
基于高密度SNP标记对112份温热改良玉米自交系的遗传特征鉴定与表型性状评价
2024-04-30 10:27:56王栋郭爽聂蕾何玥涂亮蒋喻林刘鹏飞王安贵祝云芳郭向阳吴迅陈泽辉
江苏农业科学 2024年6期
关键词:遗传多样性
王栋 郭爽 聂蕾 何玥 涂亮 蒋喻林 刘鹏飞 王安贵 祝云芳 郭向阳 吴迅 陈泽辉
摘要:种质资源表型和基因型鑒定评价是深入发掘优势基因资源、创制优势种质材料的前提和基础,对于培育优势玉米杂交种具有十分重要的意义。以112份温热改良玉米自交系为供试材料,利用包含6万个SNP标记的Maize SNP 50芯片对其进行基因型鉴定,并结合该群体在不同生态环境下的表型鉴定结果,对不同自交系的抗性、所属杂种优势类群等进行评价。基因型分析结果发现,112份自交系首先被划分为Iodent种质(Ⅰ类)、Suwan种质(Ⅱ类)和温热改良种质(Ⅲ类)3类,再次被划分为分别以HCL645、T32、QR273、黄早四、PH4CV和B73为代表自交系的6个亚群;进一步被划分为以HCL645、T32、QB446、QR273、黄早四、PH4CV、郑58、齐319和B73为代表自交系的9个亚群。结果显示,112份自交系的散粉期、株高、穗位高、穗长、穗粗、穗行数、行粒数、秃尖长、茎腐病、南方锈病和白斑病等不同性状之间呈现出一定的相关性。散粉期与株高、穗位高呈极显著正相关,与茎腐病、南方锈病和白斑病病级均呈极显著负相关,相关系数分别为0.19、0.32、-0.28、-0.32和-0.21。此外,Iodent种质表现出生育期短、穗行数多等优点;Suwan种质表现出生育期长、植株高大和抗病性强的特点;温热改良种质表现出生育期较短的优点。其中,Iodent种质和Suwan种质表现出优势互补,具有丰富的育种利用价值。
关键词:玉米自交系;遗传多样性;性状评价;SNP标记
中图分类号:S513.03 文献标志码:A
文章编号:1002-1302(2024)06-0077-07
收稿日期:2023-10-30
基金项目:贵州省科技支撑计划(编号:黔科合支撑[2022]重点029);贵州省山地农业关键核心技术攻关项目(编号:GZNYGJHX-2023012);国家自然科学基金(编号:32171981、32060460、32160451);贵州省农业科学院科技创新项目(编号:[2022]03号);贵州省育种科研基础平台创新能力建设项目(编号:黔科合服企[2022]014);黔农科院国基后补助项目(编号:[2021]16号、[2022]02号、[2022]09号);贵州省自然科学基金(编号:黔科合基础[2022]一般236);贵州喀斯特山区重要作物生物育种平台建设项目(编号:黔科合中引地[2023]033);贵州省十层次人才项目(编号:[2018]5629)。
作者简介:王栋(1997—),男,贵州剑河人,硕士研究生,研究方向为遗传育种。E-mail:wdong926400@163.com。
通信作者:陈泽辉,博士,研究员,主要从事玉米遗传育种研究。E-mail:chenzh907@sina.com。
种质资源是大自然长期演变形成的产物,携带着大量的有益基因,是品种选育和遗传改良的物质基础[1-3]。开展种质资源研究对农业发展、保护生物多样性和生态环境具有重要意义,是研究和培育高产、优质、抗病虫新品种的重要环节,更是提升一个国家作物育种领域核心竞争力的关键[4]。玉米种质资源具有丰富的遗传多样性,在长期的驯化、人工选择过程中,形成了丰富的表型和多样的基因型。基因鉴定和表型评价是种质资源研究、认知和改良利用的基础[5]。Liu等利用Maize SNP 50芯片对240份玉米自交系进行了基因鉴定,并将240份自交系划分为5个类群[6]。Wu等利用高通量的SNP芯片,将367份自交系划分为5个类群[7]。张亚菲等对黄淮海夏玉米区36个品系的农艺性状进行了聚类分析,将其划分为三大类6个亚类[8]。郭欢乐等对139份玉米地方种进行表型性状鉴定,将其划分为3个类群[9]。王新涛等对572份玉米种质资源进行粗缩病抗性鉴定,筛选出22份抗病材料,并利用40对简单重复序列(SSR)标记对22份抗病材料和6份测验种进行遗传多样性分析,将28份材料划分为7个类群,并揭示了来自美国杂交种78599的玉米自交系所组成亚群的粗缩病抗性较好[10]。王婷等基于69对SSR引物,将73份玉米种质资源划分为6个类群,并结合瘤黑粉病的抗性鉴定结果,揭示了高抗瘤黑粉病材料主要集中在旅大红骨和PB种质[11]。然而,种质资源研究仍存在资源收集保护力度不够、育种利用效率低、精准鉴定不足等问题[12-13],尤其是针对温热改良材料的研究需要进一步深入。
因此,本研究以112份重要玉米自交系为供试材料,利用包含6万个单核苷酸多态性(single nucleotide polymorphism,SNP)标记的Maize SNP 50芯片对其进行基因型鉴定,结合该群体的散粉期、株高、穗位高、穗长、穗粗、穗行数、行粒数、秃尖长,茎腐病、南方锈病和白斑病等表型鉴定结果,对不同自交系的抗性、所属杂种优势类群等进行评价,揭示不同自交系间的遗传差异,并剖析不同杂种优势类群间的表型性状特点,以期为后续优异基因挖掘、种质遗传改良和强优势组合选育提供理论参考。
1 材料与方法
1.1 供试材料
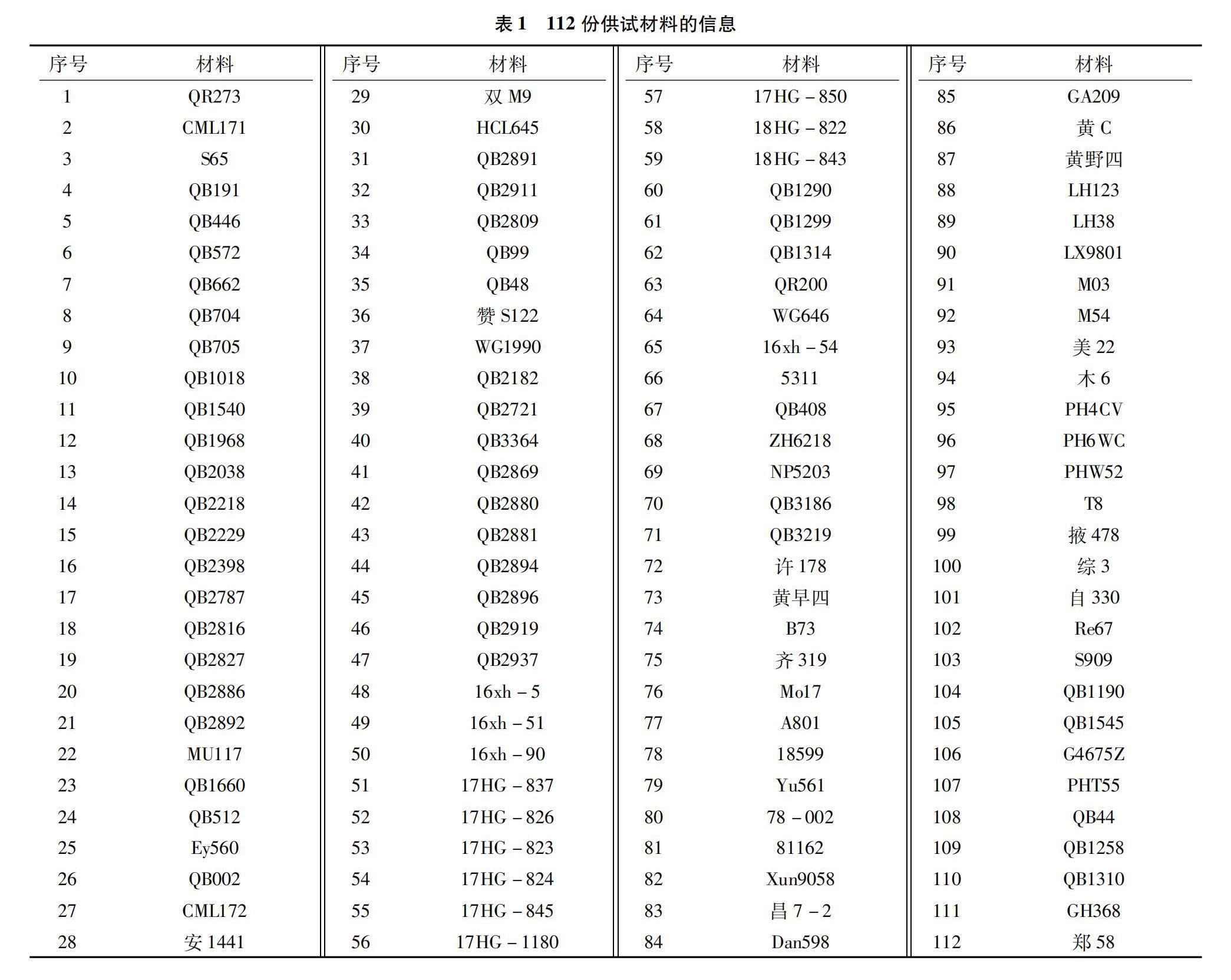
本研究所用材料为112份温热改良玉米自交系,详细信息见表1。
1.2 田间试验设计
2021年11月,将112份供试材料种植于海南省三亚市,行长3 m,行距6 cm,株距25 cm,双行区,自然散粉,田间管理参照当地生产水平进行。分别调查不同自交系的散粉期、株高、穗位高、穗长、穗粗、穗行数、行粒数、秃尖长、茎腐病和南方锈病发生情况等。
2022年6月,112份供试材料分别种于云南省的文山州、石林市和墨江市,进行白斑病抗性鉴定。
1.3 基因型鉴定
基因型鉴定工作已于2019年由笔者所在课题组完成[14]。于供试材料5叶期时取幼嫩叶片备用,采用改良十六烷基三甲基溴化铵(CTAB)法提取基因组DNA,并用琼脂糖凝胶电泳和分光光度计检验基因组DNA的完成性和浓度。检测合格的DNA样品送北京康普森生物技术有限公司进行后续杂交和鉴定试验。基因型鉴定采用包含6万个SNP标记的MaizeSNP50芯片,相关工作由北京康普森生物技术有限公司完成,后续的SNP读取、高质量位点筛选等参照Wu等描述的方法[14]进行。
1.4 表型测定
散粉期、株高、穗位高、穗长、穗粗、穗行数、行粒数和秃尖长等性状参考石云素的方法[15]进行评价与描述。玉米茎腐病和南方锈病参考NY/T 1248.5—2006 《玉米抗病虫性鉴定技术规范》进行评价。玉米白斑病参考专利CN115058486A 《一种玉米白斑病的抗病性鉴定方法》进行评价,本研究白斑病病级取3个环境综合病情值。
1.5 群体结构分析
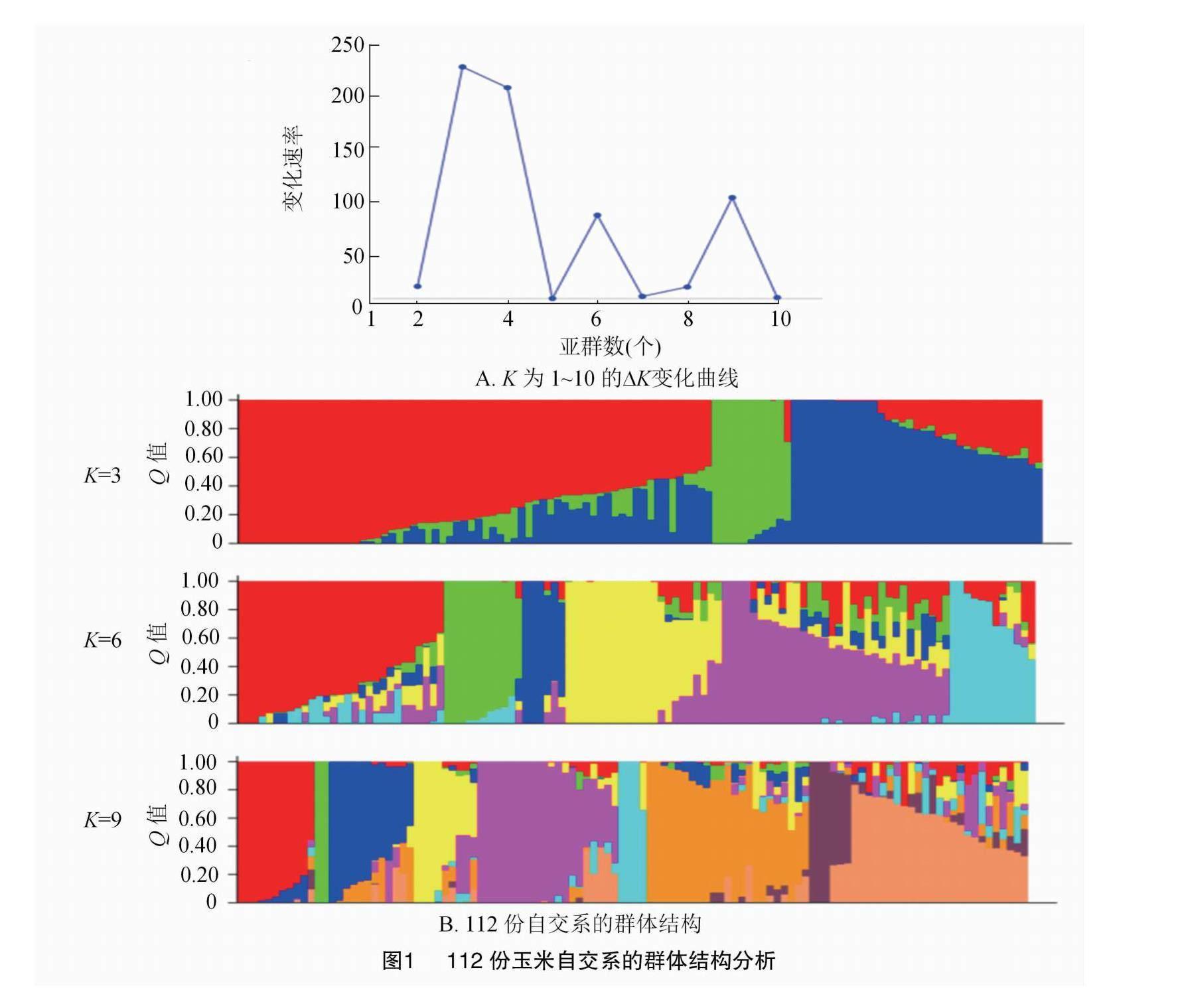
基于基因型鉴定结果,筛选出3 758个高质量SNPs,并利用Structure 2.3.4软件对112份自交系进行群体结构分析,设置亚群数量(K)为1~10个,迭代次数为3次,模拟参数迭代(length of bum-in period)和蒙特卡罗迭代(markov chain monte carlo,MCMC)均设置为5 000次;将计算所得结果输入Structure Harvester(https://taylor0.biology.ucla.edu/struct_harvest/),计算2个连续后验概率值的变化速率(ΔK)和每个材料在1~10亚群的Q值,以 Q= 0.5为分界线,最终确定亚群数量和结构。
2 结果与分析
2.1 群体结构分析
根据Structure 2.3.4软件对112份自交系进行群体结构分析结果(图1),发现K值取3、6和9时均有峰值。因此,112份自交系首先被划分为Iodent种质(Ⅰ类)、Suwan种质(Ⅱ类)和温热改良种质(Ⅲ类)3类;再次被划分为分别以HCL645、T32、QR273、黄早四、PH4CV和B73为代表自交系的6个亚群;进一步被划分为分别以HCL645、T32、QB446、QR273、黄早四、PH4CV、郑58、齐319和B73为代表自交系的9个亚群。
2.2 表型分析
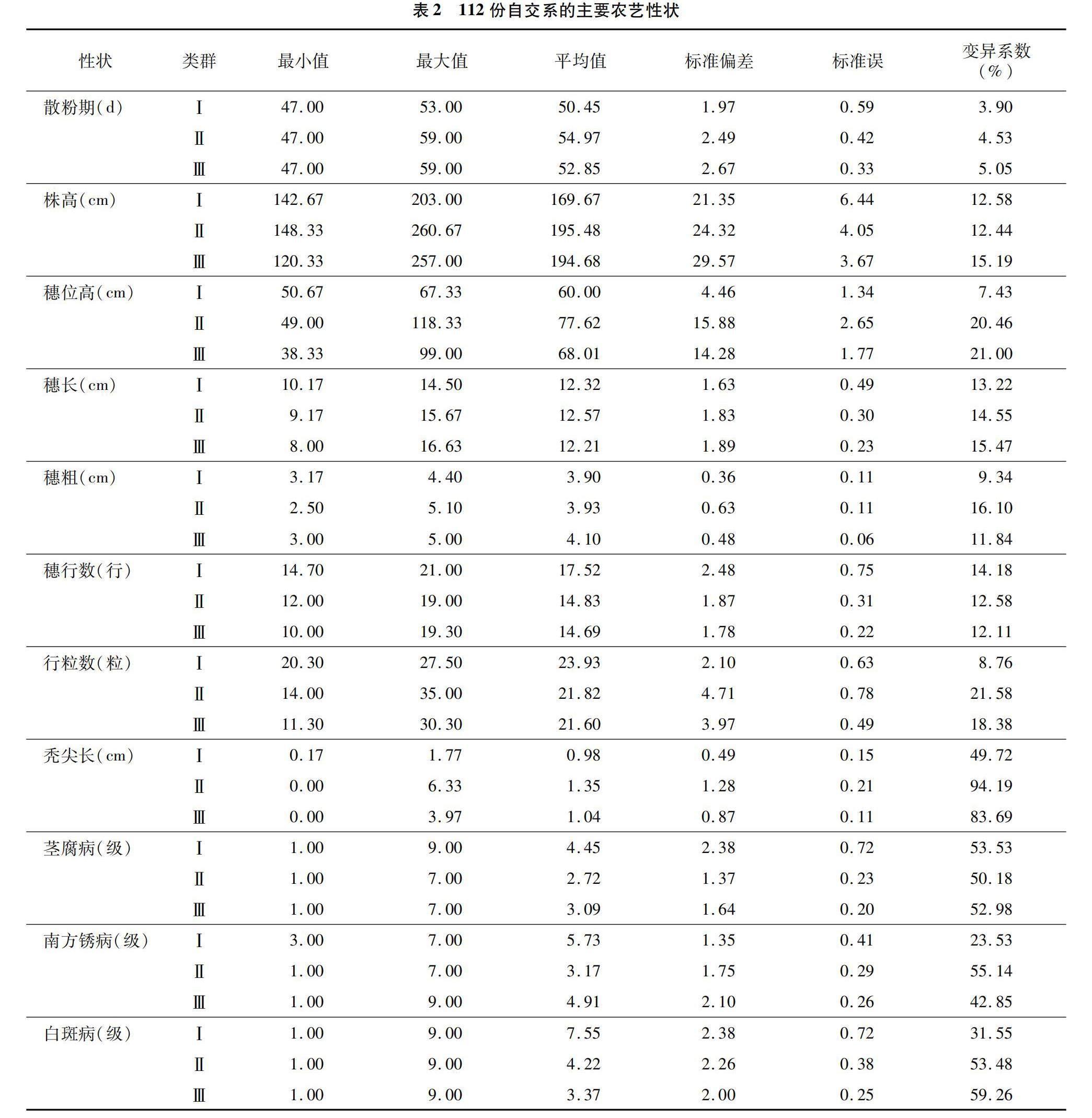
表型鉴定结果(表2)显示,对于散粉期(即从播种到散粉所需天数)性状,Ⅰ类群、Ⅱ类群和Ⅲ类群的变化范围分别为47.00~53.00、47.00~59.00、47.00~59.00 d,变异系数分别为3.90%、4.53%和5.05%。3个类群株高的变化范围分别为142.67~203.00、148.33~260.67、120.33~257.00 cm,变异系数分别为12.58%、12.44%和15.19%;穗位高的变化范围分别为50.67~67.33、49.00~118.33、38.33~99.00 cm,变异系数分别为7.43%、20.46%和21.00%;穗长的变化范围分别为10.17~14.50、9.17~15.67、8.00~16.63 cm,变异系数分别为13.22%、14.55%和15.47%;穗粗的变化范围分别为3.17~4.40、2.50~5.10、3.00~5.00 cm,变异系数分别为9.34%、16.10%和11.84%;穗行数的变化范围分别为14.70~21.00、12.00~19.00、10.00~19.30行,变异系数分别为14.18%、12.58%和12.11%;行粒数的变化范围分别为20.30~27.50、14.00~35.00、11.30~30.30粒,变异系数分别为8.76%、21.58%和18.38%;秃尖长的变化范围分别为0.17~1.77、0.00~6.33、0.00~3.97 cm,变异系数分别为49.72%、94.19%和83.69%;莖腐病的病级变化范围分别为1.00~9.00、1.00~7.00、1.00~7.00,变异系数分别为53.53%、50.18%和52.98%;南方锈病的病级变化范围分别为3.00~7.00、1.00~7.00和1.00~9.00,变异系数分别为23.53%、55.14%和42.85%;Ⅰ类群、Ⅱ类群和Ⅲ类群的白斑病病级变化范围均为1.00~9.00,变异系数分别为31.55%、53.48%和59.26%。
Pearson相关性分析结果(图2)表明,散粉期与株高、穗位高呈显著正相关,与茎腐病、南方锈病和白斑病病级均呈显著负相关,相关系数分别为0.19、0.32、-0.28、-0.32和-0.21;白斑病病级与穗行数呈显著正相关,与株高和穗位高呈显著负相关,相关系数分别为0.38、-0.24和-0.20;株高与穗位高、穗长和穗粗呈显著正相关,相关系数为0.73、0.23和0.26;穗长与南方锈病病级呈显著负相关,与秃尖长、穗粗和行粒数呈显著正相关,相关系数分别为-0.19、0.26、0.27和0.59;穗粗与穗行数呈显著正相关,相关系数为0.28;行粒数与秃尖长呈显著负相关,相关系数为-0.26。
2.3 不同类群间的性状差异
对3个类群的散粉期、株高、穗位高、穗长、穗粗、穗行数、行粒数、秃尖长、茎腐病、南方锈病和白斑病等性状进行Scheffe比较,结果(表3)显示,3个类群间的多个性状存在显著差异。Iodent(Ⅰ)和Suwan(Ⅱ)间的散粉期、株高、穗位高、穗行数、茎腐病、南方锈病和白斑病等性状存在着显著或极显著差异;Iodent(Ⅰ)与Suwan(Ⅱ)相比较,Iodent(Ⅰ)的散粉期较短,株高、穗位高较低,穗行数较多,但对茎腐病、南方锈病和白斑病的抗性较差。Iodent(Ⅰ)和温热改良系(Ⅲ)间的散粉期、株高、穗行数、茎腐病和白斑病等性状存在着显著或极显著差异;Iodent(Ⅰ)与温热改良系(Ⅲ)相比较,Iodent(Ⅰ)的散粉期较短,株高较低,穗行数较多,但对茎腐病和白斑病的抗性较差,温热改良系(Ⅲ)则相反。Suwan(Ⅱ)和温热改良系(Ⅲ)间的散粉期、穗位高和南方锈病等性状存在着显著或极显著差异;Suwan(Ⅱ)和温热改良系(Ⅲ)相比较,Suwan(Ⅱ)的散粉期较长,穗位高较高,南方锈病抗性较强。
综上,Iodent(Ⅰ)生育期短,株型清秀,穗行数多,抗病性差;Suwan(Ⅱ)生育期长,植株高大,抗病性强;温热改良系(Ⅲ)生育期较短,抗性一般。
3 讨论与结论
3.1 温热玉米种质改良形成新的杂种优势类群
近年来,研究者们利用SSR、SNP和InDel等类型的标记,对大量玉米种质资源进行系统性的评价与分析,剖析其遗传多样性与群体结构,便于后续的种质改良与创新。赵小敏等利用41对SSR引物对56份自交系进行遗传多样性分析,将其划分为7类[16]。刘志斋等利用40个覆盖玉米全基因组的核心SSR标记进行分子鉴定,将820份自交系划分为塘四平头、Reid、Lancaster、旅大红骨与P群等5个类群[17]。师亚琴等利用SNP标记将79份玉米自交系划分为两大类7个亚群(Reid、黄改类、P群、Lancaster、Iodent、迪卡选系和先锋改良群)[18]。本研究基于6万个SNP的基因型鉴定结果,利用Structure 2.3.4软件对112份温热自交系进行群体结构分析,首先将供试群体分为3个亚群,其结果与系谱来源大致相同。其中,Ⅰ类群主要包括Iodent种质;Ⅱ类群主要包括Suwan等热带种质;Ⅲ类群以温热改良系为主。此外,112份材料可进一步划分为6类,分别以HCL645、T32、QR273、黄早四、PH4CV和B73为代表;甚至可划分为9类,分别以HCL645、T32、QB446、QR273、黄早四、PH4CV、郑58、齐319和B73为代表,说明玉米在自然或人工驯化過程中不断形成新的种质[19]。即当一个杂种优势群体融入其他的遗传成分,其背景产生差异化,新的类群增加。
3.2 温热改良玉米种质促进优势性状融合
Pearson相关性分析结果(图2)表明,不同性状之间呈现出一定的相关性。散粉期与株高、穗位高呈显著正相关,与茎腐病、南方锈病和白斑病病级呈显著负相关;白斑病病级与穗行数呈显著正相关,与株高、穗位高呈显著负相关;株高与穗位高、穗长和穗粗呈显著正相关;穗长与南方锈病病级呈显著负相关,与秃尖长、穗粗和行粒数呈显著正相关;穗粗与穗行数呈显著正相关;行粒数与秃尖长呈显著负相关。Iodent种质与Suwan种质相比较,Iodent的散粉期较短,株高、穗位高较低,穗行数较多,但对茎腐病、南方锈病和白斑病的抗性较差。Iodent散粉期和株型等性状均优于Suwan,但其抗病性较差;相反,Suwan散粉期和株型等性状较差,但抗病性较强;两者遗传背景差异大,优势互补。因此,可将Suwan导入Iodent改善其抗性差的问题;同样,可将Iodent导入Suwan改善热带种质生育期晚、植株繁茂等问题。郭向阳等利用Iodent种质和Suwan种质构建基础群体,进行温热融合,成功创制出配合力高、株型清秀、无感光性且抗性强的QB5725、QB5746和QB5775等SI(Suwan-Iodent)种质,很好地将Iodent与Suwan的特点聚集在一起,且SI种质所配杂交种已大面积生产,如康农玉8009、YD819等[20]。报道显示,温带种质生育期短、株型清秀、丰产性好,但抗病性差;热带种质抗病性强、品质好,但生育期长、植株高大[21-22],本研究结果与之一致。即热带种质与温带种质的遗传背景差异大,性状优势互补,选择潜力和利用价值丰富;将两者的有利基因聚集,改变原有的适应性和农艺性状,提高温带资源的抗性,改善热带种质的株型与生育期,扩大玉米资源的遗传基础,与前人的报道[23-24]基本一致。然而,因光周期敏感等问题热带种质在温带地区缺乏适应性,在进行温热互融时,难以选出符合目标性状的理想株系[25-26]。前人研究表明,通过与早熟或适应性资源进行杂交或回交,是解决热带种质光周期问题的重要途径之一[27-28]。Iodent作为早熟资源,用于热带种质改良可有效解决热带种质光周期等问题,郭向阳等利用Suwan种质与不同温带玉米种质组配,对不同类型杂交种的生育期、株高、穗位高等性状进行评价,发现Suwan-Iodent模式的杂交种优于Suwan与其他类型的杂交组合,且光周期不敏感,可正常开花结实;Suwan与其他类型的杂交组合在温带地区则存在着由光周期反应引起的一系列问题[29]。现阶段,虽已有许多成功的案例将热带种质与温带种质融合,但温热种质融合受生态环境和基因连锁影响较大,仍需要大量的田间实践与研究。
参考文献:
[1]Matsuoka Y,Vigouroux Y,Goodman M M,et al. A single domestication for maize shown by multilocus microsatellite genotyping[J]. Proceedings of the National Academy of Sciences of the United States of America,2002,99(9):6080-6084.
[2]Doebley J. The genetics of maize evolution[J]. Annual Review of Genetics,2004,38:37-59.
[3]Doebley J F,Gaut B S,Smith B D. The molecular genetics of crop domestication[J]. Cell,2006,127(7):1309-1321.
[4]张爱民,阳文龙,方红曼,等. 作物种质资源研究态势分析[J]. 植物遗传资源学报,2018,19(3):377-382.
[5]王晓鸣,邱丽娟,景蕊莲,等. 作物种质资源表型性状鉴定评价:现状与趋势[J]. 植物遗传资源学报,2022,23(1):12-20.
[6]Liu C L,Hao Z F,Zhang D G,et al. Genetic properties of 240 maize inbred lines and identity-by-descent segments revealed by high-density SNP markers[J]. Molecular Breeding,2015,35(7):146.
[7]Wu X,Li Y X,Shi Y S,et al. Fine genetic characterization of elite maize germplasm using high-throughput SNP genotyping[J]. Theoretical and Applied Genetics,2014,127(3):621-631.
[8]张亚菲,刘松涛,曹雯梅,等. 黄淮海夏玉米品种主要性状遗传多样性研究[J]. 种子,2021,40(4):96-100.
[9]郭欢乐,汤 彬,李涵,等. 湖南省玉米地方品种表型性状综合评价及类群划分[J]. 作物杂志,2022(6):33-41.
[10]王新涛,王 艳,杨 青,等. 玉米抗粗缩病种质资源的发掘和遗传多样性分析[J]. 分子植物育种,2017,15(12):5172-5177.
[11]王 婷,王铁兵,王 鹏,等. 玉米种质资源对瘤黑粉病的抗性鉴定与遗传多样性分析[J]. 玉米科学,2022,30(6):30-38.
[12]王强盛,苏雪莲,张 慧.地方农业种质资源保护存在问题与利用途径[J]. 江苏农业科学,2023,51(4):23-27.
[13]李永祥,李会勇,扈光辉,等. 玉米应用核心种质的构建与应用[J]. 植物遗传资源学报,2023,24(4):911-916.
[14]Wu X,Wang A G,Guo X Y,et al. Genetic characterization of maize germplasm derived from Suwan population and temperate resources[J]. Hereditas,2019,156:2.
[15]石云素.玉米種质资源描述规范和数据标准[M]. 北京:中国农业出版社,2006.
[16]赵小敏,柏光晓,任 洪,等. 西南区应用的7个玉米骨干系与美国自交系的SSR标记分析[J]. 山地农业生物学报,2017,36(1):24-29.
[17]刘志斋,吴 迅,刘海利,等. 基于40个核心SSR标记揭示的820份中国玉米重要自交系的遗传多样性与群体结构[J]. 中国农业科学,2012,45(11):2107-2138.
[18]师亚琴,孟庆立,杨少伟,等. 利用SNP标记划分玉米自交系杂种优势类群[J]. 中国种业,2022(8):79-82.
[19]Li C H,Guan H H,Jing X,et al. Genomic insights into historical improvement of heterotic groups during modern hybrid maize breeding[J]. Nature Plants,2022,8(7):750-763.
[20]郭向阳,吴 迅,王安贵,等. 玉米Suwan和Iodent改良系育种应用探索与分析[J]. 植物遗传资源学报,2021,22(6):1676-1683.
[21]祝云芳,陈泽辉,郭向阳,等. 基于温热玉米Suwan-Lancaster群体改良自交系QB446的选育与应用[J]. 贵州农业科学,2020,48(7):1-5.
[22]吴文强,吴 迅,郭向阳,等. 玉米温热种质改良后代籽粒脱水相关性状的评价[J]. 山地农业生物学报,2018,37(6):76-79.
[23]陈泽辉,祝云芳,王安贵,等. 玉米Tuxpeno和Suwan种质的改良研究[J]. 贵州农业科学,2010,38(2):1-4.
[24]郭向阳,胡 兴,祝云芳,等. 热带玉米Suwan1群体导入不同类型温带种质的遗传分析[J]. 玉米科学,2019,27(4):9-13,21.
[25]Holland J B,Goodman M M. Combining ability of tropical maize accessions with U.S.germplasm[J]. Crop Science,1995,35(3):767-773.
[26]段民孝,刘新香,张华生,等. 快速钝化玉米热带亚热带种质光敏性方法及其育种利用研究[J]. 种子,2018,37(8):103-106.
[27]陈洪梅,汪燕芬,姚文华,等. 导入热带种质的温带玉米自交系的利用潜力[J]. 作物学报,2011,37(10):1785-1793.
[28]高增玉,武玉华,和剑涵,等. 利用桥梁亲本导入玉米热带种质优良基因的研究[J]. 河北农业科学,2013,17(4):53-55,58.
[29]郭向阳,赵 强,吴迅,等. 玉米Iodent种质在Suwan材料改良利用中的有效性分析[J]. 种子,2020,39(4):41-45.
猜你喜欢
江苏农业科学(2016年8期)2017-02-15 20:03:31
江苏农业科学(2016年8期)2017-02-15 19:33:33
江苏农业科学(2016年8期)2017-02-15 19:32:36
长江蔬菜·学术版(2016年12期)2017-01-12 20:57:59
中国科技博览(2016年25期)2016-12-20 20:04:30
农家科技下旬刊(2016年9期)2016-12-15 10:42:25
河北渔业(2016年9期)2016-11-05 10:40:55
中国科技博览(2016年11期)2016-05-06 02:35:39
现代园艺(2016年3期)2016-03-24 16:02:02
天津农业科学(2015年5期)2015-05-30 17:34:07
