
不同浓度根皮苷对苹果砧木根系生长及生理特性的影响
2024-06-11 20:39:45李博洋任胜寒刘怀特王月张学英徐继忠
山东农业科学 2024年3期
李博洋 任胜寒 刘怀特 王月 张学英 徐继忠
摘要:本试验以苹果砧木组培苗为试材,设置O(CK)、0.1、0.5、1.0mmol/L和1.5mmol/L共5个根皮苷浓度处理,研究砂培条件下各处理对苹果砧木组培苗生长、根系构型及根系生理生化特性的影响。结果表明,与CK相比,外施不同浓度根皮苷均抑制苹果砧木组培苗根系生长,显著降低组培苗鲜重和干重,均显著降低根长、根表面积、根体积、根平均直径和株高净增长量(0.1mmol/L浓度处理除外),且随着根皮苷浓度升高而降幅增大。外源根皮苷胁迫导致苹果砧木组培苗根系超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性降低,与根系根皮苷代谢相关的查尔酮合酶、查尔酮异构酶及根皮苷水解酶活性显著降低,丙二醛(MDA)、超氧阴离子、脯氨酸含量显著升高,可溶性蛋白含量显著降低。综之,随着根皮苷浓度增加,苹果砧木组培苗根系抗氧化保护性酶活性和可溶性蛋白含量降低,MDA、超氧阴离子含量及脯氨酸含量升高,根系生长受到抑制,从而抑制苹果砧木组培苗生长。
关键词:根皮苷;苹果砧木组培苗;根系构型;抗氧化酶活性
中图分类号:S661.101 文献标识号:A 文章编号:1001-4942(2024)03-0048-06
同一地点连续多年种植同一种作物,即使是在正常栽培管理条件下,也会对相同或近缘植物的生长产生抑制作用,此现象称为连作障碍,又称为再植障碍、重茬障碍,连作障碍严重影响作物的生长发育,导致根系发育不好、分生能力差及地上部生长缓慢、植株矮小、叶面积变小等症状,很多老果园因此会出现更新再植问题,严重限制着果树产业的健康发展。作物根系分泌物以及前茬作物腐解产生的酚酸类有害物质是引起再植病害的主要原因之一。有研究表明根系分泌物会导致叶片气孔导度降低、蒸腾作用减弱和根长、干重和鲜重减少。酚酸类物质会影响根的生长和形态,使叶绿素含量降低。
根皮苷是苹果属含量最高的酚类化合物,是引起重茬障碍的关键酚酸类物质。根皮苷与作物生长发育和抗逆性有着密切关系。王婧茹研究表明根皮苷積累能够影响苹果植株体内生长素的分布。张素素等研究表明,一定浓度的根皮苷可使桃苗酶活性降低,丙二醛含量上升。尹承苗等研究表明,根皮苷、根皮素和酚酸类等5种物质均降低了平邑甜茶幼苗生长量、根系线粒体相关功能指标和相关酶活性指标。张江红等用1.0mmol/L根皮苷处理平邑甜茶幼苗,发现幼苗的根系长度和表面积大幅下降,根尖的超微结构受到破坏,根系的吸收能力受到严重影响。根皮苷浓度0.001mmol/L可促进平邑甜茶幼苗生长,但浓度达到1mmol/L时开始抑制平邑甜茶幼苗生长,降低干物质积累,且随着处理浓度增加抑制作用加强。王青青等研究发现一定浓度的根皮苷会使平邑甜茶TCA循环相关的酶活性降低,使根系呼吸速率下降。
前人的大部分研究主要集中于根皮苷对平邑甜茶幼苗生长的影响,而关于不同浓度根皮苷对苹果组培苗根系生长及其相关生理变化的系统研究较少。本试验以苹果砧木组培苗为试材,在沙培条件下研究不同浓度根皮苷对其生长、根系构型、根系酶活性、根系渗透调节物质含量的影响,为进一步揭示根皮苷的作用机制提供理论依据。
1 材料与方法
1.1 试验材料
2019年10月采集八棱海棠及其他砧木的果实,剥种沙藏,2020年春播种,2021年将这些砧木实生苗定植于保定市满城区抗重茬试验园。该园上茬苹果品种为新红星,树龄30年,于2021年春季刨树。2021年6月20日挑选长势良好的植株并挂牌,采集外植体带回河北农业大学进行组培快繁。2022年6月,选择已生根移栽且长势一致的编号为20号的组培苗(冀砧3号后代)作为试材,栽植于12cm×12cm营养钵内进行砂培,并用Hoagland营养液每隔2d处理一次,待长至6片左右真叶,用不同浓度根皮苷进行处理。
1.2 试验设计与处理
试验采用随机区组设计,重复5次,每重复6株,每处理30株。试验共设置4个根皮苷浓度处理:0.1mmol/L根皮苷(T1)、0.5mmol/L根皮苷(T2)、1.0mmol/L根皮苷(T3)、1.5mmol/L根皮苷(T4),以浇施等量清水为对照(CK)。每隔2d处理1次,每次100mL,共处理5次,共处理15d。于第5次处理后30d取样测定各项生理生化指标。
1.3 测定指标与方法
每隔10d用卷尺测量株高。于第5次处理后30d取样,用千分之一电子天平分别称取根、茎、叶鲜重,烘干后称干重,并计算总生物量。试验结束前,每个处理均收获植株根系,利用Win-RHIZO图像扫描系统量化根长、根表面积、根体积、根直径、根尖数和根分叉数。
采用氮蓝四唑光还原法测定超氧化物歧化酶(SOD)活性,采用愈创木酚法测定过氧化物酶(POD)活性,采用紫外吸收法测定过氧化氢酶(CAT)活性。采用考马斯亮蓝法测定可溶性蛋白含量,采用硫代巴比妥酸法测定丙二醛含量,采用羟胺氧化法测定超氧阴离子含量,采用酸性茚三酮显色法测定脯氨酸含量。使用试剂盒(上海晶抗生物技术有限公司产品)测定查尔酮合酶、查尔酮异构酶和根皮苷水解酶活性。
1.4 数据处理与分析
用Microsoft Excel进行数据处理,用Sigma-plot 14.0软件作图,用DPS数据处理软件进行方差分析与显著性检验(Duncan's新复极差法)。
2 结果与分析
2.1 不同浓度根皮苷对苹果砧木组培苗生长的影响
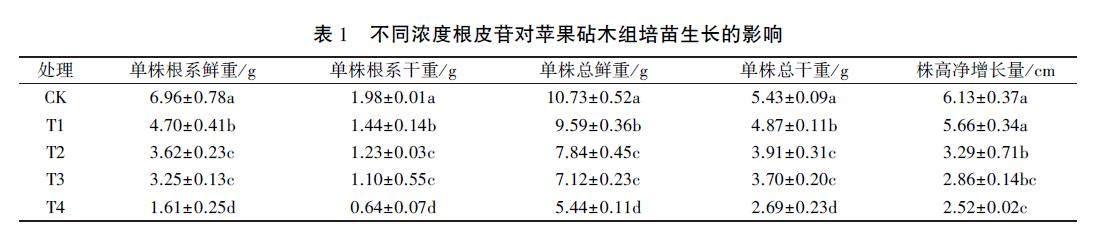
由表1可以看出,根皮苷处理显著抑制组培苗生长,对根系的抑制作用更明显。随着根皮苷浓度增加,各处理组培苗单株总鲜重、总干重、根系鲜重、根系干重与株高净增长量与CK相比,除T1处理株高净增长量外均为显著下降,T2和T3处理间各指标差异均不显著。其中,与CK相比,T1、T2、T3、T4处理组培苗单株总鲜重分别降低10.62%、26.93%、33.64%和49.30%,单株总干重分别降低10.31%、27.99%、31.86%和50.46%,单株根系鲜重分别降低32.47%、47.99%、53.30%和76.87%,单株根系干重分别降低27.27%、37.88%、44.44%和67.68%,株高净增长量分别降低7.67%、46.33%、53.34%和58.89%。
2.2 不同浓度根皮苷对苹果砧木组培苗根系构型的影响
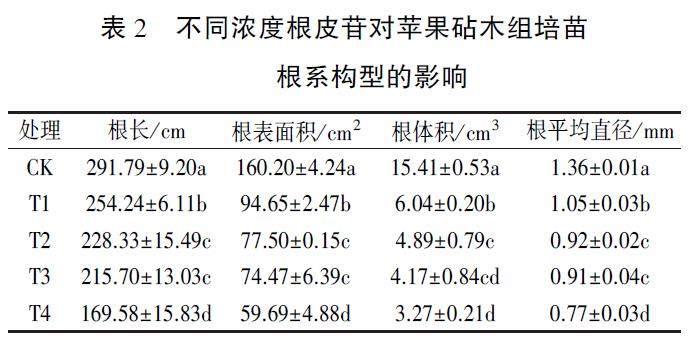
由表2可知,随着根皮苷浓度增加,苹果砧术组培苗的根长、根表面积、根体积和根平均直径对其响应越加剧烈,且除T2、T3处理问差异不显著外,不同处理间响应程度存在显著差异。与CK相比,T1处理根长、根表面积、根体积和根平均直径分别降低12.87%、40.92%、60.80%和22.79%,T2各指标分别降低21.75%、51.62%、68.27%和32.35%,T3各指标分别降低26.08%、53.51%、72.94%和33.09%,T4各指标分别降低41.89%、62.74%、78.78%和43.38%。
2.3 不同浓度根皮苷对苹果砧木组培苗根系酶活性的影响
2.3.1 对组培苗根系抗氧化酶活性的影响
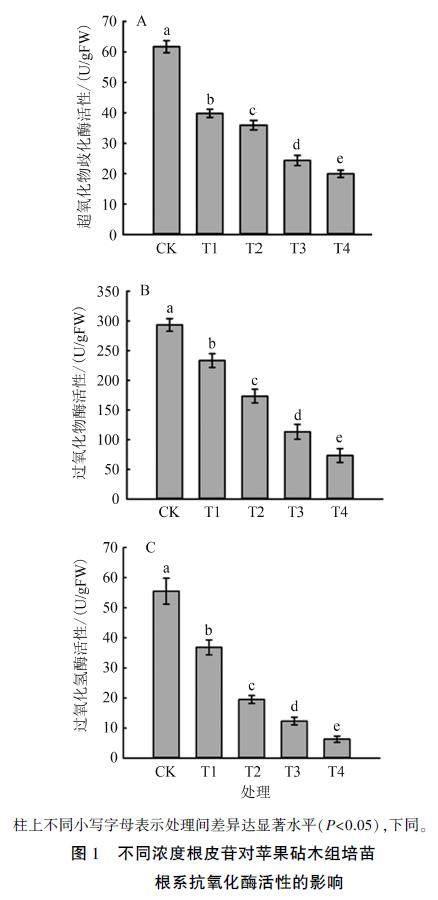
由图1A可知,苹果砧术组培苗根系超氧化物歧化酶(SOD)活性随着根皮苷浓度增加而降低。T1、T2、T3、T4处理组培苗根系SOD活性较CK分别显著降低35.50%、41.75%、60.55%、67.64%。
由图1B可知,苹果砧木组培苗根系过氧化物酶(POD)活性随着根皮苷浓度增加而降低。T1、T2、T3、T4处理POD活性较CK分别显著降低20.45%、40.91%、61.36%、75.00%。
由图1C可知,苹果砧木组培苗根系过氧化氢酶(CAT)活性受外源根皮苷干扰较大,随着根皮苷浓度增加,CAT活性呈现出大幅下降趋势。T1、T2、T3、T4处理CAT活性较CK分别显著降低33.63%、64.86%、77.78%、88.59%。
2.3.2 对组培苗根系根皮苷代谢相关酶活性的影响
由图2A可知,随着外源根皮苷浓度逐渐增加,苹果砧木组培苗根系查尔酮合酶活性呈现出下降趋势,表现为CK>T1>T2>T3>T4,其中T1与T2、T3与T4处理间差异不显著。与CK相比,T1、T2、T3、T4处理查尔酮合酶活性分别显著降低13.87%、36.35%、39.52%、45.01%。
由图2B可知,随着外源根皮苷浓度逐渐增加,苹果砧木组培苗根系查尔酮异构酶活性呈现出逐渐下降趋势。T1、T2、T3、T4处理查尔酮异构酶活性较CK分别显著降低12.01%、12.85%、30.52%、41.45%,但Tl与T2、T3与T4处理间差异不显著。
由图2C可知,对照(CK)苹果砧木组培苗根系根皮苷水解酶活性显著高于各处理,达到1.52IU/L,T1、T2、T3处理之间差异不显著,但均显著高于T4。与CK相比,T1、T2、T3、T4处理根皮苷水解酶活性分别降低11.67%、14.29%、18.90%,52.86%。
2.4 不同浓度根皮苷对苹果砧木组培苗根系渗透调节物质及丙二醛和超氧阴离子含量的影响
由图3A可知,对照(CK)苹果砧术组培苗根系可溶性蛋白含量显著高于各处理,T1、T2、T3、T4处理可溶性蛋白含量较CK分别降低14.26%、17.14%、25.65%、37.82%。
由图3B可知,随着根皮苷浓度增加,苹果砧木组培苗根系丙二醛(MDA)含量逐渐增加,与CK相比,T1、T2、T3、T4处理的MDA含量分别显著提高20.48%、36.42%、52.85%、71.11%。
由图3C可知,添加外源根皮苷后,T1、T2、T3、T4处理苹果砧木组培苗根系超氧阴离子含量较CK分别提高15.40%、37.53%、57.89%、84.26%,均达显著水平。
由图3D可知,T4处理苹果砧术组培苗根系脯氨酸含量最高,达到426.7μg/g,湿著高于CK和其他处理。T1、T2、T3、T4处理的脯氨酸含量较CK分别显著提高17.54%、32.64%、81.47%、118.72%。
3 讨论
外源添加酚酸类物质如根皮苷、肉桂酸、甲苯酸等会阻碍植物地上部的生长发育。施加一定浓度的根皮苷会明显抑制苹果幼苗的株高、茎粗、鲜重以及干重,同时,酚酸类物质也会抑制地下部的生长。研究表明,植株根系是植物感受土壤环境变化的第一部位,植物往往通过产生相应的变化使根系以适应变化后的环境,而根系形态是衡量根系功能效率和生理特性的重要指标。重茬胁迫会导致苹果根系结构受到破坏,鲜重和质量降低。本试验结果表明,外施根皮苷抑制苹果砧木组培苗生长,尤其是根系的生长,且随着根皮苷浓度的提高其总生长量逐渐降低,其中根系干重、鲜重及根长显著降低:同时,根皮苷浓度为0.1mmol/L时,苹果砧木组培苗的根长、根表面积、根体积和根平均直徑就显著低于CK,且随着根皮苷浓度的增加,其对4个生长指标的抑制作用加重。这说明根皮苷会不同程度地减少根系的吸收面积,根系的吸收功能受到抑制。
在正常条件下,植物细胞中自由基的产生与清除在抗氧化系统的参与下保持着动态平衡。当植物处于外界不利环境时,其自由基与抗氧化酶系统的动态平衡会被打破,从而对植物造成伤害。SOD、POD、CAT活性是抗氧化酶系统中的重要指标,这三种酶可以保护植物清除产生的自由基,以减轻伤害。在重茬胁迫下,苹果幼苗叶片的抗氧化酶活性降低;连作导致平邑甜茶幼苗抗氧化酶活性降低,同时游离脯氨酸含量增加,加剧了膜脂过氧化程度。本试验结果表明,外施根皮苷导致根系内SOD、POD、CAT活性降低,表明抗氧化酶系统与活性氧之间的平衡被打破。查尔酮合酶、查尔酮异构酶和根皮苷水解酶通过增加基因表达量而达到清除ROS提高抗性的作用。随着根皮苷浓度的提高,查尔酮合酶、查尔酮异构酶和根皮苷水解酶活性显著降低,说明施用根皮苷后,幼苗无法及时清除体内的ROS,加剧了根皮苷对组培苗的伤害。
细胞质膜是植物应对感知外界环境的第一层保护膜。植物受到胁迫时,会导致膜脂过氧化或膜脂脱脂作用,从而破坏膜结构。可溶性蛋白和脯氨酸是重要的渗透调节物质和营养物质,其积累是植物在胁迫下的一种适应性机制,以维持细胞内外渗透平衡。发生胁迫后,会导致MDA、超氧阴离子、脯氨酸含量升高。本试验结果显示,根皮苷处理的苹果砧木组培苗根系可溶性蛋白含量降低,MDA、超氧阴离子含量和脯氨酸含量上升,膜脂过氧化程度加剧,植株代谢失衡,从而抑制苹果砧木组培苗生长。
4 结论
本研究得出,随着根皮苷浓度增加,苹果砧木组培苗生长尤其根系生长受到抑制,同时导致抗氧化保护酶和根皮苷代谢相关酶活性、可溶性蛋白含量降低,MDA、超氧阴离子含量及脯氨酸含量升高,从而抑制苹果砧木组培苗生长。
基金项目:国家苹果产业技术体系项目(CARS-27);河北省重点研发计划项目(21326308D-02-03);河北省现代农业产业技术体系水果创新团队项目(HBCT2018100403)
