
翅果油树重要害虫榆兴透翅蛾成虫触角的超微结构1)
2024-05-11 09:36:04余涛郑御丰杨兆富董艳玲
东北林业大学学报 2024年5期
余涛 郑御丰 杨兆富 董艳玲
(植保资源与病虫害治理教育部重点实验室(西北农林科技大学),杨凌,712100)
榆兴透翅蛾(Synanthedonulmicola)是一种重要的鳞翅目经济害虫,隶属于透翅蛾科(Sesiidae)透翅蛾亚科(Sesiinae),近年来对翅果油树(Elaeagnusmollis)造成严重危害。该种透翅蛾害虫一年发生1~2代,成虫主要以花蜜为食,而幼虫具有典型的钻蛀危害特性,主要蛀食翅果油树树干基部,危害初期,可见油渍状湿斑和黄褐色胶液,随虫龄增长其危害加重,严重时甚至可将地下根韧皮部成圈蛀空,导致翅果油树地上部分死亡,直接影响翅果油树产量和品质,造成严重的经济损失[1]。
触角是昆虫感知外界环境信息的重要感觉器官,对昆虫取食、躲避天敌、繁衍等适应性行为具有重要作用[2]。由于受到的环境选择压力不同,不同物种的触角上,通常着生有不同类型的感器,种间的感器变异对理解物种适应性演化也具有重要意义[3]。鉴于此,深入研究触角感器的功能,有助于揭示触角感器与昆虫生活习性的关系,达到对害虫有效防治的目的。因此,对重要害虫触角感器的类型、分布等研究是防治工作中不可或缺的重要一环。
目前,对鳞翅目昆虫触角感器的研究已有大量报道,其中对成虫触角感器的研究较多[4-9]。徐伟等[5]对杨背麦蛾(Anacampsispopulella)成虫触角感器的扫描电镜观察发现,雌、雄蛾均有8种感器,分别为毛形感器、锥形感器、刺形感器、耳形感器、Böhm氏鬃毛、鳞形感器、栓锥形感器、腔锥形感器;田彩红等[9]对草地贪夜蛾(Spodopterafrugiperda)成虫触角感器进行扫描电镜观察发现11种感器,较杨背麦蛾触角感器多出3种感器,分别为芽胞感器、坛形感器、腔乳头状感器;狄贵秋等[6]对美国白蛾(Hyphantriacunea)成虫触角电镜观察中发现,舌形感器为雄蛾所特有。由此可见,成虫触角的感器类型与分布在种间甚至同种雌、雄间都可能存在一定差异,这些差异可为重要害虫的物种鉴定、有效防治提供必要的理论依据[10]。然而,目前对透翅蛾科昆虫触角超微结构的研究较少,针对重要害虫榆兴透翅蛾的相关研究迄今未见报道。
本研究利用扫描电子显微镜技术(SEM)对榆兴透翅蛾成虫触角超微结构进行研究,通过比较触角感器类型、形态、分布模式,旨在寻找榆兴透翅蛾雌、雄成虫的性二型差异,以期为揭示触角感器与昆虫生活习性的关系及进一步探讨触角感器的功能和害虫防治提供理论依据。
1 材料与方法
供试昆虫:本研究样本主要采自山西省临汾市乡宁县翅果油树产区(地理中心坐标为35°41′N、110°30′E),所有样本均制作成针插标本,一部分用于形态学鉴定,另一部分用于扫描电镜观察。所有研究材料及凭证标本均保存于西北农林科技大学昆虫博物馆。
试剂和仪器:本试验所用试剂包括不同体积分数(70%、80%、85%、90%、95%、100%)的乙醇溶液,卡诺固定液(V(无水乙醇)∶V(冰醋酸)=3∶1)。仪器为SMZ1500体视变焦显微镜(日本Nikon公司),S4800冷场发射扫描电镜、EM-DSC20离子溅射仪(日本Hitachi公司)。
试验方法:取榆兴透翅蛾雌、雄虫各3头,用手术刀、解剖针在解剖镜下,将触角与虫体分离,将其放入离心管(含有体积分数为70%的乙醇溶液)中,超声清洗2~3次,每次10~15 s,震荡后置于表面皿(盛有体积分数为70%的乙醇溶液)中,在显微镜下,用毛刷刷去触角表面鳞粉、杂质,然后依次在体积分数为80%、85%、90%、95%、100%的乙醇溶液中脱水10 min,脱水后将自然干燥后的样品用导电胶粘在样品台上,用离子溅射仪对其表面喷金镀膜3 min。喷金后,在电子扫描仪中于15 kV加速电压时进行观察、拍照。触角感器的描述及分类主要参照文献[11],并结合前人研究结果[12-13]进行鉴定和命名。
数据处理:采用ImageJ 1.8.0软件(http://rsb.info.nih.gov/ij/)测量触角感器的长度、基部宽度,并使用SPSS 22.0软件(美国IBM软件分析公司)中独立样本t检验分析雌、雄蛾间差异。
2 结果与分析
2.1 榆兴透翅蛾触角的形态特征
图1为榆兴透翅蛾成虫触角形态及部分感器分布。雌、雄蛾触角均呈栉齿状,由柄节、梗节、鞭节三部分构成。柄节近似柱状,其上面覆盖有大量鳞片,基部着生于凹陷的触角窝内;梗节较柄节稍短,基部具有呈簇状分布的Böhm氏鬃毛以及少量鳞形感器;鞭节细长,雌、雄蛾均由56~58个鞭小节组成,近端部膨大逐渐呈弯钩状,均覆有大量鳞片,鳞片下触角表面具有褶皱状隆起,周围分布各种感器,主要分布在鞭节腹面。
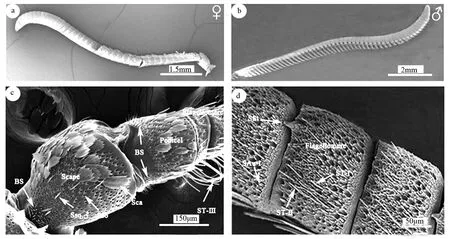
a为榆兴透翅蛾雌蛾触角整体图;b为榆兴透翅蛾雄蛾触角整体图;c为触角柄节、梗节;d为触角鞭小节腹面;BS为Böhm氏鬃毛;Ssq为鳞形感器;ST为毛形感器;SSt为栓锥形感器;SAu为耳形感器;Sca为鳞片;Sso为鳞片孔;Scape为柄节;Pedicel为梗节;Flagellomere为鞭节。图1 榆兴透翅蛾触角形态及部分感器分布
2.2 榆兴透翅蛾触角感器类型及特征
本研究在榆兴透翅蛾成虫触角上共发现8种感器类型,分别为Böhm氏鬃毛、毛形感器(ST-I、ST-II、ST-III)、鳞形感器、刺形感器、锥形感器、耳形感器I、腔锥形感器、栓锥形感器(图2)。其中Böhm氏鬃毛仅分布于柄节和梗节,其余7种感器类型主要分布于鞭节,但也有少量的鳞形感器分布于柄节和梗节。同时,研究还发现榆兴透翅蛾成虫触角上大部分感器的形态在雌、雄间无较大差异,但在感器长度与基宽等特征上具有一定差异(表1)。
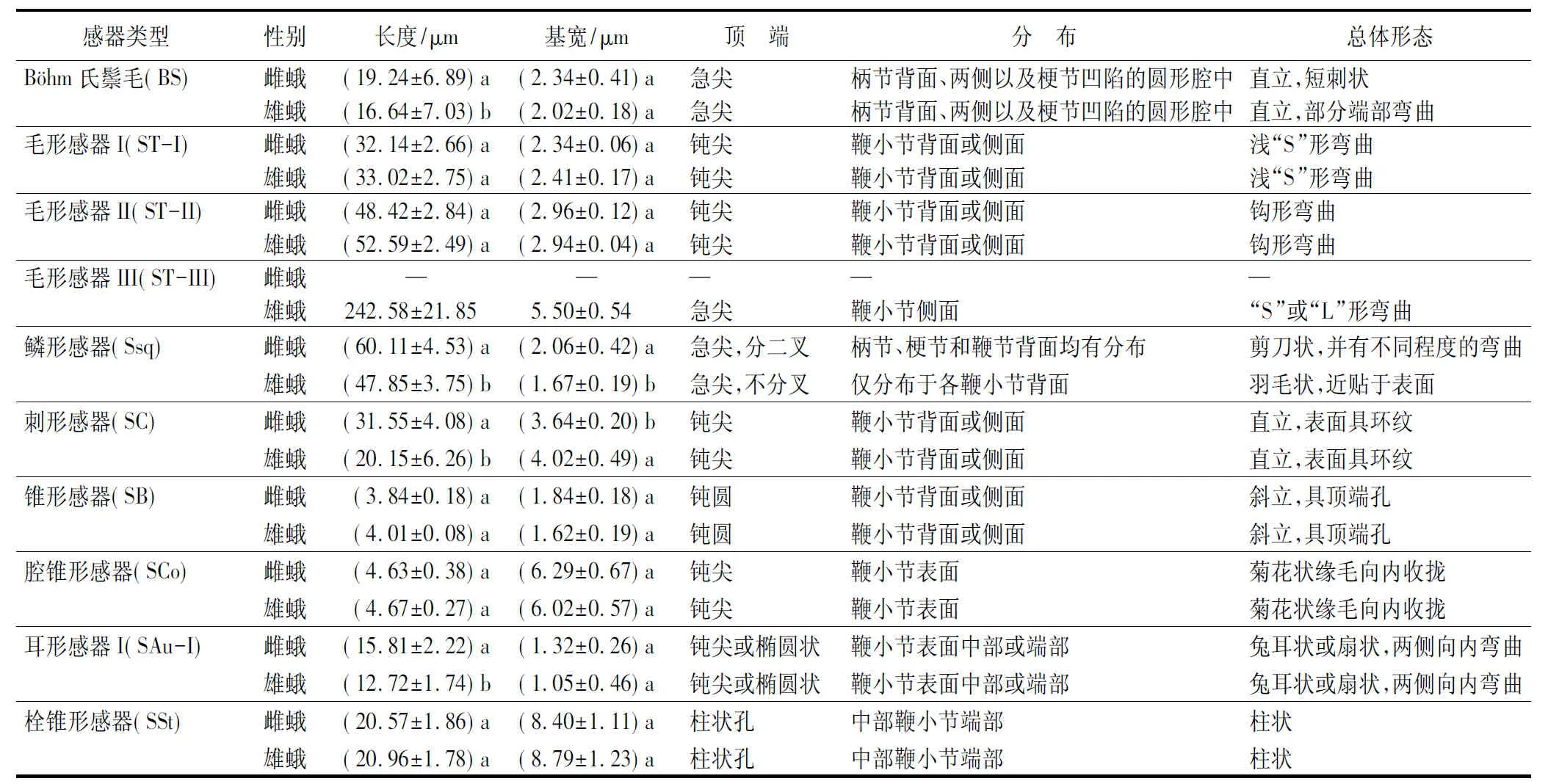
表1 榆兴透翅蛾触角不同类型感器的形态特征
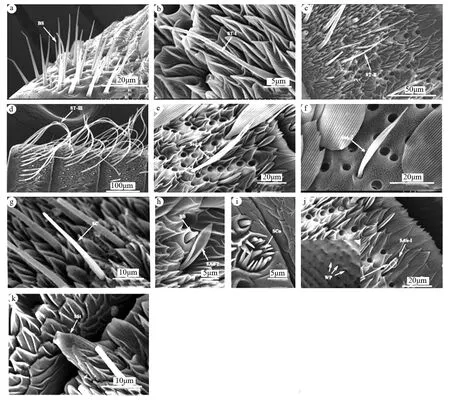
a为柄节或梗节近基部侧面Böhm氏鬃毛;b为毛形感器I;c为鞭小节腹面毛形感器II;d为鞭小节侧面毛形感器III;e为雌蛾触角上的鳞形感器;f为雄蛾触角上的鳞形感器;g为刺形感器;h为具顶端孔的锥形感器;i为被外缘毛包围的腔锥形感器;j为兔耳状的耳形感器I;k为鞭小节近端部的栓锥形感器。BS为Böhm氏鬃毛;ST-I为毛形感器I;ST-II为毛形感器II;ST-III为毛形感器III。Ssq为鳞形感器;SC为刺形感器;SB为锥形感器;SCo为腔锥形感器;SAu-I为耳形感器I;SSt为栓锥形感器;WP为表面微孔。图2 榆兴透翅蛾触角感器形态及种类
2.2.1Böhm氏鬃毛(BS)
Böhm氏鬃毛短刺状,基部具不明显臼状窝,从基部到端部渐细,端部尖锐,表面较光滑,无环纹或纵纹,多分布于柄节和梗节的侧面(图2a)。雌、雄蛾的Böhm氏鬃毛的长度分别为(19.24±6.89)、(16.64±7.03)μm,存在显著性差异;基部宽度分别为(2.34±0.41)、(2.02±0.18)μm,无显著性差异(表1)。
2.2.2毛形感器(ST)
毛形感器是榆兴透翅蛾分布最广、数量最多的感器,其基部无基窝,深陷于触角腹面凹陷腔内,表面具斜环纹,端部钝圆,从基部到端部渐细,多分布于触角各鞭小节的侧面和腹面,根据长度和弯曲程度将其分为3个亚型,分别为毛形感器I、毛形感器II、毛形感器III。
毛形感器I长度最短,基部粗大,至端部逐渐尖细,顶端钝圆,表面具斜环纹,整体呈浅s形弯曲,主要位于各鞭小节的腹面,背面近端部也有分布(图2b)。雌、雄蛾的毛形感器I长度分别为(32.14±2.66)、(33.02±2.75)μm,基部宽度分别为(2.34±0.06)、(2.41±0.17)μm,均无显著性差异(表1)。
毛形感器II长度相对适中,整体呈钩形,表面具斜环纹,逐渐向腹面弯曲,与触角表面形成夹角,主要位于各鞭小节的腹面和侧面(图2c)。雌、雄蛾的毛形感器II长度分别为(48.42±2.84)、(52.59±2.49)μm,存在显著性差异,基部宽度分别为(2.96±0.12)、(2.94±0.04)μm,无显著性差异(表1)。
毛形感器III长度最长,大多呈不规则弯曲状,端部锐尖,整体较毛形感器I、II更尖细,表面具斜环纹,聚集分布于各鞭小节的侧面,最末端鞭小节无分布(图2d)。仅雄蛾触角上有毛形感器III,长度为(242.58±21.85)μm,基部宽度为(5.50±0.54)μm(表1)。
2.2.3鳞形感器(Ssq)
鳞形感器基部具臼状窝,端部急尖,中间宽两边窄,呈明显柳叶状,表面具8~10个纵纹,近贴于触角腹面。榆兴透翅蛾雌、雄虫鳞形感器的形态具有明显差异,雌蛾鳞形感器近端部二分叉,而雄蛾鳞形感器近端部不分叉,具有明显的性二型差异(图2e-f)。雌、雄蛾的鳞形感器长度分别为(60.11±4.53)、(47.85±3.75)μm,基部宽度分别为(2.06±0.42)、(1.67±0.19)μm,均存在显著性差异(表1)。
2.2.4刺形感器(SC)
刺形感器基部具有向上突起的臼状窝,端部钝尖,从基部到端部渐细,表面具环状螺旋纹且无孔,呈长刺状,不弯曲,分布于触角各鞭小节的腹面(图2g)。雌、雄蛾的刺形感器长度分别为(31.55±4.08)、(20.15±6.26)μm,基部宽度分别为(3.64±0.20)、(4.02±0.49)μm,均存在显著性差异(表1)。
2.2.5锥形感器(SB)
锥形感器基部着生于触角表面突起的空腔内,端部具有一个顶端孔,中下部表面光滑,上部具有较浅的纵纹。数量较少,仅触角鞭节中部的部分鞭小节有2~3个(图2h)。雌、雄蛾的锥形感器长度分别为(3.84±0.18)、(4.01±0.08)μm,基部宽度分别为(1.84±0.18)、(1.62±0.19)μm,均无显著性差异(表1)。
2.2.6腔锥形感器(SCo)
腔锥形感器着生于表面凹陷的空腔内,中间有一柱状锥形突,锥形突表面具明显纵纹,周围缘毛向中间锥形突弯曲聚拢,具纵纹,分布于各鞭小节腹面(图2i)。雌、雄蛾中间感觉锥的长度分别为(4.63±0.38)、(4.67±0.27)μm,腔穴宽度分别为(6.29±0.67)、(6.02±0.57)μm,均无显著性差异(表1)。
2.2.7耳形感器(SAu)
耳形感器基部无臼状窝,外壁较厚,表面具有多条纵纹,纵纹内有均匀排列的小孔,主要分布于触角各鞭小节的腹面,数量众多,根据表面是否向内弯曲分为2个亚型,分别为耳形感器I、耳形感器II,但在榆兴透翅蛾触角上并未发现耳形感器II。
耳形感器I从基部开始,两边向内弯曲呈兔耳状,中部较宽,纵纹内小孔明显,主要分布于各鞭小节腹面(图2j)。雌、雄蛾耳形感器I的长度分别为(15.81±2.22)、(12.72±1.74)μm,存在显著差异,中间宽度分别为(2.49±0.29)、(1.98±0.32)μm,基部宽度分别为(1.32±0.26)、(1.05±0.46)μm,无显著性差异(表1)。
2.2.8栓锥形感器(SSt)
栓锥形感器呈柱状,基部从触角腹面各鞭小节近端部伸出,无臼状窝,中下部具较深的纵向沟壑,上部表面较光滑,并从四周向内凹陷形成空腔,每鞭小节端部均分布有1个(图2k)。雌、雄蛾栓锥形感器的长度分别为(20.57±1.86)、(20.96±1.78)μm,基部宽度分别为(8.40±1.11)、(8.79±1.23)μm,均无显著差异(表1)。
3 讨论
本研究发现榆兴透翅蛾雌、雄的成虫触角上共具有8种(10个亚型)感器,分别为Böhm氏鬃毛(BS)、毛形感器(ST-I、ST-Ⅱ、ST-III)、鳞形感器(Ssq)、刺形感器(SC)、锥形感器(SB)、耳形感器I(SAu-I)、腔锥形感器(SCo)、栓锥形感器(SSt)。毛形感器数量最多,分布范围最广,其次为耳形感器、栓锥形感器,而锥形感器数量最少(2~3个),其余感器数量不一。
在大多数昆虫的生命周期中,雄虫需要通过触角接受雌性释放的信息素并寻找雌虫进行交配,而雌虫主要负责释放信息素并繁殖后代,二者所承担的责任不同,这也在一定程度上导致了触角感器的雌雄二型现象,主要体现在雌、雄成虫触角感器的种类、形态以及数量、长度等方面,本研究也发现榆兴透翅蛾触角感器存在一定的性二型差异,这些超微结构的差异也进一步丰富了不同种昆虫乃至同种昆虫雌、雄之间的鉴别特征[14-16]。
Böhm氏鬃毛是一种能够感受重力刺激的感器[17]。榆兴透翅蛾的Böhm氏鬃毛与微红梢斑螟(Dioryctriarubella)[18]、玉带凤蝶(Papiliopolytes)[19]、豆卜馍夜蛾(Bomolochatristalis)[20]等大多数鳞翅目昆虫的Böhm氏鬃毛相似,推测其具有感受重力刺激并调节触角反应的功能。毛形感器作为昆虫触角上数量最多、分布范围最广的感器[21],在昆虫的生命活动中具有重要作用。本研究发现,榆兴透翅蛾雄虫触角上具有毛形感器III,而在雌蛾触角上缺失,且毛形感器III弯曲程度大,与触角表面形成不同的夹角,推测其具有识别信息素和感受环境刺激的功能[9,22-23]。榆兴透翅蛾的鳞形感器与大多数鳞翅目昆虫的鳞形感器相似[23-25],但雌、雄蛾触角上的鳞形感器具有性二型差异,这在其他鳞翅目昆虫中较为少见。据前人研究报道,同种昆虫触角感器的雌雄二型现象是由于识别信息素、寄主释放的化合物、生境中的外界物理刺激造成的[26-27],因此推测其具有感受外界机械运动并识别信息素的功能。刺形感器表面无孔、外壁较厚且长度较长,具有支撑保护和感受外界物理刺激的功能[2]。
锥形感器、耳形感器、腔锥形感器、栓锥形感器均具有嗅觉感器的功能,能够感受微量信息素、植物气味等化学信号。研究发现,榆兴透翅蛾的锥形感器、栓锥形感器端部具有一个顶端孔,耳形感器表面具有大量均匀排列的微孔,能够很好的接受化学信号。Pophof[28]在对家蚕(Bombyxmori)的触角感器进行嗅觉反应时,发现腔锥形感器对2-己烯醇、芳樟醇、其他萜类化合物的味道有反应,因此推测这四种感器均为嗅觉感器。
综上所述,本研究运用扫描电子显微镜技术初步探明了榆兴透翅蛾雌、雄成虫触角感器的类型和差异,并通过与其他鳞翅目昆虫触角感器的研究结果相比较,对榆兴透翅蛾成虫触角感器的功能进行了讨论,建议借助电生理实验等,进一步探究各种类型触角感器的具体功能,明晰不同感器性二型的内在机制。
猜你喜欢
热带农业科学(2023年8期)2023-09-25 07:57:00
中国棉花(2022年1期)2022-04-06 07:24:18
四川蚕业(2021年3期)2021-02-12 02:38:48
昆虫学报(2020年6期)2020-08-06 06:42:52
河南农业科学(2019年5期)2019-05-28 09:33:54
电子测试(2018年18期)2018-11-14 02:30:16
陕西理工大学学报(自然科学版)(2018年5期)2018-11-06 04:53:34
蚕学通讯(2017年3期)2017-03-23 08:00:02
河北林业科技(2016年5期)2016-11-08 03:12:48
山地农业生物学报(2015年2期)2015-12-02 04:27:40
