
神黄豆种子休眠及解除方法研究
2024-06-08 00:21:03杨江敏李富荣余贵湘张恩向董诗凡蔡文良刀保辉
安徽农业科学 2024年10期
关键词:种子休眠
杨江敏 李富荣 余贵湘 张恩向 董诗凡 蔡文良 刀保辉

摘要 為探究神黄豆种子的休眠原因及解除方法,对神黄豆种子的生物学特性和萌发抑制物进行观测,采用热水、浓硫酸和赤霉素(GA3)浸泡处理,打破种子休眠。结果表明:神黄豆种子为椭圆形,种皮黄褐色,千粒重为378.66 g,含水量为9.97%,种子生活力为100%,发芽潜力较大。完整种子浸泡215 h后,吸水率仅为0.57%,透水性较差。种皮和种胚的甲醇浸提液,对白菜种子的萌发和幼苗根长的生长具有显著的抑制作用。经98%浓硫酸浸泡10 min,种子发芽率最高(53.33%),用不同温度热水浸泡至自然冷却,萌发促进作用较小,发芽率最高仅为5.56%,GA3浸泡处理种子未萌发。种皮吸水障碍是神黄豆种子休眠的主要原因。4 ℃层积过程中,可溶性糖含量变化较小,25 ℃层积过程中,可溶性糖含量先上升后下降,可溶性蛋白质和粗脂肪含量总体呈下降趋势,且25 ℃层积过程中的减少速率更快,说明种子解除休眠过程中需要消耗大量营养物质。
关键词 神黄豆;种子休眠;解除休眠
中图分类号 S722 文献标识码 A
文章编号 0517-6611(2024)10-0133-04
doi:10.3969/j.issn.0517-6611.2024.10.028
Study on Dormancy and Release of Cassia agnes Brenan Seed
YANG Jiang-min,LI Fu-rong,YU Gui-xiang et al
(Dehong Prefecture Forestry Science Research Institute,Ruili,Yunnan 678600)
Abstract In order to explore the dormancy reason and dormancy release method of Cassia agnes seeds,in this research,we mainly studied the biological characteristics and germination inhibitors of C.agnes seeds,hot water,concentrated sulfuric acid and gibberellin (GA3) were used to break the seed dormancy.The results showed that the seed was elliptic,the seed skin was yellowish-brown,smooth and hard,the 1 000-seed weight was 378.66 g,the water content was 9.97%,the seed viability was 100%,and the germination potential was great.After soaking the whole seeds for 215 h,the water absorption rate was only 0.57%,which was significantly different from that of seeds broken skin.The methanol extract from the seed coat and seed embryo of C.agnes had significant inhibitory effects on the germination of Chinese cabbage seeds and the growth of seedling roots.The germination rate was higher (53.33%) when the seeds were soaked in 98% concentrated sulfuric acid for 10 min.Soaking seeds in the water with different temperature until natural cooling had little promoting effect on germination,and the highest germination rate was only 5.56%.The seeds soaked in gibberellin (GA3) can not promote the germination of seeds.The main reasons of C.agnes seeds dormancy were the water absorption barrier.During the stratification at four temperature,the soluble sugar content had little change,in the process of stratification at twenty-five temperature,the soluble sugar content first increased and then decreased,while the soluble protein and crude fat contents showed a decrease trend and the rate of decrease at twenty-five temperature stratification was faster,indicating that a large amount of nutrients were needed in the process of seed dormancy removal.
Key words Cassia agnes Brenan;Seed dormancy;Broken dormancy
基金项目 热带珍贵树种种质资源库建设项目。
作者简介 杨江敏(1990—),女,云南芒市人,工程师,硕士,从事林业草原培育和经营研究。*通信作者,工程师,硕士,从事林业草原培育和经营研究。
收稿日期 2023-08-18;修回日期 2023-10-25
神黄豆(Cassia agnes Brenan)为豆科(Leguminosae)决明属(Cassia)乔木植物[1],产于云南、广西等省区以及中南半岛的山地林中,喜高温、阳光充足的环境[2]。神黄豆生长快,抗性强,树形优美,夏季盛开时花团锦簇,具有浓郁的芳香,花期较长,具有较高的观赏价值,是优良的园林绿化树种,同时还具有一定的药理作用[3-6]。神黄豆的合理开发与推广应用对于丰富城市园林景观及推进植物药理研究具有积极意义。神黄豆在云南地區有天然分布,多生于茂密的丛林中,自然条件下种子萌发比较困难,不利于进一步开发利用。目前,国内对神黄豆的研究主要集中在果实、茎叶和茎部所含化合物的分离鉴定、药理作用分析、引种栽培评价和树种分类鉴别上,而有关神黄豆种子休眠原因及解除方法的研究鲜见报道。Nikolaeva[7]将种子休眠分为4种类型:种皮的阻碍作用引起休眠,种子成熟时胚未发育完全,种胚自身生理所致,混合因素引起的休眠。机械损伤种皮、热水、浓硫酸、激素浸泡等方式能有效解除种子休眠[8-11],层积处理能促进种胚发育成熟,改变内源激素含量,增强种子内的呼吸作用等,打破种子休眠[12]。因此,笔者对神黄豆种子休眠原因、解除方法和层积过程中的生理指标变化进行研究,探索打破神黄豆种子休眠方法,以进一步丰富神黄豆植物学研究方面的新资料,为基础研究提供理论参考。
1 材料与方法
1.1 试验材料
以2019年10月采自云南省瑞丽市武甸村的神黄豆种子为试验材料,采种后敲打荚果,取出带瓤的种子,再剥去种瓤,得出净种,备用。该试验所用的种子均为同一家系。
1.2 试验方法
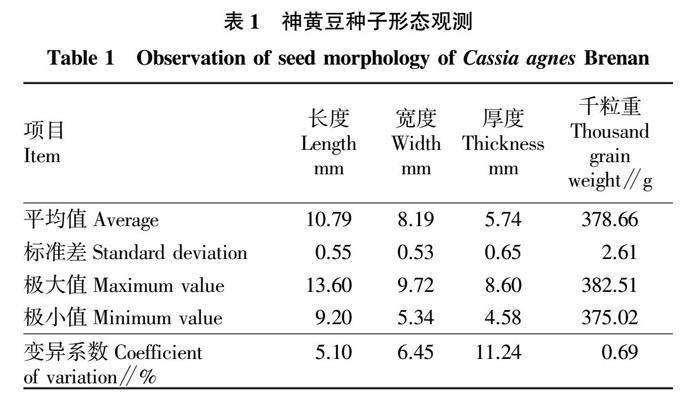
1.2.1 种子形态特征。
随机选取100粒种子,观察其形状、颜色和光滑度,总结种子形态特征。用游标卡尺测定种子的长度、宽度和厚度(精确到0.01 mm)。采用百粒法测量种子的千粒重[13],测定9组。用SPSS 13.0进行种子的形态变异分析。
1.2.2 种子含水量和吸水率。
随机选取20粒种子进行称重,记为W1,烘干至种子重量保持不变,记为W2,重复3次。种子含水量(%)=(W1-W2)/W1×100%。
分别选取完整种子、破皮种子各20粒,称重,于25 ℃的恒温水浴箱中浸泡,间隔4 h取出称重,夜晚8~10 h,直至重量稳定,依次记录。吸水率=[(湿重-干重)/干重]×100%。
1.2.3 种子生活力。
参考李炎坤等[14]的研究方法,采用氯化三苯基四氮唑(TTC)染色法测定种子生活力。随机取30粒种子,破皮处理后,用蒸馏水浸泡32 h,使种子充分吸胀,再将种子剖成2瓣,露出种胚,取其中一半放入0.1% TTC溶液中,避光染色12 h,染色结束后立即观察染色情况,对照种子用蒸馏水煮沸15 min,重复3次。
1.2.4 种子萌发特性。
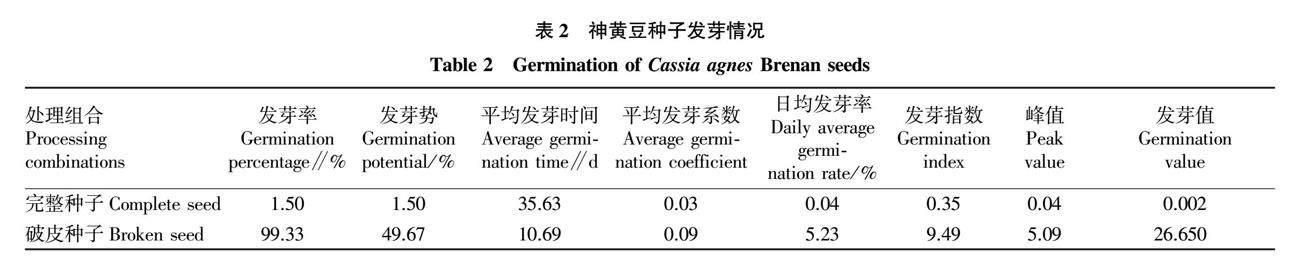
参考闫芳[15]的方法,选取600粒种子,将种子分为2组,每组300粒,其中1组擦破种皮。将完整种子和破皮种子分别置于铺有滤纸的培养皿中,黑暗条件下于培养箱(温度25 ℃,湿度10%)中进行萌发试验。种子发芽以露白为标志[16],萌发期内按时统计发芽率。参考蔡年辉等[17]的方法计算发芽率、发芽势、日均发芽率、平均发芽时间、平均发芽系数、发芽指数、峰值、发芽值。
1.2.5 种子内源抑制物质的粗提及生物测定。
参考赵敏等[18]的方法,将种子剥离为种皮和胚乳(含胚)2部分,分别制备0.1 g/mL浸提液,选取90粒白菜种子(云南祺合农业科技有限公司,醉贵妃Q-6,纯度95.0%,净度98.0%,发芽率85%以上),分为3组,每组30粒,均匀放置在培养皿中,分别加入4 mL不同浓度(50%、75%、100%)的浸取液,重复3次,在恒温培养箱(18 ℃)中进行萌发,以蒸馏水处理为对照,24 h后统计发芽率(当胚根长度达到种子长度的1/2时,记为发芽),72 h后测量白菜幼苗的茎长和根长。
1.2.6 不同处理方法对神黄豆种子萌发的影响。
参考前人研究方法[19-21],分别用3种不同溶液对种子进行处理,热水(100、90、80 ℃)浸泡至自然冷却;浓硫酸溶液(98%、80%、70%)浸泡10 min,期间用玻璃棒持续搅拌,处理后在流水下冲洗种子1 h;GA3溶液(700、500、300mg/L)避光浸泡72 h,每个处理90粒种子,每组30粒,重复3次,置于25 ℃恒温箱中进行萌发试验。以上试验以清水处理为对照,胚根突破种皮露白作为发芽标志,每天记录发芽情况。
1.2.7 不同温度层积处理对生理指标的影响。
取一定量的神黄豆种子放入50%多菌灵可湿性粉剂800倍液浸泡4 h,用清水冲洗后,按照3∶1的比例将种子与沙子混匀(含水量保持在50%左右),分别放入4和25 ℃的恒温箱中进行层积处理,每30 d取样1次,分别测定种子可溶性糖、可溶性蛋白质和脂肪的含量变化。可溶性糖含量采用蒽酮比色法测定,可溶性蛋白质含量采用考马斯亮蓝法测定,粗脂肪含量使用索氏提取法测定[22]。
1.2.8 数据分析。应用Microsoft Excel 2003进行试验数据处理,方差分析选用SPSS 13.0,采用Tukey法进行多重比较。
2 结果分析
2.1 种子生物学特性
2.1.1 種子形态特征。
神黄豆种子为椭圆形,种皮黄褐色,光滑坚硬,长度为(10.79±0.55)mm,宽度为(8.19±0.53)mm,厚度为(5.74±0.65) mm,种子厚度的变异系数最大,达11.24%,长度和宽度相对稳定,千粒重为(378.66±2.61)g,9组数据的变异系数为0.69%,无明显的变异趋势,颗粒重量较平均(表1)。
2.1.2 种子含水量。经测定,
神黄豆种子的平均干种子重7.782 g,平均烘干重7.005 g,平均含水量为9.97%,含水量变化较小。
2.1.3 种子生活力。经TTC染色法测定,参试的神黄豆种子生活力100%(图1),发芽潜力高。
2.1.4 种子萌发特性。
破皮处理和完整的神黄豆种子发芽率、发芽势、平均发芽系数、日均发芽率、发芽指数、峰值和发芽值差异极显著(P<0.01),平均发芽时间差异不显著(P>0.05)。破皮处理种子的平均发芽率、发芽势、发芽系数、日均发芽率、发芽指数、峰值和发芽值在发芽期内均大于完整种子,说明破皮处理能有效促进种子萌发(表2)。
2.2 神黄豆种子休眠原因
2.2.1 种子吸水率。破皮处理和完整种子吸水率差异极显著(P<0.01)。破皮种子吸水率随浸种时间的延长呈抛物线型变化,在0~48 h为快速吸水期,该阶段吸水率达到116.7%,占吸水总量的75.8%,浸种167 h后,吸水量逐渐趋于饱和;完整种子浸泡215 h后,吸水率仅为0.57%,说明种子存在吸水障碍(图2)。
2.2.2 种皮、种胚甲醇浸提液对白菜种子萌发和幼苗生长的影响。
由表3可知,种皮和种胚的甲醇浸提液对白菜种子的发芽率和幼苗根长的生长具有显著的抑制作用(P<0.05),且随着浸提液浓度的升高抑制作用逐渐增强。种胚甲醇浸提液对白菜种子的萌发抑制作用高于种皮,浸提液浓度为0.100 g/mL时,白菜种子的萌发率分别为15.56%和10.00%,相比CK分别下降了82.27百分点和88.61百分点,白菜幼苗的根长为3.27和5.46 mm,相比CK分别缩短了11.35和9.16 mm;种皮和种胚甲醇浸提液对白菜幼苗茎长的抑制作用不明显。
2.3 种子休眠解除方法
经100、90、80 ℃热水浸泡至自然冷却,各处理间差异不显著(P>0.05),100 ℃热水处理的种子发芽率、发芽势、发芽指数为5.56%、3.33%、0.23,萌发促进作用较小;98%、80%、70%浓硫酸处理10 min,期间用玻璃棒不断搅拌,98%浓硫酸处理与80%浓硫酸处理间发芽势的差异达到显著水平(P<0.05),98%浓硫酸、80%浓硫酸处理与70%浓硫酸处理间的发芽率和发芽指数差异显著(P<0.05),98%浓硫酸处理的种子发芽率、发芽势、发芽指数最高,分别达到53.33%、36.66%、2.37,总体发芽情况较好;经GA3浸泡的种子观测期内均未萌发(表4)。
2.4 不同温度层积过程中生理指标的变化
2.4.1 可溶性糖含量的变化。
从图3可知,不同温度层积处理可溶性糖含量变化差异极显著(P<0.01)。25 ℃层积过程中,0~30 d可溶性糖含量快速增加,30~60 d可溶性糖含量缓慢增加,在60 d时达到最大值86.7 mg/g,60~90 d可溶性糖含量迅速下降,120 d时下降至最小值3.3 mg/g。前期由于淀粉和脂肪的降解转化,可溶性糖含量逐渐增加,第60天开始,部分种子开始萌发,种子呼吸作用增强,可溶性糖被胚利用或被种子呼吸作用消耗而降低。4 ℃层积过程中可溶性糖含量变化较小。
2.4.2 可溶性蛋白质含量的变化。
从图4可知,不同温度层积处理蛋白质含量变化差异极显著(P<0.01),层积过程中,可溶性蛋白质含量总体呈下降趋势,且25 ℃层积过程中下降速率较快,可溶性蛋白质含量持续减少,说明可溶性蛋白质被水解利用,为种子萌发提供能量。
2.4.3 粗脂肪含量的变化。
由图5可知,不同温度层积处理粗脂肪含量变化差异极显著(P<0.01)。
4 ℃层积过程中,0~30 d粗脂肪含量下降较快,与层积前相比下降约42.2%;25 ℃层积过程中,0~30 d粗脂肪含量与层积前相比下降约67.7%。粗脂肪含量前期快速下降,后期变化幅度较小,且25 ℃层积过程中的下降速率大于4 ℃层积处理,说明粗脂肪不断分解转化,为种子萌发提供能量。
3 讨论与结论
种子萌发的第一步是吸水[23]。曹运梅等[9]报道了铜钱树种子果壳和种皮存在机械障碍,未经处理种子较难萌发。神黄豆完整种子浸泡215 h,吸水率仅为0.57%,说明种皮结构致密,透水性差,阻碍了种子的吸水。种子内源抑制物也是引起休眠的主要原因之一[24-25]。刘洋等[26]报道了乌苏里鼠李种子内部物质对种子萌发存在抑制作用。笔者在研究中发现,神黄豆种皮和种胚的甲醇浸提液对白菜种子的萌发和幼苗根长的生长具有显著的抑制作用,且随着浓度的升高抑制作用逐渐增强,说明神黄豆种皮和种胚均存在抑制白菜种子萌发的物质,至于该物质成分及其对神黄豆自身萌发是否具有抑制作用有待于后续研究。王付娟等[27-28]研究表明,热水和浓硫酸浸泡能够有效促进野生大豆、美丽决明和绒果决明种子萌发。神黄豆种子经不同温度热水浸泡至自然冷却,最高发芽率仅为5.56%;98%浓硫酸和80%浓硫酸浸泡处理种子10 min能有效提高种子萌发率;经不同浓度GA3溶液浸泡处理后,种子均未萌发,可能是种皮阻碍了激素的作用。赖小连等[29]研究表明,刻伤夏蜡梅的种皮能够有效提高发芽率。萌发试验中,擦破神黄豆种皮,种子萌发率达到99.33%,说明种皮的不透水性阻碍了种子萌发。淀粉、蛋白质、脂肪等种内贮藏物质在相应酶的作用下分解并转化助力种子萌发,为种胚及幼苗的成长提供必要的营养物质与能量[30]。层积过程中,可溶性糖、可溶性蛋白质和粗脂肪含量总体呈下降趋势,为种子萌发提供了能量来源。
综上所述,擦破种皮和98%浓硫酸、80%浓硫酸浸泡處理10 min,能够有效提高神黄豆种子的发芽率。种皮坚硬、透性差是导致神黄豆种子休眠的主要原因,种皮和种胚甲醇浸提液对白菜种子萌发和幼苗生长影响试验中,虽然存在一定的抑制作用,但对神黄豆种子休眠的影响还需进一步确定。可溶性糖、可溶性蛋白质和粗脂肪不断分解转化,表明贮藏物质参与并调节了神黄豆种子休眠解除的过程,但不同温度层积处理后种子发芽率与各生理指标的内在联系还需进一步试验和研究。
参考文献
[1] 《中国高等植物彩色图鉴》编委会等.中国高等植物彩色图鉴:第4卷[M].北京:科学出版社,2015:306-307.
[2] 胡勇,陈丽琼,朱霞,等.决明属化学成分与药理作用研究进展[J].现代中药研究与实践,2013,27(5):69-71.
[3] 张淑红.神黄豆[J].植物杂志,2003(6):28.
[4] 刘兴剑,汤诗杰,窦剑,等.几种热带苏木科观赏植物的引种观察[J].江苏农业科学,2009,37(5):193-195.
[5] 王闪闪,黄文忠,曾广智,等.神黄豆茎叶中酚性成分研究[J].云南民族大学学报(自然科学版),2019,28(2):105-108.
[6] 成功,龚济达,薛达元,等.云南省陇川县景颇族药用植物传统知识现状[J].云南农业大学学报,2013,28(1):1-8.
[7] NIKOLAEVA M G.Physiology of deep dormancy in seeds (Fiziologiya glubokogo pokoya semyan)[M].Jerusalem:Israel Program for Scientific Translations,1969.
[8] BASKIN J M,BASKIN C C.A classification system for seed dormancy[J].Seed science research,2004,14(1):1-16.
[9] 曹运梅,张建,向云,等.铜钱树种子休眠特性及破眠技术研究[J].种子,2019,38(8):73-76.
[10] 李兵兵,魏小红,徐严.麻花秦艽种子休眠机理及其破除方法[J].生态学报,2013,33(15):4631-4638.
[11] 程广有,唐晓杰,高红兵,等.东北红豆杉种子休眠机理与解除技术探讨[J].北京林业大学学报,2004,26(1):5-9.
[12] 孙跃春,樊奋成,张英俊.预冷打破种子休眠的研究进展[J].种子,2004,23(10):52-53.
[13] 国家技术监督局.农作物种子检验规程 其他项目检验:GB/T 3543.7—1995[S].北京:中国标准出版社,1996:11-12.
[14] 李炎坤,卓一南,曾湘达,等.岗梅种子生活力测定方法的优化[J].种子,2018,37(11):128-131.
[15] 闫芳.黄瑞香种子休眠机理研究[D].兰州:甘肃农业大学,2016.
[16] 魏琳,朱仁斌,程积民,等.不同处理对本氏针茅种子萌发影响的研究[J].种子,2012,31(7):25-27.
[17] 蔡年辉,许玉兰,邓丽丽,等.茎干通直与扭曲云南松种子萌发特性比较[J].西南林业大学学报,2017,37(5):9-14.
[18] 赵敏,王炎.膜荚黄芪种子萌发抑制物质特性的初步研究[J].中草药,2001,32(7):643-646.
[19] 武承云.不同处理方法对甘肃鸢尾种子萌发的影响[J].青海农林科技,2021(1):93-95.
[20] 罗小燕,李欣勇,孙英男,等.不同处理对山蚂蝗种子萌发和幼苗生长的影响[J].热带农业科学,2017,37(11):16-21.
[21] 赵冰,董进英,张冬林.温度、光照和赤霉素浓度对秀雅杜鹃种子萌发的影响[J].种子,2014,33(5):26-30.
[22] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:207-208.
[23] MURDOCH A J.Seeds:Ecology,biogeography,and evolution of dormancy and germination[J].Crop science,2000,40(2):564-565.
[24] 司倩倩,臧德奎,刘丹,等.玉铃花种子休眠原因及其内源激素含量变化[J].北方园艺,2017(6):91-95.
[25] 付楠,宋慧,王淑君,等.种子的休眠与破除研究进展[J].安徽农业科学,2018,46(24):10-12,15.
[26] 刘洋,郭娇,王娟,等.乌苏里鼠李种子休眠机理及萌发特性研究[J].西北农林科技大学学报(自然科学版),2020,48(12):127-135.
[27] 王付娟,刘书含,李淑梅,等.野生大豆种皮障碍休眠解除及萌发的研究[J].大豆科学,2019,38(5):733-739.
[28] 陈莹,孙苗苗,钟丽媚,等.不同浸种处理对美丽决明和绒果决明种子萌发的效果研究[J].种子,2019,38(5):70-73.
[29] 赖小连,颜立红,颜玉娟,等.夏蜡梅种子形态结构和种子萌发研究[J].种子,2021,40(2):86-90.
[30] 廖云娇,李雪,董学会.不同变温层积过程中东北红豆杉种子生理生化特性和胚形态的变化[J].中国农业大学学报,2010,15(1):39-44.
猜你喜欢
防护林科技(2020年11期)2020-12-30 03:55:36
林业科技(2020年6期)2020-12-07 06:05:36
现代农业科技(2019年20期)2019-12-05 02:45:02
江苏农业科学(2019年16期)2019-11-02 13:16:49
南方农业·下旬(2019年12期)2019-04-22 00:36:00
农家科技中旬版(2019年1期)2019-03-13 12:30:36
中国食品学报(2019年3期)2019-01-13 01:47:39
安徽农业科学(2018年6期)2018-05-14 08:59:42
天津农业科学(2018年3期)2018-03-24 09:59:46
江苏农业科学(2017年12期)2017-09-16 04:19:17
