
滨海复垦土地不同间混作模式增产及改良效应
2024-05-11 00:00:00朱奕豪李际峰董晓亮王松涛刘志全吴振陈为峰
山西农业大学学报(自然科学版) 2024年1期








摘要:[目的]通过研究高粱与野生大豆不同种植模式在滨海废弃盐田复垦土地上的作物增产及改良效应,探讨不同种植模式提高复垦土地生产力的机理及引起高粱根际土壤细菌菌群变化的主要环境因子,可为优化滨海废弃盐田复垦土地种植模式、种地养地相结合提供依据。[方法]在废弃盐田复垦形成的土地进行小区定位试验,设置高粱单作(S)、野生大豆单播(WS)、1 行高粱1 行野生大豆间作(S1WS1)、1 行高粱2 行野生大豆间作(S1WS2)、高粱与野大豆同行混播(SWS)共5 个处理,以高粱生物量、产量及根际土壤为研究对象,采用高通量测序技术,解析不同种植模式根际土壤细菌的菌群变化,并对细菌群落结构与土壤环境因子进行冗余分析。[结果]与高粱单作(S)相比,高粱-野生大豆间混作S1WS1、S1WS2、SWS 模式作物增产率分别为10. 16%、13. 15%、40. 68%,作物增产率与土壤硝态氮、碱性磷酸酶呈显著正相关,土地当量比均大于1,体现了间混作优势;S1WS1、S1WS2、SWS 模式提高了土壤有机质、氮磷养分含量、酶活性,降低了土壤pH 和含盐量,SWS 模式土壤改良效果最好;S1WS2、SWS 模式提高了土壤细菌多样性,SWS 模式细菌多样性指数最高,Ace、Chao、Shannon 指数分别为S 模式的1. 12 倍、1. 12 倍、1. 04 倍,细菌多样性指数与土壤硝态氮、脲酶呈显著正相关;放线菌门、变形菌门与绿弯菌门为丰度大于10% 的优势菌门,土壤硝态氮、碱性磷酸酶、氮磷比为影响细菌群落结构的主要环境因子。S1WS1、S1WS2、SWS 模式均提高了作物产量、土地当量比、土壤有机质和氮磷含量、酶活性。[结论]本试验条件下,相比于其它模式,SWS 模式作物增产率与土地当量比均最大,最有利于发挥间混作优势;土壤有机质和氮磷含量均最高,土壤改良效果最好。
关键词:种植模式; 种间相互作用; 作物增产; 土壤改良; 高通量测序
中图分类号:S154.3;S156.4 文献标识码:A 文章编号:1671-8151(2024)01-0089-12
我国土地资源禀赋不高,耕地质量总体偏低[1],废弃盐田复垦土地在经历了“农田-盐田-农田”的一系列演变过程后更加需要开展土地综合整治。盐田在卤水长期作用下,土壤中氯化钠等盐分不断积累,致使土壤钠离子浓度较高[2],而高盐、贫瘠的土壤环境会严重限制作物种类的选择。因此,具有耐盐碱、耐旱、耐涝、耐瘠薄等多重抗性的高粱[3-6]成为废弃盐田复垦土地农业种植的优势作物,受到农民的广泛推崇并大面积连年种植。
高粱连作的弊端会逐年加重,诸如土地养分偏耗、土壤微生境失衡,并加速土壤退化[7],不仅如此,高粱单作使得当地水土、气候及农业资源均得不到有效整合和合理利用,影响农业种植收益与生态效益。为此,亟需开展高粱与绿肥间套作种植模式研究,以缓解连作障碍,并达到用地养地相结合的目的。据统计,我国耕地有三分之二的播种面积采用间套作种植模式,全国粮食的50% 产量依靠间作套种获得[8]。在众多的间作套种方式中,豆科禾本科作物间作是较佳的种植模式,广泛应用于我国各地[9]。目前,豆科禾本科作物间作在玉米-大豆[7,10-12]、玉米-蚕豆[13]、玉米-苜蓿[11]、玉米- 花生[11,14]、小麦- 大豆[15]、小麦- 蚕豆[9,16-17]、燕麦-豌豆[18-19]、燕麦-花生[20]、甘蔗-大豆[21]中进行了较多的研究,但对高粱-豆科绿肥间作的研究鲜有报道,仅有少量研究是针对高粱-苜蓿轮作进行的[22]。另外,前人对高粱的研究主要集中于逆境胁迫[3-6]、栽培技术[23-25]、缓解或克服连作障碍[22,26-29]等方面,高粱(禾本科)-苜蓿(豆科)[22]轮作及甘蔗(禾本科)-大豆(豆科)[21]间作均可分别缓解高粱、甘蔗的连作障碍,还可以改善根际土壤环境,提高作物产量。研究人员已对不同禾本科-豆科作物间作进行了较多研究,对间作模式中作物的生长、产量、氮磷养分利用及土壤微生物的变化作出了合理的解释。但由于研究区域经历了“农田-盐田-农田”的复垦过程,与普通农田以及荒地、盐碱地等土地开垦的性质不同,在这种独特的环境条件下,难以在同一水平相互比较并进行规律总结。此外,高粱-野生大豆间作种植模式对废弃盐田复垦土地的改良效应及对作物的增产效应还未进行相关的研究。
本研究以废弃盐田复垦土地高粱-野生大豆不同间作模式为研究对象,系统研究高粱和野生大豆不同间作模式对废弃盐田复垦土地的改良效应及对作物的增产效应。通过分析作物产量、根际土壤理化性状并结合根际土壤细菌α 多样性与细菌群落组成的变化,探明不同间作模式下根际土壤理化性状与土壤优势菌群的相关性,明确高粱、野生大豆对土壤养分利用的竞争关系与种间互作效应,从而为废弃盐田复垦土地土壤改良和高效开发利用提供理论依据,为构建高产优质的间作模式提供技术支持。
1 材料与方法
1. 1 研究区概况
试验在山东省寿光市清水泊农场进行,试验田为2015 年由废弃盐田复垦成耕地,地理位置为37°13 ′48 ″ N,119°12 ′3 ″ E,该区属暖温带大陆性季风气候,雨热同季,年均气温12. 7 ℃,年均降雨量400~500 mm,多集中在6-8 月内。土壤类型为盐化潮土,土种为壤质重度卤化物盐化潮土,土壤质地类型以粉砂壤土为主,间有砂质壤土、黏壤土。试验田复垦当年即开始种植高粱,每年4 月底至6 月初播种,10 月中旬收获。
1. 2 试验设计
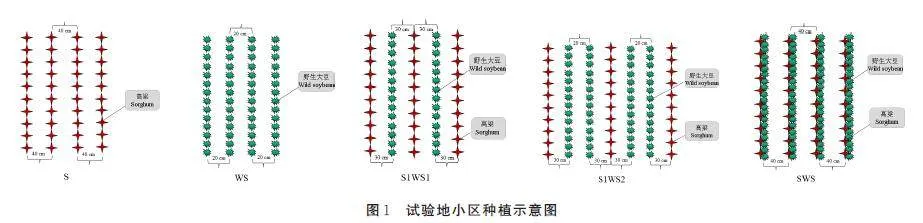
试验设置了高粱单作(S)、野生大豆单播(WS)、1 行高粱1 行野生大豆间作(S1WS1)、1 行高粱2 行野生大豆间作(S1WS2)、高粱与野生大豆同行混播(SWS)5 个处理,3 次重复,共15 个小区,小区面积为100 m2(10 m×10 m),种植模式见图1,间作比例和播种量见表1。供试品种为红缨子高粱(贵州红缨子农业科技发展有限公司生产)和野生大豆(取自黄河三角洲地区,为当地特有的野生资源)。2020 年5 月20 日播种,10 月15 日收获,高粱行距40 cm,高粱与野生大豆行距30 cm,野大豆行距20 cm,播种方式为行播。供试肥料为磷酸二铵(N-P2O5-K2O:15-42-0,贵州开磷集团股份有限公司生产),施肥量为560 kg·hm-2,播种时同时施加。
1. 3 取样方法与项目测定
每个种植小区随机量取2 m×2 m 样方3 个,齐地面砍倒作物,现场称重,对2 m×2 m 样方内的高粱株数、穗数进行记录,带回实验室后计算穗粒数、千粒重,进而换算成高粱公顷产量;计算野生大豆千粒重及2 m×2 m 样方总粒数,再换算成野生大豆公顷产量。间作增产率的计算方法为土地当量法,将间作区2 种作物产量分别与单种区比较再相加就得间作增产率[19]。
于高粱收获时取根际土壤样品,将高粱或野生大豆根系用铁铲从土壤中挖出(确保根系完整),慢慢抖掉与根系结合比较松散的土壤,然后用经过灭菌的镊子刮取附着在植株根系上的薄层(lt;10 mm)土壤作为根际土壤[21,30],10 株的根际土弄碎,混匀作为1 份土样,去除根系、树叶、石块等杂物后,做好标记,装于自封袋中。土样分3 部分,一部分取后立即放入冰盒保存,带回实验室保存于−80 ℃超低温冰箱,用于高通量测序;一部分新鲜土壤带回实验室保存于−20 ℃冰箱,用于土壤硝态氮、铵态氮测定;一部分自然风干后研磨,过100 目筛,用于土壤其他理化性状和酶活性测定。
烘干残渣法测定土壤盐分(SS);电极电位法测定土壤pH 值;重铬酸钾外加热法测定土壤有机质(SOM);半微量凯氏定氮法测定土壤全氮(TN);紫外分光光度法测定土壤NO3--N;靛酚蓝比色法测定土壤NH4+-N;高氯酸-硫酸法测定土壤全磷(TP);碳酸氢钠浸提-钼锑抗比色法测定土壤有效磷(AP);苯酚-次氯酸钠比色法测定脲酶(URE);3,5- 二硝基水杨酸比色法测定蔗糖酶(SUC);磷酸苯二钠比色法测定碱性磷酸酶(PHO)[31-32]。
Illumina MiSeq 测序及序列筛选[33]:扩增时在每个样品的上游引物5′端添加一段长度为8 个碱基的特异性多肽(barcode),用于区分样品。将扩增的nifH 基因PCR 产物经2% 琼脂糖凝胶电泳后回收,用NANO Quant(Tecan, Mnnedorf, Switzerland)测定浓度。将同一个处理的多个样品混匀,使各样品DNA 浓度一致后,用Illumina MiSeq测序平台进行双末端测序,测序服务委托上海美吉生物医药科技有限公司完成。
1. 4 种间相互作用的评价
1. 4. 1 土地当量比
土地当量比(LER)作为衡量土地利用效率大小的重要指标之一,当其gt;1 时,表明提高了土地利用效率,间作系统有间作优势;反之则表现为间作劣势[34-35]。LERB 与LERY 分别表示生物量土地当量比和产量土地当量比。
1. 4. 2 相对种间竞争力
相对种间竞争力(A)用于衡量间作系统中一种作物相对于另一种作物对光温水肥等自然资源竞争能力的大小,本文只计算了高粱相对于野生大豆的资源竞争能力大小(A),当其gt;0 表明高粱对资源的竞争能力大于野生大豆,当其lt;0 表明高粱对资源的竞争能力小于野生大豆。AB 与AY 分别表示高粱相对于野生大豆的生物量种间相对竞争力和产量相对种间竞争力。
1. 4. 3 相对拥挤系数
相对拥挤系数(K)用于表示间作与单作相比是否具有产量优势。当Kgt;1 表明间作与单作相比具有产量优势;当Klt;1 表明间作与单作相比没有产量优势。KB 与KY 分别表示生物量相对拥挤系数和产量相对拥挤系数。
1. 4. 4 相对竞争强度
相对竞争强度(RCI)表示特定作物组合的竞争能力,可根据作物产量或生物量计算得出。当RCIgt;0 表明种间竞争更高;当RCIlt;0 表明种内竞争更高;当RCI=0 表明种间竞争等于种内竞争。RCIS 与RCIWS 分别表示高粱产量相对竞争强度和野生大豆产量相对竞争强度。
1. 5 数据分析
采用Excel 2010 进行数据整理,用SPSS 22. 0进行数据显著性和相关性统计分析,用Origin 8. 0绘图,用Canoco 5. 0 进行冗余分析(RDA)。
2 结果与分析
2. 1 不同种植模式对高粱生物量与产量的影响
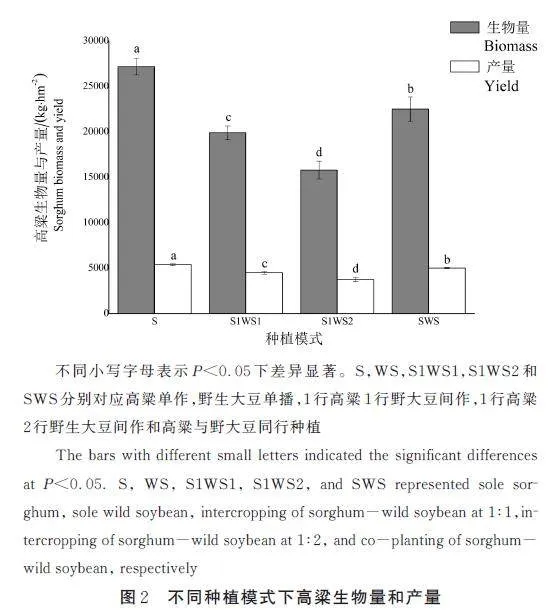
从图2 可以看出,不同种植模式对高粱的生物量与产量有显著影响(Plt;0. 05)。在3 种间作模式中,高粱的生物量和产量的高低与其种植占比相同,为SWSgt;S1WS1gt;S1WS2。相比S1WS1、S1WS2 模式,SWS 模式的高粱生物量分别增加了13. 08% 和42. 50%,产量分别增加了11. 52% 和33. 67%。土地当量法计算间作增产率,与S 相比,S1WS1、S1WS2、SWS 模式分别增产率为10. 16%、13. 52%、40. 68%。
2. 2 不同种植模式对作物种间相互作用的影响
如表2 所示,3 种间作模式的LERB、LERY 均大于1,最高值1. 44、1. 41 均为SWS 模式,其与S1WS1、S1WS2 模式相比差异显著;AB、AY 均大于0,最高值1. 01、1. 44 均为S1WS2 模式,KB、KY 均大于1,最高值均出现在SWS 模式中;RCIS 在S1WS1、S1WS2 模式中小于0,而在SWS 模式中大于0,RCIWS在3 种间作模式中均大于0。
2. 3 不同种植模式对根际土壤理化性质和酶活性的影响
由表3 可知,S1WS1、SWS 模式高粱根际土壤pH 高于高粱单作(S)模式,而S1WS2 模式高粱根际土壤pH 略微低于S 模式,5 种种植模式根际土壤pH 均偏碱性(7. 87~8. 03)。种植野生大豆(WS)后,与S 模式相比,其余4 种种植模式根际土壤含盐量均出现轻微下降。土壤SOM,AP,C/P,URE 和SUC 的最大值均出现在SWS 模式中,其SOM 和AP 含量、URE 和SUC 活性与S 模式相比,均达显著差异水平,分别是S 模式的1. 12 倍,1. 07 倍,1. 03 倍和1. 04 倍;土壤TN,TP,NO3-N,NH4-N 和N/P 的最大值均出现在WS 模式中,但其TN,TP 含量及N/P 与SWS 模式相比差异不显著;S 模式的C/N 为6. 67,是5 种种植模式中的最大值,但各处理间差异不显著。
2. 4 不同种植模式根际土壤细菌群落多样性与组成特征
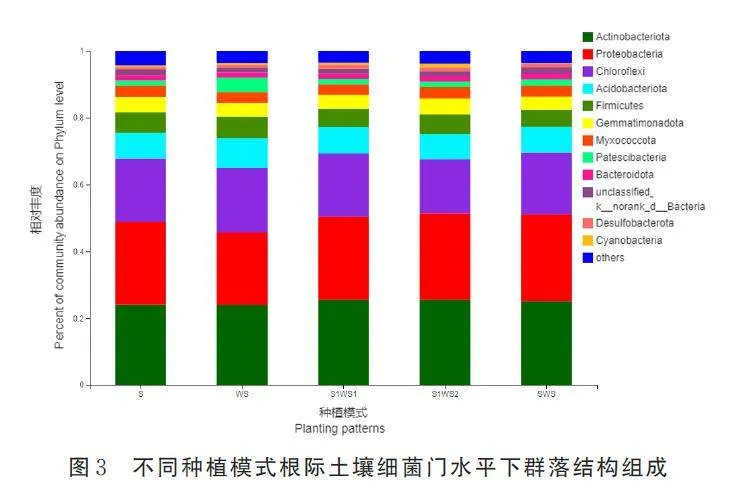
运用高通量测序技术对作物根际土壤进行16S 测序,5 种种植模式30 个样品经筛选得到有效序列1 755 385 条,平均长度为414 bp,以97% 的相似水平进行OTU 聚类,共获得7018 个OTU,分布在48 个细菌门,591 个细菌科,1082 个细菌属。5种种植模式的样品覆盖度在0. 967 4~0. 975 1,说明本研究获得的细菌序列覆盖度较好,其测序深度可以满足细菌群落结构组成及多样性分析。单因素方差分析结果显示(表4),不同处理间Ace 指数、Chao 指数差异显著(Plt;0. 05),Shannon 指数在SWS 模式与其余4 种模式相比差异显著。Ace指数、Chao 指数和Shannon 指数最大值均出现在SWS 模式中,分别为S 模式的1. 12 倍、1. 12 倍、1. 04 倍。
测序数据表明,所有样品中细菌群落的主要门为放线菌门(24. 03%~25. 51%,表示该菌门占细菌总序列百分比值)、变形菌门(21. 60%~25. 93%)、绿弯菌门(16. 19%~19. 27%)、酸杆菌门(7. 62%~9. 03%) 、厚壁菌门(5. 19%~6. 31%)、芽单胞菌门(3. 87%~4. 72%)、粘菌门(3. 27%~3. 57%) 、Patescibacteria 菌门(1. 50%~4. 37%)和拟杆菌门(1. 48%~1. 76%),这些门的相对丰度占细菌总序列的93. 91%~95. 06%(图3)。变形菌门和拟杆菌门在SWS 模式的相对丰度高于其余4 种种植模式,而厚壁菌门和芽单胞菌门在SWS 模式中含量最低,与S、WS、S1WS1 和S1WS2 模式相比,厚壁菌门分别降低了20. 08%、22. 68%、4. 32%、12. 43%,芽单胞菌门分别降低了14. 71%、5. 03%、8. 06%、21. 30%。
2. 5 高粱根际土壤细菌群落与环境因子之间的关系
为进一步分析不同种植模式高粱根际土壤环境因子对细菌门水平群落结构的影响,对细菌群落结构与根际土壤环境因子进行RDA 分析(图4)。结果表明:RDA 前两个排序轴的特征值分别为0. 540 和0. 133,分别解释54. 0% 和13. 3% 的细菌物种变化,本研究所选的14 个土壤环境因子的特征值,共解释了73. 5% 的总方差(表5)。其中,放线菌门、变形菌门、芽单胞菌门与有机质、全氮、全磷、硝态氮、铵态氮、有效磷、脲酶、蔗糖酶、碱性磷酸酶呈正相关,与pH、含盐量呈负相关;酸杆菌门、厚壁菌门与放线菌门、变形菌门、芽单胞菌门呈现相反的趋势。置换检验的结果显示(表6),硝态氮、氮磷比、蔗糖酶是主导细菌群落变化的主要因子。
3 讨论
3. 1 高粱与野生大豆间套作效应分析
间作套种在调节粮经饲作物用地矛盾,促进农林牧作物协调发展等方面发挥着重要作用[8]。其不仅可以提高耕地的土地利用效率,还有利于农田水、肥、气、热等资源的高效利用。豆科作物以其特有的固氮特性被作为绿肥资源长期使用,其生物固氮是自然界中最为高效的固氮系统[36],与豆科作物间作可以增产增收,提高总产出,是替代传统单作的优选种植模式[8]。本研究显示,与高粱单作(S)模式相比,S1WS1 模式、S1WS2 模式及SWS 模式中的作物增产率分别为10. 16%、13. 52%、40. 68%,作物增产率与硝态氮、碱性磷酸酶呈显著正相关(Plt;0. 05)(表7)。同时,S1WS1 模式、S1WS2 模式及SWS 模式的产量土地当量比(LERY)均大于1(表2),体现了间作优势。普遍认为,造成禾豆间作优势的重要原因是禾本科对豆科作物的养分掠夺增强了豆科作物自身的固氮作用[7],不仅如此,禾豆间作还能提高两者根系黄酮类物质的分泌,促进豆科作物结瘤固氮[37],这进一步明确了禾豆间作的优势。本研究中SWS 模式的增产率和LERY 均大于S1WS1 模式与S1WS2 模式,说明其间作优势优于S1WS1 模式与S1WS2 模式。有研究表明[7,16],禾本科-豆科间作中禾本科处于竞争优势地位,本研究结果与之相同,AB 和AY 均大于0(表2),表明高粱处于竞争优势地位,野生大豆处于竞争劣势地位。RCIS在S1WS1 模式与S1WS2 模式小于0,而在SWS模式中大于0,说明高粱在S1WS1 模式与S1WS2模式中以种内竞争为主,在SWS 模式中主要为种间竞争。RCIWS 在S1WS1、S1WS2 和SWS 这3 种模式中均大于0,表明野生大豆在3 种模式中均以种间竞争为主。在S1WS1、S1WS2 模式中,RCIS小于0,RCIWS大于0,说明高粱-野生大豆间作通过减轻高粱的种内竞争来提高高粱的生长速率,奠定了间作产量和生物量优势形成(LERgt;1、Kgt;1)(表2)。SWS 模式受两者根系伸展空间的限制[9],且野生大豆幼苗时期的根瘤尚未大量形成,固氮能力较弱,致使同行播种的高粱和野生大豆对养分竞争的影响较大。一般情况下,禾本科作物的养分竞争能力强于豆科作物,但在土壤养分尤其是有效氮含量低的环境中,豆科作物往往具有较强的种间竞争力[9],随着野生大豆地上与地下部生物量的增加,固氮能力增强,在高粱进入养分吸收高峰期后,高粱对养分的竞争力逐渐增强,两者也由养分竞争关系转变为互补关系,种间的互补效应对间作作物产量发挥着积极的作用。
3. 2 不同种植模式对废弃盐田复垦土地改良效应分析
耕层土壤质量与作物生长发育紧密相关,作物产量是耕层土壤各理化性状的综合体现[38]。为维持农业的可持续生产,协调作物的持续高产与土壤养分输入输出的动态平衡,必然要采取用地养地相结合的策略,但连年单作不仅起不到养地的效果,还会产生连作障碍,张爱加等[21]、张月萌等[39]的研究结果皆表明,与豆科作物间作可有效缓解连作障碍。
本研究中,结合土壤理化性状可以看出(表3),高粱与野生大豆间作套种后,对比高粱单作(S),高粱根际土壤的各养分含量均有显著提升,这是由于间作下野生大豆根际土壤中的养分通过质流和扩散运移至高粱根际土壤中[40-41]。已有研究表明,间作的豆科作物可产生直接或间接的氮素转移供给禾本科作物,而禾本科作物可以减缓氮素对豆科作物的“ 氮阻遏”现象,从而增加豆科作物的固氮量[41]。也就是说高粱竞争野生大豆根际养分,野生大豆根际氮素被高粱吸收,在根际氮素浓度降低时,会刺激野生大豆结瘤与固氮,使其固氮能力增强。对比S 模式,S1WS1、S1WS2、SWS 模式的根际土壤pH 降低,这是由于与野生大豆间作后,通过提高生物固氮释放H+、根系分泌物解离H+及有机磷的脱磷反应释放H+等降低根际土壤pH 值[41],从而改善该地区的土壤酸碱度,土壤中H+浓度及碱性磷酸酶活性的增加,可促进磷的活化,增加有效磷的含量,S1WS1、S1WS2、SWS 模式有效磷含量较S 模式分别增加0. 57%、6. 00%、7. 35%。同时,间作还能够提高土壤中微生物活性,促进腐殖质分解以增加有机质含量,有机质含量的增加有利于土壤团聚体数量增多,使土壤结构得到改善,这些都是高粱与野生大豆根际共生的结果。因此,高粱-野生大豆间混作可改善高粱根际土壤的养分状况,培肥地力。
土壤酶是土壤有机体的代谢动力,其活性大小可反映土壤有机质与养分转化的强弱。本研究中,高粱、野生大豆根际土壤的脲酶、蔗糖酶、碱性磷酸酶活性在野生大豆单播(WS)及S1WS1、S1WS2、SWS 模式中均大于高粱单作(S),研究结果与赵雅娇[41]的基本相似。脲酶活性可表征土壤氮素状况,蔗糖酶与土壤有机质、氮磷含量及微生物数量均有相关性,在间混作高粱根际土壤中,养分含量的增加尤其是碳氮养分的增加说明脲酶和蔗糖酶活性在禾豆间作系统下活性较大。碱性磷酸酶可促进土壤中的有机磷转化为无机磷,形成能被作物直接吸收利用的磷形态,而在本研究中S1WS2、SWS 模式下根际土壤有效磷含量显著大于单作,间作中野生大豆在高粱强竞争有效磷的情况下仍然表现出有效磷含量增加的趋势,进一步说明间作提高了禾豆间作系统中作物的碱性磷酸酶活性。所以,高粱-野生大豆间作可以提高高粱根际土壤酶活性,促进土壤的生物有效性。
已有研究证实,间作种植不仅能够改变根际土壤微生物的生长和繁殖,还可以使微生物的多样性及群落组成结构发生变化[42]。本研究中,S1WS2、SWS 模式的Ace 与Shannon 指数高于S模式,Chao 指数在S1WS1、S1WS2、SWS 这3 种模式中均高于S 模式,在禾本科牧草-紫花苜蓿间作[41]、小麦-苜蓿间作[43]等研究中也发现了相同规律,说明间混作对提高根际土壤微生物多样性具有普遍性。造成这种现象的原因可能是因为间作中的2 种作物因根系交错叠加作用使其根系在土壤中产生更多的碳水化合物、氨基酸和有机酸等碳源,分泌物的成分和含量发生变化,导致土壤微生物结构发生改变,从而影响根际土壤微生物的数量和多样性[44-45]。SWS 模式中的Ace 指数、Chao 指数、Shannon 指数要高于S1WS1 和S1WS2模式,表明同行播种更有利于提高土壤细菌多样性。相关性分析显示(表7),Ace 指数、Chao 指数、Shannon 指数与硝态氮、脲酶呈显著正相关,与pH呈显著负相关,说明土壤养分含量的增加、酶活性的提高及土壤质量的改善都会影响根际土壤微生物多样性的变化。
本研究发现的放线菌门、变形菌门、绿弯菌门、酸杆菌门、芽单胞菌门、厚壁菌门、拟杆菌门等微生物菌群在已有研究中也被证实普遍存在于盐碱土中[46],其中放线菌门、变形菌门和绿弯菌门是本研究根际土壤中的3 个主要类群(相对丰度大于10%),放线菌门和变形菌门在S1WS1、S1WS2、SWS 模式中的丰度均大于S 模式,放线菌门、变形菌门分别在S1WS1、SWS 模式中丰度最大,说明放线菌门和变形菌门是影响间混作优势的主要菌群。放线菌门菌群参与土壤有机质转化,其丰度大小可表示有机质含量的高低;变形菌门菌群的丰度与碳的利用方式有关,与土壤有机质的变化存在较高的相关性[41],本研究中2 类菌群丰度的变化均证实了根际土壤有机质含量在高粱-野生大豆间混作下高于其单作(S)的现象(表3)。RDA清晰地显示了影响根际土壤细菌群落的关键因子和影响程度(图4),放线菌门和变形菌门与土壤有机质、氮磷含量及酶活性呈正相关关系,而绿弯菌门和酸杆菌门的变化刚好与放线菌门和变形菌门的变化相反。因此,放线菌门和变形菌门的丰度越大,证明土壤质量越高;绿弯菌门和酸杆菌门的丰度越大会抑制土壤养分的积累,不利于地力提升与土地改良。本研究中根际土壤硝态氮、氮磷比、碱性磷酸酶是主导土壤细菌群落结构产生变化的主要因子(表6),表明不同种植模式的土壤理化性质不同,改变了微生物物种的组成和结构。
4 结论
本研究围绕滨海废弃盐田复垦耕地高粱与野大豆不同种植模式,分析不同模式下的增产效应及土地改良效应,结果表明:与高粱单作(S)相比,S1WS1、S1WS2、SWS 模式增产率分别为10. 16%、13. 52%、40. 68%,土地当量比都大于1,表现出明显的间作优势,作物增产率与硝态氮、碱性磷酸酶呈显著正相关,SWS 模式增产率最高,土地当量比最大;S1WS1、S1WS2、SWS 模式中高粱根际土壤有机质、氮磷含量增加,酶活性提高,pH和含盐量下降,土壤Ace 指数、Chao 指数、Shannon指数均增大,土壤硝态氮、脲酶与土壤细菌多样性呈显著正相关,土壤地力得到提升,土壤质量得到改善,土壤养分含量及微生物多样性在SWS 模式最大;放线菌门、变形菌门、绿弯菌门为丰富度大于10% 的优势菌门,放线菌门、变形菌门与土壤有机质、氮磷含量及酶活性呈正相关关系,其在S1WS1、S1WS2、SWS 模式中的丰度均增加,变形菌门在SWS 模式中的丰度最大,RDA 分析得出硝态氮、碱性磷酸酶、碳氮比为影响高粱根际土壤细菌群落变化的主要因子。综上,间混作野生大豆增加了高粱根际土壤生物多样性和土壤养分含量,改善了土壤生态环境,本研究条件下SWS 模式土地改良效果最好,土壤最健康。
参考文献
[1]王晓青. 黄淮海平原高标准农田项目建设后综合效益及影响因素研究[D]. 日照:曲阜师范大学, 2019.
Wang X Q. Study on comprehensive benefits and influencingfactors of construction projects of well-facilitated farmland inHuang-Huai-Hai region[D]. Rizhao: Qufu Normal University,2019.
[2]田仕文, 周长海. 废弃滨海盐田绿化改造工程研究与应用[J].海岸工程, 2013, 32(3): 53-58.
Tian S W, Zhou C H. Research and applications on the reformengineering of abandoned coastal saline soil in greening project[J]. Coastal Engineering, 2013, 32(3):53-58.
[3]Huang R D. Research progress on plant tolerance to soil salinityand alkalinity in sorghum[J]. Journal of Integrative Agriculture,2018, 17:739-746.
[4]张华文, 王润丰, 徐梦平, 等. 根际盐分差异性分布对高粱幼苗生长发育的影响[J]. 中国农业科学, 2019, 52(22): 4110-4118.
Zhang H W, Wang R F, Xu M P, et al. Effects ofheterogeneous salinity across rhizosphere on the growth ofsorghum seedlings[J]. Scientia Agricultura Sinica, 2019, 52(22):4110-4118.
[5]张瑞栋, 肖梦颖, 徐晓雪, 等. 高粱种子对萌发温度的响应分析与耐低温萌发能力鉴定[J]. 作物学报, 2020, 46(6):907-919.
Zhang R D, Xiao M Y, Xu X X, et al. Responses of sorghumhybrids to germination temperatures and identification of lowtemperature resistance[J]. Acta Agronomica Sinica, 2020, 46(6):907-919.
[6]高雪, 朱林, 张会丽. 盐胁迫对甜高粱和青贮玉米不同器官K+, Na+含量的影响[J]. 河南农业科学, 2017, 46(12): 29-35.
Gao X, Zhu L, Zhang H L. Effects of soil saline stress onK+ and Na+ content of different organs of sweet sorghum andsilage corn[J]. Journal of Henan Agricultural Sciences, 2017,46(12):29-35.
[7]赵德强, 李彤, 侯玉婷, 等. 玉米大豆间作模式下干物质积累和产量的边际效应及其系统效益[J]. 中国农业科学, 2020, 53(10): 1971-1985.
Zhao D Q, Li T, Hou Y T, et al. Benefits and marginal effectof dry matter accumulation and yield in maize and soybeanintercropping patterns[J]. Scientia Agricultura Sinica, 2020, 53(10):1971-1985.
[8]代会会. 豆科间作和地表覆盖对作物生长和土壤养分的影响研究[D]. 上海:上海大学, 2015.
Dai H H. Effects of intercropping with leguminous crops andsurface mulching on the growth of crops and soil nutrients[D].Shanghai: Shanghai University.
[9]刘振洋, 吴鑫雨, 汤利, 等. 小麦蚕豆间作体系氮素吸收累积动态及其种间氮素竞争关系[J]. 植物营养与肥料学报, 2020,26(7): 1284-1294.
Liu Z Y, Wu X Y, Tang L, et al. Dynamics of N acquisitionand accumulation and its interspecific N competition in a wheatfababean intercropping system[J]. Journal of Plant Nutritionand Fertilizer, 2020, 26(7):1284-1294.
[10]杨峰, 黄山, 崔亮, 等. 玉米/大豆套作下作物叶片氮、磷动态特征及其相关性分析[J]. 植物营养与肥料学报, 2013, 19(4): 781-789.
Yang F, Huang S, Cui L, et al. Dynamic changes andcorrelations of P and N concentrations in crop leaves underrelay intercropping system of maize and soybean[J]. Journal ofPlant Nutrition and Fertilizer, 2013, 19(4):781-789.
[11]聂胜委, 陈源泉, 隋鹏, 等. 玉米与不同植物间作对田间氨挥发的影响[J]. 中国农业科学, 2011, 44(3): 634-640.
Nie S W, Chen Y Q, Sui P, et al. Ammonia volatilization inintercropping field of maize with different crops[J]. ScientiaAgricultura Sinica, 2011, 44(3):634-640.
[12]张亦涛, 任天志, 刘宏斌, 等. 玉米追氮对玉米∥大豆间作体系产量和土壤硝态氮的影响及其后茬效应[J]. 植物营养与肥料学报, 2016, 22(1): 104-110.
Zhang Y T, Ren T Z, Liu H B, et al. Effect of topdressingnitrogen of intercropped maize strip on intercropped crop yieldsand soil nitrate nitrogen as well as its residual effect[J]. Journalof Plant Nutrition and Fertilizer, 2016, 22(1): 104-110.
[13]Fan F L, Zhang F S, Song Y N, et al. Nitrogen fixation offaba bean (Vicia faba L.) interacting with a non-legume in twocontrasting intercropping systems[J]. Plant and Soil, 2006,283(1-2): 275-286.
[14]朱亚琼, 简大为, 郑伟, 等. 不同种植模式下豆科绿肥对土壤改良效果的影响[J]. 草业科学, 2020, 37(5): 889-900.
Zhu Y Q, Jian D W, Zheng W, et al. Effects of improving soilfertility by planting different leguminous greenmanure plantsunder different mixed cropping patterns [J]. PrataculturalScience, 2020, 37(5):889-900.
[15]张久东, 包兴国, 曹卫东, 等. 河西灌区小麦与豆科作物间作和复种模式研究[J]. 核农学报, 2015, 29(4): 786-791.
Zhang J D, Bao X G, Cao W D, et al. Study on plantingpatterns about wheat inter-cropping and multiple cropping ofleguminous crops in Gansu hexi oasis irrigation area [J].Journal of Nuclear Agricultural Sciences, 2015, 29(4) :786-791.
[16]柏文恋, 张梦瑶, 刘振洋, 等. 小麦与蚕豆间作体系根系形态与磷吸收的定量解析[J]. 应用生态学报, 2021, 32(4): 1317-1326.
Bai W L, Zhang M Y, Liu Z Y, et al. Quantitative analysis ofroot morphology and phosphorus absorption in wheat and fababean intercropping system[J]. Chinese Journal of AppliedEcology, 2021, 32(4): 1317-1326.
[17]柏文恋, 张梦瑶, 任家兵, 等. 小麦/蚕豆间作作物生长曲线的模拟及种间互作分析[J]. 应用生态学报, 2018, 29(12):4037-4046.
Bai W L, Zhang M Y, Ren J B, et al. Simulation of crop growcurve and analysis of interspecific interaction in wheat and fatabean intercropping system[J]. Chinese Journal of AppliedEcology, 2018, 29(12):4037-4046.
[18]杨亚东, 冯晓敏, 胡跃高, 等. 豆科作物间作燕麦对土壤固氮微生物丰度和群落结构的影响[J]. 应用生态学报, 2017, 28(3): 957-965.
Yang Y D, Feng X M, Hu Y G, et al. Effects of legume-oatintercropping on abundance and community structure of soil Nfixingbacteria[J]. Chinese Journal of Applied Ecology, 2017,28(3):957-965.
[19]张小明, 来兴发, 杨宪龙, 等. 施氮和燕麦/箭筈豌豆间作比例对系统干物质量和氮素利用的影响[J]. 植物营养与肥料学报, 2018, 24(2): 489-498.
Zhang X M, Lai X F, Yang X L, et al. Effects of nitrogenapplication and intercropping ratio on dry matter production andnitrogen use efficiency of the oat and common vetchintercropping system [J]. Journal of Plant Nutrition andFertilizers, 2018, 24(2):489-498.
[20]冯晓敏, 杨永, 臧华栋, 等. 燕麦花生间作系统作物氮素累积与转移规律[J]. 植物营养与肥料学报, 2018, 24(3):617-624.
Feng X M, Yang Y, Zang H D, et al. Characteristics of cropnitrogen accumulation and nitrogen transfer in oat and peanutintercropping system [J]. Journal of Plant Nutrition andFertilizers, 2018, 24(3): 617-624.
[21]张爱加, 周明明, 林文雄. 不同种植模式对甘蔗根际土壤生物学特性的影响[J]. 植物营养与肥料学报 2013, 19(6):1525-1532.
Zhang A J, Zhou M M, Lin W X. Effects of differentcultivation patterns on microorganism of soil rhizosphere insugarcane fields[J]. Journal of Plant Nutrition and Fertilizers,2013, 19(6):1525-1532.
[22]王劲松, 樊芳芳, 郭珺, 等. 不同作物轮作对连作高粱生长及其根际土壤环境的影响[J]. 应用生态学报, 2016, 27(7):2283-2291.
Wang J S, Fan F F, Guo J, et al. Effects of different croprotations on growth of continuous cropping sorghum and itsrhizosphere soil micro-environment[J]. Chinese Journal ofApplied Ecology, 2016, 27(7): 2283-2291.
[23]邹剑秋. 高粱育种与栽培技术研究新进展[J]. 中国农业科学, 2020, 53(14): 2769-3773.
Zou J Q. New research progress on sorghum breeding andcultivation techniques[J]. Scientia Agricultura Sinica, 2020,53(14): 2769-3773.
[24]王海莲, 王润丰, 刘宾, 等. 不同生长时期收获对甜高粱农艺性状及营养品质的影响[J]. 中国农业科学, 2020, 53(14):2804-2813.
Wang H L, Wang R F, Liu B, et al. Effects of harvesting atdifferent growth stage on agronomic and nutritional qualityrelated traits of sweet sorghum[J]. Scientia Agricultura Sinica,2020, 53(14): 2804-2813.
[25]王劲松, 董二伟, 焦晓燕, 等. 不同种植模式对高粱晋糯3 号产量和养分吸收的影响[J]. 作物杂志, 2019(5): 166-172.
Wang J S, Dong E W, Jiao X Y, et al. Effects of differentplanting patterns on yield and nutrient absorption of sorghumjinnuo 3[J]. Crops, 2019(5):166-172.
[26]樊芳芳, 王劲松, 董二伟, 等. 连作对高粱生长及根区土壤环境的影响[J]. 中国土壤与肥料, 2016(3): 127-133.
Fan F F, Wang J S, Dong E W, et al. Effects of sorghumcontinuous cropping on the growth of sorghum and soilenvironment[J]. Soil and Fertilizer Sciences in China, 2016(3): 127-133.
[27]李光, 白文斌, 任爱霞. 高粱不同连作年限对其根系分泌物组成和化感物质含量的影响[J]. 生态学杂志, 2017, 36(12):3535-3544.
Li G, Bai W B, Ren A X. Effects of continuous croppingduration of sorghum on components of root exudates andcontents of allelochemicals[J]. Chinese Journal of Ecology,2017, 36(12): 3535-3544.
[28]樊芳芳, 白文斌, 王磊, 等. 不同调控措施对连作高粱生长及根际土壤微生物群落结构的影响[J]. 河南农业科学, 2020,49(10): 70-77.
Fan F F, Bai W B, Wang L, et al. Effects of different controlmeasures on the growth and rhizosphere soil microbialcommunity structure of continuous cropping sorghum [J].Journal of Henan Agricultural Sciences, 2020, 49(10): 70-77.
[29]王磊, 樊芳芳, 白文斌, 等. 不同连作年限高粱生长及根际土壤微生物群落功能多样性差异[J]. 山西农业科学, 2020, 48(9): 1456-1460.
Wang L, Fan F F, Bai W B, et al. Differences of sorghumgrowth and functional diversity of rhizosphere soil microbialcommunity of different continuous cropping years.[J]. Journalof Shanxi Agricultural Sciences, 2020, 48(9):1456-1460.
[30]周泉, 王龙昌, 邢毅, 等. 间作紫云英下油菜根际土壤微生物群落功能特征[J]. 应用生态学报, 2018, 29(3): 909-914.
Zhou Q, Wang L C, Xing Y, et al. Effects of intercroppingChinese milk vetch on functional characteristics of soilmicrobial community in rape rhizosphere[J]. Chinese Journalof Applied Ecology, 2018, 29(3): 909-914.
[31]朱奕豪, 刘晓丽, 陈为峰, 等. 黄河三角洲废弃盐田复垦土壤碳氮磷生态化学计量学特征[J]. 水土保持学报, 2020, 34(6): 352-360.
Zhu Y H, Liu X L, Chen W F, et al. Eco-stoichiometriccharacteristics of soil carbon, nitrogen, and phosphorus inreclaimed area of abandoned salt pan in the Yellow River Delta[J]. Journal of Soil and Water Conservation, 2020, 34(6):352-360.
[32]朱奕豪, 陈为峰, 宋希亮, 等. 黄河三角洲废弃盐田复垦土壤质量动态演变分析[J]. 应用基础与工程科学学报, 2021, 29(3): 545-561.
Zhu Y H, Chen W F, Song X L, et al. Reclaimed soil qualitydynamic evolution of abandoned salt field in the Yellow RiverDelta[J]. Journal of Basic Science and Engineering, 2021, 29(3): 545-561.
[33]朱奕豪, 李青梅, 刘晓丽, 等. 不同土地整治类型新增耕地土壤微生物群落特征研究[J]. 生态环境学报, 2022, 31(5):909-917.
Zhu Y H, Li Q M, Liu X L, et al. Characteristics of soilmicrobial community in newly cultivated land under differentland consolidation types [J]. Ecology and EnvironmentalSciences, 2022, 31(5): 909-917.
[34]郭童鑫, 姚晓华, 吴昆仑, 等. 青稞和豌豆根系形态和养分效率对种植模式和施肥水平的响应[J]. 植物营养与肥料学报,2023, 29(6): 1048-1059.
Guo T X, Yao X H, Wu K L, et al. Root morphology andnutrient efficiency of Tibetan barley and peas in response toplanting patterns and fertilization levels[J]. Journal of PlantNutrition and Fertilizers, 2023, 29(6): 1048-1059.
[35]冯晓敏, 高翔, 臧华栋, 等. 燕麦-绿豆间作效应及氮素转移特性[J].植物学报, 2023, 58( 1): 122-131.
Feng X M, Gao X, Zang H D, et al. Intercropping effect andnitrogen transfer characteristics of oat-mungbean intercrop[J].Chinese Bulletin of Botany, 2023, 58( 1): 122-131.
[36]陈晓芬, 陈静蕊, 秦文婧, 等. 红壤荒地上7 种豆科绿肥养分富集能力综合评价[J]. 中国草地学报, 2020, 42(4):153-160.
Chen X F, Chen J R, Qin W J, et al. Comprehensiveassessment of nutrient enrichment capacity of 7 leguminousgreen manures on red soil wasteland[J]. Chinese Journal ofGrassland, 2020, 42(4): 153-160.
[37]Liu Y C, Qin X M, Xiao J X, et al. Intercropping influencescomponent and content change of flavonoids in root exudatesand nodulation of fata bean[J]. Journal of Plant Interactions,2017, 12(1):187-192.
[38]刘战东, 张凯, 米兆荣, 等. 豫北潮土灌区土壤肥力特征与作物产量的关系研究[J]. 灌溉排水学报, 2019, 38(8): 31-37.
Liu Z D, Zhang K, Mi Z R, et al. Characteristics of soilfertility and its relation with crop yield in fluvo-aquic soilirrigation area of north Henan[J]. Journal of Irrigation andDrainage, 2019, 38(8):31-37.
[39]张月萌, 王倩姿, 孙志梅, 等. 间作豆科作物对山药田土壤化学和生物学性质的影响[J]. 应用生态学报, 2018, 29(12):4071-4079.
Zhang Y M, Wang Q Z, Sun Z M, et al. Effects of yam/leguminous crops intercropping on soil chemical and biologicalproperties of yam field [J]. Chinese Journal of AppliedEcology, 2018, 29(12):4071-4079.
[40]刘晓燕, 梁强, 庞天, 等. 甘蔗套种马铃薯机械化栽培对土壤微生物多样性及甘蔗养分吸收的影响[J]. 南方农业学报,2021, 52(2): 297-306.
Liu X Y, Liang Q, Pang T, et al. Influence of mechanizedcultivation on sugarcane/potato intercropping in soil microbialdiversity and nutrient absorption of sugarcane[J]. Journal ofSouthern Agriculture, 2021, 52(2): 297-306.
[41]赵雅娇. 紫花苜蓿/禾本科牧草间作优势及其氮高效机理和土壤微生态效应研究[D]. 兰州:甘肃农业大学, 2020.
Zhao Y J. Study on advantage of alfalfa/gramineae forageintercropping and mechanism of nitrogen efficiency and effect ofsoil microecological [D]. Lanzhou: Gansu AgriculturalUniversity, 2020.
[42]He Y, Ding N, Shi J C, et al. Profiling of microbial PLFAs:Implications for interspecific interactions due to intercroppingwhich increase phosphorus uptake in phosphorus limited acidicsoils[J]. Biology and Biochemistry, 2013, 57(12): 625-634.
[43]逄好胜, 胡举伟, 李鑫, 等. 小麦-苜蓿间作对土壤微生物群落功能多样性的影响[J]. 贵州农业科学, 2015, 43(8):160-164.
Pang H S, Hu J W, Li X, et al. Effects of wheat and alfalfaintercropping rhizospheric microbial on functional diversity ofsoil community[J]. Guizhou Agricultural Sciences, 2015, 43(8):160-164.
[44]胡国彬, 董坤, 董艳, 等. 间作缓解蚕豆连作障碍的根际微生态效应[J]. 生态学报, 2016, 36(4): 1010-1020.
Hu G B, Dong K, Dong Y, et al. Effects of cultivars andintercropping on the rhizosphere microenvironment foralleviating the impact of continuous cropping of faba bean[J].Acta Ecologica Sinica, 2016, 36(4):1010-1020.
[45]Li Q S, Wu L K, Chen J, et al. Biochemical and microbialproperties of rhizospheres under maize/peanut intercropping[J]. Journal of Integrative Agriculture, 2016, 15(1): 101-110.
[46]牛世全, 景彩虹, 廖世齐, 等. 河西走廊盐碱土细菌种群结构多样性的研究[J]. 西北师范大学学报(自然科学版), 2013,49(2): 90-95.
Niu S Q, Jing C H, Liao S Q, et al. Bacterial populationstructure diversity in saline-alkali soil in Hexi corridor[J].Journal of Northwest Normal University(Natural Science),2013, 49(2): 90-95.
(编辑:韩志强)
猜你喜欢
河北果树(2022年1期)2022-02-16 00:41:30
科学(2020年2期)2020-08-24 07:56:52
中国化肥信息(2018年12期)2018-03-01 03:13:00
中国中药杂志(2016年24期)2017-04-18 17:42:52
中国中药杂志(2017年4期)2017-03-28 17:06:08
中国中药杂志(2017年3期)2017-03-20 21:11:11
科技创新导报(2016年28期)2017-03-14 11:32:29
农民致富之友(2016年22期)2017-02-07 03:17:13
农民致富之友(2016年22期)2017-02-07 03:09:44
农民致富之友(2016年22期)2017-02-07 02:29:24
