
在中国广东侵染胜红蓟的中国胜红蓟黄脉病毒的鉴定和全基因组分析
2024-06-08 07:07:43乔蕊周雪平李方方
植物保护 2024年3期
乔蕊 周雪平 李方方

摘要
2022年9月在廣东省罗定市发现了叶片表现为黄色网状症状的胜红蓟病株。为了明确胜红蓟叶片的黄脉症状是否由双生病毒感染引起,本研究使用检测双生病毒的简并引物PA/PB进行PCR扩增,获得约500 bp的片段。根据该序列设计特异性引物扩增并且克隆得到了病毒DNAA的全基因组序列。通过BLAST比对发现,获得的DNAA与中国胜红蓟黄脉病毒(ageratum yellow vein China virus, AYVCNV)海南分离物(OQ421190)的DNAA的相似性最高,相似度为98.11%。系统进化树分析显示,获得的病毒DNAA与海南分离物(OQ421190)在同一分支,说明具有较近的亲缘关系。以上研究结果表明侵染胜红蓟的病毒是AYVCNV的分离物。这是关于AYVCNV在广东地区侵染胜红蓟的首次报道,可为当地病毒病的防控提供参考。
关键词
中国胜红蓟黄脉病毒; PCR技术; 基因组序列; 系统发育分析
中图分类号:
S 432. 41
文献标识码: A
DOI: 10.16688/j.zwbh.2023216
Identification and complete genome sequence analysis of ageratuminfecting ageratum yellow vein China virus from Guangdong, China
QIAO Rui, ZHOU Xueping*, LI Fangfang*
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,
Chinese Academy of Agricultural Sciences, Beijing 100193, China)
Abstract
The ageratum plants with yellow vein symptoms were found in Luoding city, Guangdong province, in September 2022. To determine whether the ageratum plants showing vein yellowing symptoms were caused by geminiviruses, PCR technology was used to amplify about 500 bp sequence using the degenerate primers PA/PB for detecting the presence of geminiviruses in this study. Based on the amplified 500 bp sequence, the complete genome sequences of DNAA component were obtained by PCR amplification, cloning, and sequencing. BLAST comparison showed that the DNAA component shared the highest sequence identity (98.11%) with the Hainan isolate of ageratum yellow vein China virus (OQ421190). Phylogeny analysis showed that the complete genome sequence of DNAA component was most closely related to the Hainan isolate of AYVCNV (OQ421190). These results suggest that the virus infecting ageratum is an isolate of AYVCNV. This is the first report on AYVCNV infecting ageratum in Guangdong, which can provide a reference for local virus disease prevention and control.
Key words
ageratum yellow vein China virus; PCR technology; genome sequence; phylogeny analysis
双生病毒科Geminiviridae具有最大数量的植物DNA病毒,病毒粒子大小约为22 nm×38 nm[1],基因组大小2.5~5.2 kb,存在单组分或者双组分两种类型的单链环状基因组DNA。双生病毒依靠烟粉虱、叶蝉等昆虫介体传播[23],危害粮食、饲料、纤维、蔬菜、果树等作物,导致世界范围(特别是在热带和亚热带地区)内农业生产的巨大损失,有时甚至是毁灭性的损失。番茄、小麦、大豆、烟草、玉米和苹果等农作物都是双生病毒的重要寄主[45]。据报道,双生病毒在中国可侵染118种植物,其中75种为非农作物[6]。非农作物寄主为双生病毒提供了中转场所,利于双生病毒后续的持续侵染、传播和流行[7]。苋科、菊科、旋覆花科、锦葵科、大戟科、夹竹桃科、柳叶菜科、酢浆草科、紫茉莉科和车前科等科的非农作物均受到双生病毒的侵害。
胜红蓟Ageratum conyzoides是菊科Asteraceae藿香蓟属Ageratum植物,是双生病毒主要侵染的非农作物,在我国侵染胜红蓟的双生病毒主要有胜红蓟黄脉病毒(ageratum yellow vein virus, AYVV)、中国胜红蓟黄脉病毒(ageratum yellow vein China virus, AYVCNV)、四川胜红蓟曲叶病毒(ageratum leaf curl Sichuan virus, ALCScV)、胜红蓟曲叶病毒(ageratum leaf curl virus, ALCuV)、台湾胜红蓟黄脉病毒(ageratum yellow vein Taiwan virus, AYVTV)、赛葵黄脉病毒(malvastrum yellow vein virus, MYVV)、烟草曲茎病毒(tobacco curly shoot virus, TbCSV)和云南烟草曲叶病毒(tobacco leaf curl Yunnan virus, TbLCYV)等[811]。在这些病毒中,AYVCNV在我国的寄主范围较广,除侵染胜红蓟外,番茄、烟草和辣椒也是AYVCNV的常见作物寄主,近年在海南的雪茄烟上也分离到AYVCNV的病毒分离物[12]。
AYVCNV属于双生病毒科菜豆金色花叶病毒属Begomovirus的单组分病毒,其DNAA基因组编码6个蛋白。单组分双生病毒常伴有卫星分子协助其侵染,例如AYVV DNAA与卫星分子DNAβ的侵染性克隆可以共同侵染植物并且通过昆虫传到其他植物,引起黄脉症状。将AYVCNV确定为菜豆金色花叶病毒属的一个独立种是由于该病毒与其他种类菜豆金色花叶病毒的全长序列核苷酸相似性低于86%,并且该病毒的卫星分子DNAβ与其他种类菜豆金色花叶病毒的DNAβ分子的相似度低于81%[13]。DNAβ分子在单组分基因组双生病毒中充当DNAB的角色[14],在一些双生病毒中只有DNAA与DNAβ分子共同存在才会引起严重的病害症状。有研究证明,在棉花曲叶病株中不仅分离到了木尔坦棉花曲叶病毒(cotton leaf curl Multan virus, CLCuMuV),同时也分离到了类似的DNAβ分子,而且发现DNAβ的存在是棉花叶片产生卷曲症状的重要原因[15]。
本研究针对广东省罗定市番茄种植区周围胜红蓟病株存在的病原种类进行鉴定,避免存在的主要病原对番茄生长造成危害,为后期防治奠定基础。首先利用双生病毒DNAA的检测引物PA/PB对感病的广东胜红蓟进行检测,再通过设计背向引物获取全长基因组序列,根据获得的全长基因组序列对分离物分类并分析其遗传进化关系。利用DNAβ的简并引物Beta01/02检测是否存在DNAβ分子,从而确定引起广东胜红蓟黄脉症状的主要病原。
1 材料与方法
1.1 材料来源
2022年9月在广东省罗定市番茄种植区周围采集了疑似感染双生病毒的胜红蓟病株(图1),
病株叶片均表现为黄绿相间的花叶,叶脉褪绿发黄呈黄色网状,叶片还表现皱缩、上卷等症状(图1),将其
命名为202209GDSHJ。
图1 广东省胜红蓟样品202209GDSHJ的症状
Fig.1 Symptom of ageratum sample 202209GDSHJ
collected from Guangdong province
1.2 植物总DNA的提取及目的片段的扩增
采用CTAB法提取植物总DNA,以提取的胜红蓟样品DNA为模板,使用检测双生病毒的简并引物PA/PB(PA:5′TAATATTACCKGWKGVCCSC3′;PB:5′TGGACYTTRCAWGGBCCTTCACA3′)进行PCR扩增,扩增程序为:94℃预变性3 min;94℃变性30 s,56℃退火30 s,72℃延伸30 s,共32个循环;72℃延伸10 min。将扩增产物切胶纯化回收并且克隆至pGEMT easy vector。使用通用引物M13F/M13R检测阳性克隆并测序(北京擎科生物)。利用DNAβ的简并引物Beta01/02(Beta01:5′GGTACCACTACGCTACGCAGCAGCC3′;Beta02:5′GGTACCTACCCTCCCAGGGGTACAC3′)檢测伴随的DNAβ卫星分子,PCR扩增程序同上。
根据测序结果设计背向引物202209GDSHJF:(5′TTGATAAGGCTGATATTTGGGAAAGTG
CTT3′)和202209GDSHJR:(5′TATCAGCCTTATCAACCCCGACTAATAAAT3′)扩增该病毒的全长基因组序列,PCR产物通过琼脂糖凝胶电泳检测后纯化、克隆至pGEMT easy vector,筛选阳性克隆,测序(北京擎科生物)得到病毒的全长基因组序列。
1.3 病毒基因组序列分析以及进化树构建
得到全长基因组序列后,首先通过在线程序ORF finder(www.ncbi.nlm.nih.gov/orffinder/)确定病毒的开放阅读框;根据获得的开放阅读框,利用Snapgene绘制病毒的基因组结构;使用BLAST(blast.ncbi.nlm.nih.gov/Blast/)对获得的病毒分离物进行同源序列检索;使用SDT软件[16]对病毒分离物序列、我国不同省份鉴定的AYVCNV和AYVV病毒分离物以及部分在广东省鉴定的其他双生病毒分离物进行比对,并使用MEGA 7.0软件[17]对以上病毒分离物序列通过邻接法构建系统进化树,bootstrap设置为1 000。
2 结果与分析
2.1 胜红蓟疑似双生病毒侵染叶片的病毒检测
胜红蓟是很多植物病毒的中间寄主,于2022年9月在广东省罗定市番茄种植区周围的胜红蓟植株中发现了疑似感染双生病毒的病株。为了避免胜红蓟病株存在的病原对番茄造成危害,对其病原进行了分子鉴定。
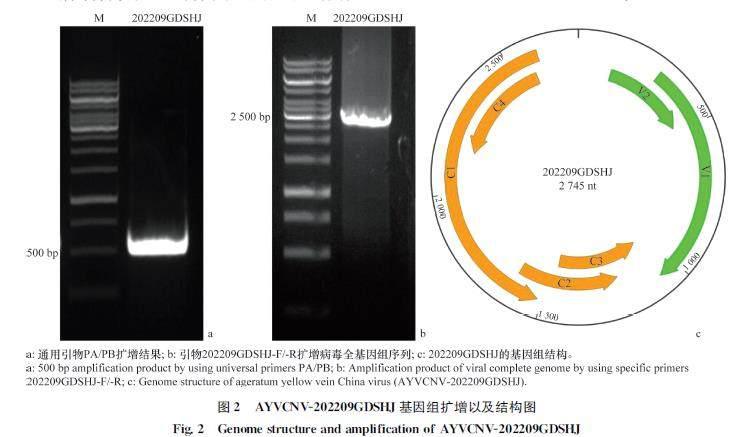
利用检测双生病毒的简并引物PA/PB 对疑似双生病毒侵染的胜红蓟病株样品的DNA进行扩增,得到约500 bp的特异性条带(图2a)。随机选取2个单克隆进行测序,结果表明它们具有99%的核苷酸序列相似性。利用DNAβ的简并引物Beta01/02在以上样品中没有检测到伴随的DNAβ卫星分子。
2.2 DNAA全长基因组序列扩增及结构分析
为了获得病毒的全长基因组核苷酸序列,在已经确定的500 bp序列基础上设计一对有15 bp重复序列的背向特异性引物,扩增病毒基因组全长(图2b)。挑取单克隆进行测序,得到2 745 nt的病毒基因组序列,上传GenBank获得登录号OQ819181。经过BLAST分析发现AYVCNV202209GDSHJ病毒分离物的基因组核苷酸序列与中国胜红蓟黄脉病毒海南分离物(AYVCNVDZ,登录号:OQ421190)的基因组核苷酸序列相似性较高,为98.11%。该分离物的基因组具有典型的单组分双生病毒基因组结构特征,尤其与已经报道的AYVCNV 的 DNAA 基因组高度一致,包含6个已知的编码蛋白(图2c):病毒链编码29.7 kD的V1 蛋白(294-1 069 nt)和13.6 kD的V2蛋白(134-484 nt),互补链上编码40.8 kD的C1 蛋白(1 516-2 604 nt)、15.5 kD的C2 蛋白(1 209-1 616 nt)、16.0 kD 的C3 蛋白(1 064-1 468 nt)和16.0 kD的C4 蛋白(2 154-2 576 nt),在病毒链与互补链之间包含有 AYVCNV 复制和转录必需元件的非编码区,其中包含一个9 nt的茎环结构(TAATATT/AC)。
2.3 AYVCNV202209GDSHJ与其他双生病毒的同源性分析
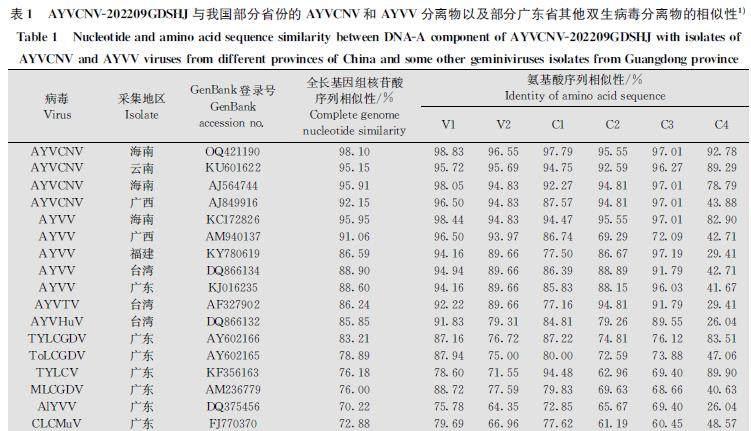
选取来自我国不同省份的AYVCNV和AYVV病毒分离物以及部分在广东省鉴定的其他双生病毒分离物,两两比较这些病毒分离物与AYVCNV202209GDSHJ的全长基因组核苷酸序列以及编码蛋白的氨基酸序列的同源性。结果表明,AYVCNV202209GDSHJ与AYVCNV(OQ421190)相似性最高,并且2个分离物基因组编码的蛋白的氨基酸相似性均最高,与我国其他地区的AYVCNV分离物的相似性在90%以上(表1)。
为了分析该病毒分离物的系统进化情况,将以上进行同源性比较的病毒分离物基因组全长核苷酸序列通过构建系统进化树(图3)分析该病毒分离物的遗传进化情况。从图中可以发现,本研究中來自广东的AYVCNV202209GDSHJ病毒分离物与来自海南的AYVCNV(OQ421190)亲缘关系较近,然而与同样采自广东的其他双生病毒分离物遗传距离较远。
3 结论与讨论
本研究对采自广东省胜红蓟样品进行双生病毒的检测和鉴定,发现AYVCNV在广东省侵染胜红蓟。根据国际病毒分类委员会(International Committee on Taxonomy of Viruses, ICTV)双生病毒科菜豆金色花叶病毒属分类原则,病毒全长基因组核苷酸序列相似度低于91% 即可划分为新种[18]。本研究分离到的AYVCNV202209GDSHJ与已经鉴定的AYVCNV(OQ421190)相似性最高,为98.10%,确定AYVCNV202209GDSHJ为双生病毒AYVCNV的一个分离物。根据Li等[6]2022年对中国双生病毒种类的发生与分布的研究总结,本研究是在广东省胜红蓟寄主上首次分离到AYVCNV分离物并且对其病毒基因组结构进行了分析。
通过对分离物AYVCNV202209GDSHJ编码产物氨基酸序列同源性分析发现,AYVCNV202209GDSHJ编码的6个蛋白的氨基酸序列与AYVCNV(OQ421190)编码的蛋白氨基酸序列高度相似,V1、V2、C1、C2和C3蛋白间的相似性均大于94%,C4的相似性为92.78%(表1)。
通过系统进化树的构建,在广东省采集鉴定的双生病毒分离物AYVCNV202209GDSHJ与海南省分离物AYVCNV(OQ421190)在同一分支,具有较近的亲缘关系,与同样是在广东省鉴定的其他双生病毒亲缘关系较远(图3)。出现这样的现象可能和昆虫介体对病毒的传播、作物栽培方式以及人类的活动导致病毒跨区域传播有关。
胜红蓟作为一种外来入侵杂草,最早只在中国香港发现,随着生物活动范围的增加,逐渐在中国各地广泛生长[1920]。胜红蓟在田间可直接或者间接影响农作物生长,在田间与农作物竞争生长环境。有调查表明,胜红蓟在贵州田间已成为优势杂草种群[21]。在广东省胜红薊可以与多种作物构成优势生物群落,例如茶树日本耳草胜红蓟群落和荔枝胜红蓟群落[22]。胜红蓟的整个生育期都与栽培植物伴生,感染病毒的胜红蓟能够作为病毒库,其是否携带病毒直接影响作物的健康。此外,胜红蓟还是大量病原物的重要寄主,尤其是病毒侵染后的胜红蓟可以作为中间寄主,为病毒提供一个合适的生存环境,在合适时机下病毒会侵染周围的农作物,从而造成农作物的经济损失。因此,对于杂草上双生病毒的检测是至关重要的,通过对侵染杂草的双生病毒的尽早防控有利于在源头上阻止双生病毒对农作物的侵染。
参考文献
[1] FIALLOOLIV E, LETT J M, MARTIN D P, et al. ICTV virus taxonomy profile: Geminiviridae [J/OL]. Journal of General Virology, 2021, 102(12): 001696. DOI: 10.1099/jgv.0.001696.
[2] LUAN Junbo, WANG Xiaowei, COLVIN J, et al. Plantmediated whiteflybegomovirus interactions: research progress and future prospects [J]. Bulletin of Entomological Research, 2014, 104(3): 267276.
[3] LU Quanyou, MA Yu, SMITH W K, et al. The identification of Tautoneura mori as the vector of mulberry crinkle leaf virus and the infectivity of infectious clones in mulberry [J]. Phytopathology, 2021, 112(2): 435440.
[4] LIU Yan, KHINE M O, ZHANG Peipei, et al. Incidence and distribution of insecttransmitted cereal viruses in wheat in China from 2007 to 2019 [J]. Plant Disease, 2020, 104(5): 14071414.
[5] DU Min, WANG Yongzhi, CHEN Cheng, et al. Molecular characterization and pathogenicity of a novel soybeaninfecting monopartite geminivirus in China [J/OL]. Viruses, 2022, 14(2): 341. DOI: 10.3390/v14020341.
[6] LI Fangfang, QIAO Rui, WANG Zhanqi, et al. Occurrence and distribution of geminiviruses in China [J]. Science Chinalife Sciences, 2022, 65(8): 14981503.
[7] MARWAL A, SAHU A K, GAUR R K, et al. Transmission and host interaction of Geminivirus in weeds [M]∥GAUR R K, HOHN T, SHARMA P. Plant virushost interaction: Molecular approaches and viral evolution. Boston: Academic Press, 2014: 143161.
[8] JIANG Tong, ZHOU Xueping. First report of malvastrum yellow vein virus infecting Ageratum conyzoides [J]. Plant Pathology, 2004, 53(6): 799.
[9] LI Pengbai, JING Chenchen, REN Hongyan, et al. Analysis of pathogenicity and virulence factors of ageratum leaf curl Sichuan virus [J/OL]. Frontiers in Plant Science, 2020, 11: 527787. DOI: 10.3389/fpls.2020.527787.
[10]WANG Xiaoyang, XIE Yan, ZHOU Xueping. Molecular characterization of two distinct begomoviruses from papaya in China [J]. Virus Genes, 2004, 29(3): 303309.
[11]孫少双,王冬雪,杨秀玲,等. 胜红蓟上一种与双生病毒伴随的重组β卫星分子的鉴定[J]. 植物保护, 2020, 46(4): 1924.
[12]陈德鑫,夏长剑,唐着宽,等. 中国胜红蓟黄脉病毒侵染海南雪茄烟的首次报道[C]∥中国植物病理学会. 植物病理科技创新与绿色防控——中国植物病理学会2021年学术年会论文集. 北京: 中国农业科学技术出版社,2021: 6.
[13]XIONG Qing, FAN Sanwei, WU Jianxiang, et al. Ageratum yellow vein China virus is a distinct begomovirus species associated with a DNA beta molecule [J]. Phytopathology, 2007, 97(4): 405411.
[14]SAUNDERS K, BEDFORD I D, BRIDDON R W, et al. A unique virus complex causes ageratum yellow vein disease [J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(12): 68906895.
[15]BRIDDON R W, MANSOOR S, BEDFORD I D, et al. Identification of DNA components required for induction of cotton leaf curl disease [J]. Virology, 2001, 285(2): 234243.
[16]MUHIRE B M, VARSANI A, MARTIN D P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation [J/OL]. PLoS ONE, 2014, 9(9): e108277. DOI:10.1371/journal.pone.0108277.
[17]KUMAR S, STECHER G, TAMURA K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets [J]. Molecular Biology and Evolution, 2016, 33(7): 18701874.
[18]BROWN J K, ZERBINI F M, NAVASCASTILLO J, et al. Revision of begomovirus taxonomy based on pairwise sequence comparisons [J]. Archives of Virology, 2015, 160(6): 15931619.
[19]吳海荣,胡学难,强胜,等. 广州地区胜红蓟物候学观察与调查研究[J]. 杂草科学, 2010, 28(3): 1821.
[20]郝建华,强胜. 外来入侵性杂草——胜红蓟[J]. 杂草科学, 2004, 23(4): 5458.
[21]李祖任,胡楠,晏升禄,等. 贵州省烟田杂草名录[J]. 山地农业生物学报, 2013, 32(4): 314321.
[22]吴万春,宁洁珍. 广州地区栽培植物—农田杂草群落学特性 [J]. 生态科学, 1990(2): 7985.
(责任编辑:田 喆)
猜你喜欢
数学物理学报(2022年5期)2022-10-09 08:56:44
河北画报(2020年8期)2020-10-27 02:54:20
新农民(2020年22期)2020-08-31 01:32:08
科学咨询(2020年10期)2020-04-01 03:43:16
乡村科技(2019年25期)2019-02-21 07:40:48
现代装饰(2018年3期)2018-05-22 02:51:43
中华奇石(2016年9期)2016-12-15 14:20:46
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:51
火花(2016年7期)2016-02-27 07:46:14
中国艺术时空(2014年2期)2014-02-28 21:32:09
