
东江三角洲河网浮游植物粒径特征研究
2024-05-07 05:55魏敬欣解常鑫杨婉玲刘乾甫曾艳艺
淡水渔业 2024年2期
魏敬欣,解常鑫,杨婉玲,刘乾甫,高 原,曾艳艺,王 超
(1.上海海洋大学海洋生态与环境学院,上海 201306;2.中国水产科学研究院珠江水产研究所,广州 510380;3.天津农学院,天津市水产生态及养殖重点实验室,天津 300384;4.农业农村部珠江流域渔业生态环境监测中心,广州 510380;5.广东省水产动物免疫与绿色养殖重点实验室,广州 510380)
浮游植物富含叶绿素、藻胆素等光合色素,可以利用光能进行光合作用产生有机物[1]。对浮游植物的粒径划分一般参照SIEBURTH等[2]的标准,可分为小型浮游植物(Micro-phytoplankton,简写为Micro-,20~200 μm)、微型浮游植物(Nano-phytoplankton,简写为Nano-,2~20 μm)和微微型浮游植物(Pico-phytoplankton,简写为Pico-,0.2~2 μm)。不同粒径的浮游植物由于其生长速率、对营养元素的摄入速率、沉降特性以及在食物网中下一步的能流去向等均各不相同,在生态系统食物链中也各自占据着重要地位,因此,开展浮游植物粒径结构的调查与研究对理解生态系统初级生产者在能量流动过程中发挥的作用具有重要意义[3]。目前,已有关于浮游植物粒径结构的研究多涉及海洋生态系统,我国南海[4]、辽河口[5]、黄茅海[6]等海域,国外的印度洋[7]、巴西Recife海港[8]、韩国牙山河口[9]等海域均有大量研究成果被报道。对海洋生态系统的现有研究结果均表明,在富营养海域,微型浮游植物所占百分比较高,能够达到50%左右,而在寡营养海域,微微型浮游植物所占百分比则较大。淡水生态系统中对浮游植物粒径结构的研究报道相对较少,王丽卿等[10]对淀山湖浮游植物粒径结构与环境因子进行调查,发现小型和微型浮游植物在淀山湖水体中贡献率较高。
东江三角洲位于广东省中部,与西、北江三角洲共同构成珠江三角洲。东江三角洲面积虽小,但其地理位置十分重要,东莞市、华南水运枢纽的黄埔新港及正在兴建的新沙港区均在其流域范围内,经济十分发达。但正因为工业化程度较高,企业、工厂数目众多,导致东江三角洲流域水环境污染严重,广东省水文局惠州分局2004年水质监测结果显示,东江三角洲河段水质类别在Ⅳ~Ⅴ类[11],水质状况较差。
浮游植物作为水生态系统的初级生产者,其群落结构特征已成为评估水环境状况的重要指标[12],相较于理化环境因子,浮游植物群落特征能更直观地反映调查时期内水环境的变化[13]。目前,对东江三角洲的研究大多集中于水动力方面[14],本研究于2021年至2022年春夏秋冬对东江三角洲河网水域中浮游植物粒径特征及其与环境因子的关系开展了季节性调查研究,旨在掌握该水域初级生产者的粒径结构特征,为大湾区典型三角洲河网水域的水环境保护和修复工作奠定相关研究基础。
1 研究地区与研究方法
1.1 采样地点与采样时间
为了尽可能展示东江三角洲水质的全面情况,在东江三角洲河网内共设置17个采样点(图1),分别为企石镇(QSZ)、石龙镇(SLZ)、台校西路(TXL)、工业路(GYL)、郭洲渡口(GZD)、航道局(HDJ)、东江大道(DJD)、宝华路(BHL)、金沙路(JSL)、东新路(DXL)、洪梅大桥(HMQ)、角尾水闸(JWS)、沿海三路(YHL)、滨江路(BJL)、沙田大道(STD)、轮渡路(LDL)和太平水道(TPS)。调查采样时间为2021年3、7、11月和2022年1月,每个季节的调查工作集中在5 d内完成。到达每个站位点后使用采水器采集水面下0.5 m处水样1 L作为测定水样。
图1 采样点站位图Fig.1 Sampling point station map
1.2 叶绿素的分级测定
将采集的测定水样先经200 μm孔径滤膜过滤,去除杂质,然后依次通过20 μm、2 μm和0.2 μm孔径滤膜,获得>20 μm、>2 μm和>0.2 μm粒径的叶绿素a浓度。抽滤后的滤膜剪碎分别放入离心管中,在离心管上标注采样时间、站位以及粒径,加入5 mL 90%丙酮溶液,震荡,于4 ℃冰箱中黑暗静置,24 h后取出,离心后(3 500 r/min;15 min),用紫外可见光分光光度计(UV-5200)测波长630 nm、664 nm、647 nm和750 nm处光密度值,分别记为OD750 nm、OD630 nm、OD647 nm和OD664 nm。叶绿素a浓度的计算公式为:
C(μg/L)=[11.85×(OD664 nm-OD750 nm)-1.54×(OD647 nm-OD750 nm)-0.08×(OD630 nm-OD750 nm)]×5÷1 000
总叶绿素a浓度为三个粒径叶绿素a浓度之和。
1.3 环境因子及测定方法

1.4 数据分析
采样站位图使用ArcMap 10.2软件绘制。环境因子、总叶绿素a浓度分布图以及浮游植物粒径结构图使用Origin 2023软件绘制。利用R软件(版本3.4.1)(http://www.r-project.org),以及“vegan”R包的RDA功能评估浮游植物各粒径叶绿素a浓度及所占百分比与环境因子之间的关系。使用IBM SPSS Statistics 27软件,以各站位浮游植物叶绿素a浓度所占百分比和环境因子为变量,绘制聚类分析树状图。
2 结果
2.1 环境因子特征
主要环境因子的时空特征如图2所示。季节差异显著的环境因子包括水温、pH、溶解氧、盐度、总磷和硅酸盐含量及水下光照度。其中,夏季的水温均值最高,为31.56 ℃,显著高于其他季节,春、秋、冬季水温均值接近,分别为21.38、20.31和19.45 ℃;pH值各月均值接近,冬季最高,为7.72,春季最低,为7.24;溶解氧秋季均值最高,为7.91 mg/L,夏季均值最低,为5.22 mg/L;盐度冬季的均值最高,为3.79 psu,夏季均值最低,为0.18 psu;总磷春季均值最高,为0.34 mg/L,夏、秋和冬季平均值接近,分别为0.22、0.22和0.20 mg/L;硅酸盐夏季均值最高,为11.73 mg/L,春秋季节接近,分别为8.14 mg/L与9.78 mg/L,冬季均值最低,为3.74 mg/L;光照夏季均值最高,为10 133 lx,秋季次之,为9 164 lx,春季最低,为3 551 lx;其他环境因子季节间差异不显著。
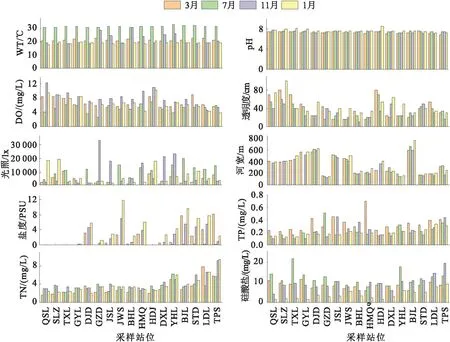
图2 环境因子特征Fig.2 Characteristics of environmental factors
空间特征差异显著的环境因子包括溶解氧、透明度、光照度、河宽、盐度、总磷和总氮,其中,溶解氧与透明度均呈现自上游至下游递减的趋势,靠近入海口的站位溶解氧与透明度均明显偏低;上游站位的河宽明显高于下游靠近入海口站位,差幅一般大于2倍;盐度的空间特征显示,几个盐度的最大值均出现在靠近入海口的站位;总磷、总氮浓度空间特征呈现出自上游至为下游递增的趋势,其中总磷在三角洲水域浓度较大,总氮在三角洲水域下方至入海口段浓度急剧上升。
2.2 叶绿素a总浓度的时空特征
叶绿素a浓度均值的季节特征显示,季节差异显著(图3)。其中夏季叶绿素a浓度均值最高,为21.09 μg/L,其次为春季的9.42 μg/L,秋季最低,为6.24 μg/L。粒径百分组成的季节特征主要表现为micro和nano的交替优势,但是相差不大。其中,夏季与冬季micro的占比略高于50%,分别为57%和51%;春季与秋季nano的占比略高于50%,分别为51%和55%。
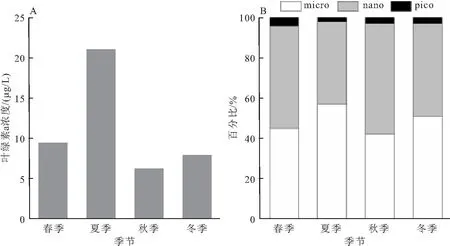
图3 叶绿素a总浓度的季节特征Fig.3 Seasonal characteristics of total chlorophyll-a concentrationA:各季节叶绿素a总浓度平均值;B:各季节叶绿素a粒径百分比
空间特征显示,春季叶绿素a总浓度在上游及河网水域站位呈现出递增趋势,在HDJ站位叶绿素a浓度达到最大值,在下游站位叶绿素a浓度明显降低,最小值出现在BJL站位。夏季各站位叶绿素a总浓度较大值均出现在靠上游的站位。秋冬季各站位叶绿素a总浓度普遍偏低,且各站位间叶绿素a总浓度较为接近。同时,我们发现各月份中叶绿素a总浓度最小值均出现在靠近入海口的站位(图4)。

图4 叶绿素a总浓度的空间特征Fig.4 Spatial characteristics of total chlorophyll-a concentration
2.3 叶绿素a粒径特征
浮游植物粒径百分组成如图5所示。整体来看,除个别季节的个别站位外,pico叶绿素a浓度的占比均很小,主要呈现为micro和nano的交替优势。季节特征显示,micro叶绿素a浓度占比季节差异显著,nano、pico叶绿素a浓度占比的季节差异不显著。
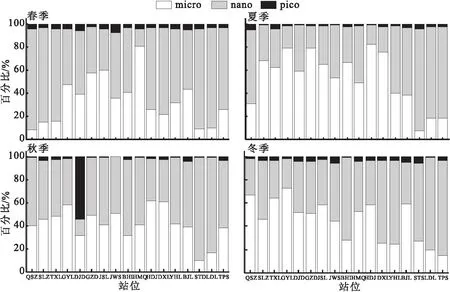
图5 浮游植物粒径百分比的空间分布Fig.5 Spatial distribution of phytoplankton particle size percentage
空间特征显示,春季小型浮游植物叶绿素a浓度在中部河网站位占比较高,在上游与下游靠近入海口站位占比较低,在GYL至HMQ站位间,小型浮游植物叶绿素a浓度占比均大于当月平均值,微型浮游植物叶绿素a浓度占比趋势与小型相反;夏季小型浮游植物叶绿素a浓度在大多数站位占比较高,只在下游STS、LDL和TPS站位占比较低,且上游与河网多数站位小型浮游植物叶绿素a浓度占比均大于当月平均值;秋季小型与微型浮游植物叶绿素a浓度占比相当,在多数站位二者占比均在50%左右,在下游STS、LDL站位,微型浮游植物叶绿素a浓度占比显著高于小型;冬季小型浮游植物叶绿素a浓度占比呈现自上游至下游递减趋势,在最靠近入海口的TPS站位占比最低,为14%,微型浮游植物叶绿素a浓度占比则呈现出递增趋势。
2.4 RDA分析
选择浮游植物各粒径浓度及总浓度与环境因子进行RDA分析,结果显示共筛选出6个关键影响因子,包括水温、河宽、盐度、光照度、硅酸盐和总氮。二维降序图如图6A所示。基于轴1的关联性结果显示,各粒径叶绿素a浓度及总浓度与水温均呈正相关关系,与盐度均呈负相关关系。基于轴2的关联性结果显示,小型浮游植物叶绿素a浓度与硅酸盐和总氮浓度呈负相关关系,微型浮游植物叶绿素a浓度及总浓度与光照、硅酸盐和总氮浓度呈正相关关系,与河宽呈负相关关系。
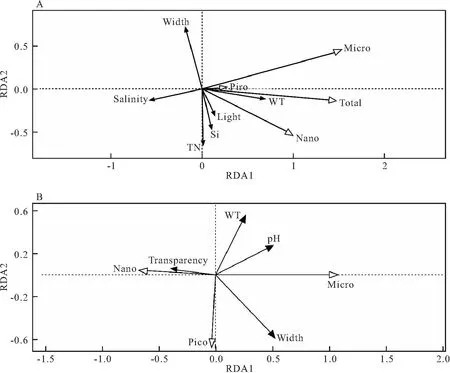
图6 浮游植物粒径特征与环境因子的RDA分析Fig.6 RDA analysis of phytoplankton particle size characteristics and environmental factorsA:各粒径浓度;B:各粒径浓度所占百分比
选择浮游植物各粒径浓度占比与环境因子进行RDA分析,结果显示共筛选出4个关键影响因子,包括水温、pH、河宽和透明度。二维降序图如图6B所示。基于轴1的关联性结果显示,小型浮游植物叶绿素a浓度占比与pH、河宽呈正相关关系,与透明度呈负相关关系,微型浮游植物叶绿素a浓度占比与之相反。基于轴2的关联性结果显示,微型浮游植物叶绿素a浓度占比与水温呈负相关关系。
2.5 调查站位的聚类分析
以各站位浮游植物叶绿素a粒径组成和环境因子为变量,对东江三角洲17个站位进行聚类分析(见图7)。结果显示,当连接距离在25左右时,所有的站位被分成了两大类。依据聚类分析结果和地理特征,可以将调查地区大致划分为两类,第一类包括QSZ、SLZ、TXL等10个站位,主要为上游站位,第二类包括DJD、HMQ、JWS等7个站位,主要为下游站位,经过分类后可以更清楚地看到浮游植物粒径组成与环境因子的关系(见表1)。我们发现,两个类群的叶绿素a粒径结构差异显著,其中两类群中Micro、Nano叶绿素a浓度占比差异显著,Pico叶绿素a浓度占比差异极显著,第一类中小型浮游植物叶绿素a浓度占比大于微型浮游植物叶绿素a浓度占比,第二类中恰好相反。另外,研究发现,两类群间差异显著的物理因子主要为盐度,差异显著的化学因子主要为pH和TP。结合RDA分析结果,发现盐度与pH出现在RDA分析出的关键影响因子中,推测盐度和pH是影响东江三角洲浮游植物粒径组成空间分布的关键环境因子。
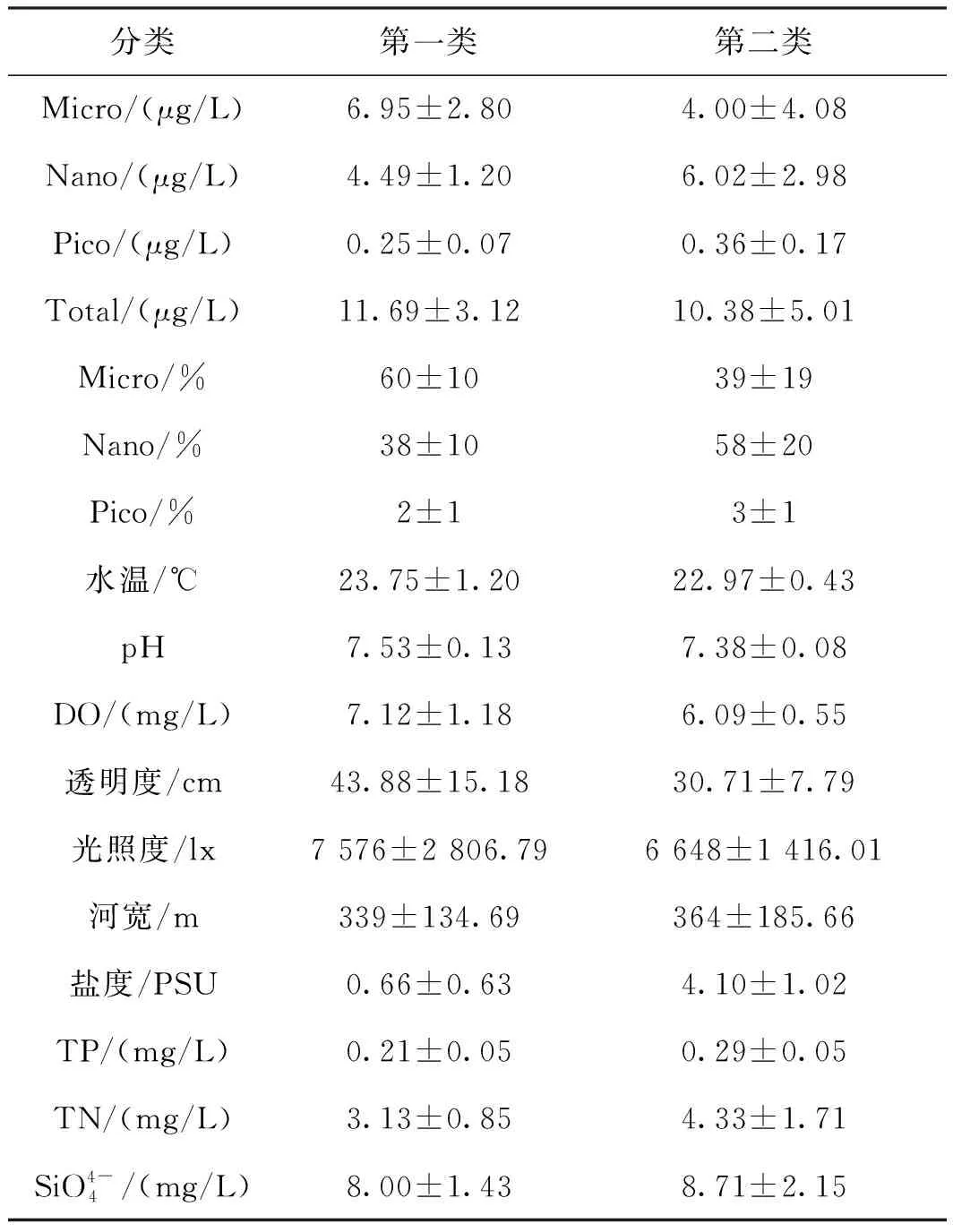
表1 两个类群叶绿素a粒径结构和环境因子的平均值Tab.1 Mean value of chlorophyll-a particle size structure and environmental factors of two taxa
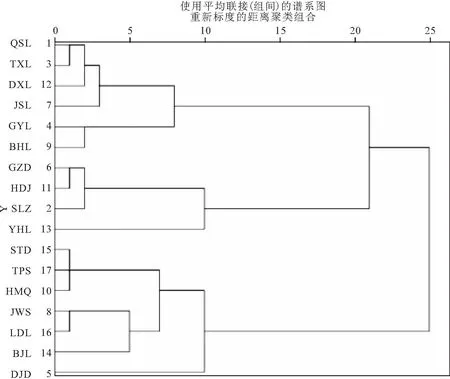
图7 聚类分析树状图Fig.7 Cluster analysis
3 讨论
3.1 叶绿素a总浓度的时空分布特征
研究水域叶绿素a总浓度的季节特征显示,最大值出现在夏季,分析原因:(1)夏季水温明显高于其他季节。研究表明,浮游植物体内酶系统控制着浮游植物的光合作用和营养运输,而温度直接影响酶系统的活性,因此,温度对浮游植物的光合作用速率以及生长速率均有重要影响。SHIMARAEVA等[15]对南贝加尔湖进行研究发现,叶绿素a浓度与水温呈正相关。国内学者研究表明,叶绿素a浓度与水温呈显著正相关关系,丰水期叶绿素a浓度显著高于枯水期与平水期[16]。RDA分析显示,水温与总叶绿素a浓度呈正相关关系,印证了以上观点。(2)夏季的高温伴随着较高的光照强度,光照强度是浮游植物生长的重要影响因素[17],其会影响浮游植物光合作用强度和放氧速率,进而影响浮游植物光合作用效率。在富营养水域,光照强度成为浮游植物生长的限制因子,且光照强度与浮游植物浓度常呈正相关关系[18],这与我们的结果一致。
研究水域叶绿素a总浓度的空间特征显示,靠近下游入海口的站位的叶绿素a总浓度明显低于其他站位。分析原因:(1)与河口型三角洲河网的特性有关。RDA分析结果显示,盐度与叶绿素a总浓度呈负相关关系。ANDRADE等[19]对格兰德河进行研究时发现,当河口盐度开始下降时,叶绿素a浓度增加,盐度与叶绿素浓度呈显著负相关,这与我们的研究结果一致。研究表明,大多数淡水浮游植物在高盐度中生长和生存能力下降。随着盐度的增加,生物会受到盐度的胁迫作用,细胞内外的渗透压会改变,此外,高盐度海水还会使细胞内积累ROS(reactive oxygen species),ROS会导致细胞结构的改变以及细胞膜的过氧化,这些因素都会抑制细胞的生长[20]。郑逸等[21]通过实验发现,当环境中盐度高于0.5%时,普通小球藻和蛋白核小球藻的生长均受到抑制作用,且抑制作用随盐度升高而增强。(2)与河口水域低透明度与低光照强度有关。在浑浊的河口环境中,由于浑浊造成的水下光衰减成为控制浮游植物生长的最重要的因素[22]。在阿根廷混合良好的巴伊亚布兰卡河口,水柱中悬浮颗粒物的光限制强烈影响了浮游植物的繁殖[23]。在荷兰浑浊的Westerschelde河口,浮游植物的产量主要由光驱动,而不是由营养物质驱动[24],以上研究结论均与我们的结果相一致。
3.2 粒径结构的时空分布特征
东江三角洲浮游植物粒径结构的季节分布特征显示,小型浮游植物叶绿素a浓度占比季节差异显著,分析原因,(1)与水环境的pH有关,RDA分析结果表明,小型浮游植物叶绿素a浓度占比与pH呈正相关关系。WANG等[25]对白洋淀水域进行研究时发现,小型浮游植物叶绿素a浓度占比与pH呈正相关,这与我们的研究结果一致。(2)与水体温度有关,RDA分析结果表明,小型浮游植物叶绿素a浓度占比与水温呈正相关关系。SATO等[26]的研究结果显示,浮游植物细胞大小随温度和营养盐浓度变化的多元线性回归结果表明,细胞大小与温度呈显著正相关关系,这与我们的结果一致。(3)与水环境中硅酸盐含量有关。葛大艳等[27]对珠三角河网进行研究时发现,颗粒沟链藻是该流域的绝对优势种,颗粒沟链藻属于小型浮游植物,而硅藻的生长与水体中硅酸盐的含量呈正相关关系。已有研究表明,硅藻细胞密度与水体中硅酸盐浓度呈正相关关系,较高的硅元素含量有助于硅藻种群的生长与竞争[28],这与我们的调查结果一致。
东江三角洲浮游植物粒径结构的空间分布特征显示,小型与微型浮游植物粒径浓度呈现交互优势,小型浮游植物叶绿素a浓度占比在上游站位占比较大,微型浮游植物叶绿素a浓度占比在自HMQ站位起始的下游站位占比较大,分析原因,(1)水温可以影响浮游植物粒径结构,RDA分析结果显示,水温与小型浮游植物叶绿素a浓度占比呈正相关,与微型浮游植物叶绿素a浓度占比呈负相关。杨红等[29]对象山港进行研究时发现,水温升高小型浮游植物的量也随之升高,而微型浮游植物的量随之下降,这与我们的结果相一致。(2)与河宽的变化有关,RDA分析结果显示,河宽与小型浮游植物叶绿素a浓度占比呈正相关,与微型浮游植物叶绿素a浓度占比呈负相关。河宽的增大增加了水体的径流量,有利于喜好流动水体的硅藻种类生长,抑制了喜好静止水体的蓝藻种类的生长,从而影响了浮游植物的粒径结构组成[30]。(3)与水体酸碱度的变化有关。pH能影响浮游植物的生长与分布,pH值过高或过低都不利于浮游植物的生长与繁殖[31]。藻类对不同pH有不同的适应种,例如,TEMPONERAS等[32]研究发现,蓝藻生物量与pH值显著正相关,马京久等[33]研究发现,汉江中下游水体中浮游植物优势种与pH呈正相关,而钱塘江着生的藻类群落结构与 pH 值呈负相关[34]。(4)我们推测,盐度是影响浮游植物粒径结构的关键环境因子。MITBAVKAR等[35]对印度河口进行研究时发现,盐度在浮游植物分布中起着重要作用,富含藻红蛋白的聚球藻有两个亚组,一组仅在盐水中发现,另一组存在于整个盐度梯度中,这证实了盐度对浮游植物群落组成的影响。LIU等[36]对鸭绿江进行研究时发现,微型浮游植物群落受到水体盐度的正向影响。许多研究表明,浮游植物群落组成结构也会受到盐度的影响,例如硅藻门、隐藻门种类更适宜生活在低盐度环境中,而甲藻门和蓝藻门种类在较高的盐度环境中也能生存[37]。葛大艳等[27]对珠三角河网进行研究时发现,硅藻是该水域的优势种,且多数硅藻种类属于小型粒径,这证实东江三角洲水域小型浮游植物也以硅藻类群为主,随着盐度的升高,硅藻类群的生长受到抑制,小型浮游植物叶绿素a浓度占比减小,从而导致微型浮游植物叶绿素a浓度占比增加。
猜你喜欢
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
人大建设(2019年5期)2019-10-08
西南石油大学学报(自然科学版)(2018年5期)2018-11-06
文学少年(有声彩绘)(2017年4期)2017-07-03
小布老虎(2016年14期)2016-12-01
广东海洋大学学报(2015年3期)2015-12-22
少儿科学周刊·少年版(2015年11期)2015-12-17
华南农业大学学报(2015年5期)2015-12-04
中国洗涤用品工业(2015年5期)2015-02-28
