol Sci,2005,5(5):616-619.DOI: 10.3923/jbs.2005.616.619.
[46]沈志军.基于微卫星标记的核果类果树亲缘关系研究[D].南京:南京农业大学,2003.SHEN Z J.Genetic relationship of peach,plum,apricot,mume and cherry based on microsatellite markers[D].Nanjing:Nanjing Agricultural University,2003.
[47]李苗苗.樱亚属植物分子亲缘地理及中国樱桃自然居群遗传多样性研究[D].西安:西北大学,2009.LI M M.Phylogeography of Prunus subgenus Cerasus and genetic diversity of P. pseudocerasus[D].Xi’an:Northwest University,2009.
[48]张琼.樱属观赏品种资源调查及部分种与品种SSR分析[D].南京:南京林业大学,2013.ZHANG Q.Study on the cultivar resources investigation of flowering cherry (Cerasus) in China and analysis of particle specises and cultivars using SSR markers[D].Nanjing:Nanjing Forestry University,2013.
[49]朱淑霞.基于形态学和SSR分子标记的樱属品种指纹图谱构建[D].南京:南京林业大学, 2020. ZHU S X. Fingerprinting of Cerasus cultivars based on morphology and SSR molecular markers[D]. Nanjing: Nanjing Forestry University, 2020.
[50]孙泽硕,蒋冬月,柳新红,等.基于SSR标记的42份樱花品种的聚类分析及DNA指纹图谱构建[J].园艺学报,2023,50(3):657-668.SUN Z S,JIANG D Y,LIU X H,et al.Cluster analysis and construction of DNA fingerprinting of 42 oriental cultivars of flowering cherry based on SSR markers[J].Acta Hortic Sin,2023,50(3):657-668.DOI: 10.16420/j.issn.0513-353x.2021-1057.
[51]LEE S,WEN J.A phylogenetic analysis of Prunus and the Amygdaloideae (Rosaceae) using ITS sequences of nuclear ribosomal DNA[J].Am J Bot,2001,88(1):150-160.
[52]刘艳玲,徐立铭,程中平.基于ITS序列探讨核果类果树桃、李、杏、梅、樱的系统发育关系[J].园艺学报,2007,34(1):23-28.LIU Y L,XU L M,CHENG Z P.Phylogenetic analysis of stone fruits such as peach,plum,apricot,mume and cherry based on ITS sequences[J].Acta Hortic Sin,2007,34(1):23-28.DOI: 10.16420/j.issn.0513-353x.2007.01.005.
[53]SHAW J, SMALL, R. Addressing the “Hardest puzzle in American pomology” phylogeny of Prunus sect. Prunocerasus (Rosaceae) based on seven noncoding chloroplast DNA regions[J]. AM J BOT, 2004, 91(6): 985-996.
[54]BORTIRI E, OH S H, JIANG J G, et al. Phylogeny and systematics of Prunus (Rosaceae) as determined by sequence analysis of ITS and the chloroplast trnL-trnF spacer DNA[J]. Systematic Botany, 2001, 26(4): 797-807. DOI: 10.1043/0363-6445-26.4.797.
[55]BORTIRI E,OH S H,GAO F Y,et al.The phylogenetic utility of nucleotide sequences of sorbitol 6-phosphate dehydrogenase in Prunus (Rosaceae)[J].Am J Bot,2002,89(10):1697-1708.DOI: 10.3732/ajb.89.10.1697.
[56]SHI S,LI J L,SUN J H,et al.Phylogeny and classification of Prunus sensulato (Rosaceae)[J].J Integr Plant Biol,2013,55(11):1069-1079.DOI: 10.1111/jipb.12095.
[57]石硕.蔷薇科李属樱亚属的系统学研究[D].北京:中国科学院大学,2013.SHI S.Systematic study on subgenus Prunus of Rosaceae[D].Beijing:University of Chinese Academy of Sciences,2013.
[58]CHEN Z L,CHEN W J,CHEN H,et al.Prunus pananensis (Rosaceae),a new species from Pan’an of central Zhejiang,China[J].PLoS One,2013,8(1):e54030.DOI: 10.1371/journal.pone.0054030.
[59]叶立新,鲁益飞,王桦,等.凤阳山樱桃:浙江樱属(蔷薇科)一新种[J].杭州师范大学学报(自然科学版),2017,16(1):19-24.YE L X,LU Y F,WANG H,et al.Cerasus Fengyangshanica(Rosaceae),a new species from Zhejiang[J].J Hangzhou Norm Univ (Nat Sci Ed),2017,16(1):19-24.DOI: 10.3969/j.issn.1674-232X.2017.01.006.
[60]ZHANG X S,JIANG Z L,YUSUPOV Z,et al.Prunus sunhangii:a new species of Prunus from central China[J].Plant Divers,2019,41(1):19-25.DOI: 10.1016/j.pld.2019.01.003.
[61]LIANG S H,QIN B,HUANG Y H,et al.Prunus quanzhouensis:A newly discovered species of Prunus subgen.Cerasus (Rosaceae) from northeastern Guangxi,China[J].Phytotaxa,2023,622(4):252-259.DOI: 10.11646/phytotaxa.622.4.2.
[62]YI X,DONG J,CHEN J,et al.Molecular and morphological evidence support a new species of Rosaceae Prunus subg.Cerasus from Wuyishan National Park,southeast China[J].PhytoKeys,2024,237:269-279.DOI: 10.3897/phytokeys.237.115098.
[63]LI M,SONG Y F,SYLVESTER S P,et al.Comparative analysis of the complete plastid genomes in Prunus subgenus Cerasus (Rosaceae):molecular structures and phylogenetic relationships[J].PLoS One,2022,17(4):e0266535.DOI: 10.1371/journal.pone.0266535.
[64]DONG J J,YI X G,WANG X R,et al.Population variation and phylogeography of cherry blossom (Prunus conradinae) in China[J].Plants,2024,13(7):974.DOI: 10.3390/plants13070974.
[65]SHEN X,ZONG W J,LI Y G,et al.Evolution of cherries (Prunus subgenus Cerasus) based on chloroplast genomes[J].Int J Mol Sci,2023,24(21):15612.DOI: 10.3390/ijms242115612.
[66]SONG Y F,ZHANG C,IDREES M,et al.Molecular phylogenetics and biogeography reveal the origin of cherries (Prunus subg. Cerasus,Rosaceae)[J].Bot J Linn Soc,2024,204(4):304-315.DOI: 10.1093/botlinnean/boad060.
[67]SHIRASAWA K,ISUZUGAWA K,IKENAGA M,et al.The genome sequence of sweet cherry (Prunus avium) for use in genomics-assisted breeding[J].DNA Res,2017,24(5):499-508.DOI: 10.1093/dnares/dsx020.
[68]BAEK S,CHOI K,KIM G B,et al.Draft genome sequence of wild Prunus yedoensis reveals massive inter-specific hybridization between sympatric flowering cherries[J].Genome Biol,2018,19(1):127.DOI: 10.1186/s13059-018-1497-y.
[69]SHIRASAWA K,ESUMI T,HIRAKAWA H,et al.Phased genome sequence of an interspecific hybrid flowering cherry,‘Somei-Yoshino’ (Cerasus × yedoensis)[J].DNA Res,2019,26(5):379-389.DOI: 10.1093/dnares/dsz016.
[70]YI X G,YU X Q,CHEN J,et al.The genome of Chinese flowering cherry (Cerasus serrulata) provides new insights into Cerasus species[J].Hortic Res,2020,7:165.DOI: 10.1038/s41438-020-00382-1.
[71]NIE C R,ZHANG Y J,ZHANG X Q,et al.Genome assembly,resequencing and genome-wide association analyses provide novel insights into the origin,evolution and flower colour variations of flowering cherry[J].Plant J,2023,114(3):519-533.DOI: 10.1111/tpj.16151.
[72]JIANG D Y,LI X K,LI Y G,et al.Chromosome-level assembly of flowering cherry (Prunus campanulata) provides insight into anthocyanin accumulation[J].Genes,2023,14(2):389.DOI: 10.3390/genes14020389.
[73]柳新红,孙中帅,蒋冬月,等.我国樱花产业主要问题的思考[J].浙江林业科技,2018,38(6):77-82.LIU X H,SUN Z S,JIANG D Y,et al.About challenges of flowering cherry industry in China[J].J Zhejiang For Sci Technol,2018,38(6):77-82.DOI: 10.3969/j.issn.1001-3776.2018.06.014.
[74]孙中帅,陈超,黄升,等.樱亚属野生种资源分布及分类研究[J].浙江林业科技,2022,42(5):66-75.SUN Z S,CHEN C,HUANG S,et al.Distribution of wild Prunus subg.Cerasus and inquiry into taxonomy[J].J Zhejiang For Sci Technol,2022,42(5):66-75.DOI: 10.3969/j.issn.1001-3776.2022.05.009.
[75]FU C L,XIE C P,IDREES M,et al.The species richness-environment relationship for cherries (Prunus subgenus Cerasus) across the Northern Hemisphere[J].Forests,2023,14(2):193.DOI: 10.3390/f14020193.
[76]LINDLEY J. Transactions of the Horticultural Society of London(volume VII) [M]. London: M. Bulmer amp; Co, 1830.
[77]KOIDZUMI G. Notes on Japanese Rosaceae[J]. Bot. Mag. (Tokyo), 1911, 25: 184-188.
[78]吴保欢.基于形态性状的中国李属樱亚属系统分类研究[D].广州:华南农业大学,2016.WU B H.Taxonomy revision and phylogenetic study based on numerical tax-onomy on Prunus L.subgenus Cerasus (Mill.) A.gray in China[D].Guangzhou:South China Agricultural University,2016.
[79]WU B H,LIU C Y,POTTER D,et al.Taxonomic reconsideration of Prunus veitchii (Rosaceae)[J].PhytoKeys,2019,115:59-71.DOI: 10.3897/phytokeys.115.29219.
[80]张毅,史传铎.山东樱属一新变种:泰山野樱花[J].植物分类学报,1999,37(1):87.ZHANG Y,SHI C D.A new variety of sakura from Shandong: wild sakura Taishan[J].J Syst Evol,1999,37(1):87.
[81]KATSUKI T,IKETANI H.Nomenclature of Tokyo cherry (Cerasus × yedoensis ‘Somei-yoshino’,Rosaceae) and allied interspecific hybrids based on recent advances in population genetics[J].Taxon,2016,65(6):1415-1419.DOI: 10.12705/656.13.
(责任编辑 吴祝华)
猜你喜欢
课堂内外·教师版(2022年4期)2022-05-23 22:23:32
上海师范大学学报·自然科学版(2021年1期)2021-03-30 05:42:11
广西植物(2020年10期)2020-12-14 03:59:42
广西植物(2020年10期)2020-12-14 03:59:42
湖北农业科学(2019年16期)2019-09-20 04:46:40
农村经济与科技(2018年16期)2018-05-14 17:05:51
图书馆界(2016年3期)2016-07-27 06:38:37
中央民族大学学报(自然科学版)(2016年2期)2016-06-27 01:28:46
新疆大学学报(哲学社会科学版)(2015年6期)2015-10-12 02:59:52
中央民族大学学报(自然科学版)(2015年3期)2015-06-11 02:13:54
