
大豆抗氧化肽的制备及对大蒜根尖过氧化的保护作用
2024-04-29 06:14:56邹文广,尹乐斌,贺超,刘聪,龙盼,李敏娟
邵阳学院学报(自然科学版) 2024年1期
邹文广,尹乐斌,贺超,刘聪,龙盼,李敏娟


摘要:为探究不同蛋白酶酶解大豆分离蛋白(soybean isolate protein,SPI)对多肽得率及体外抗氧化活性的影响,采用6种蛋白酶(碱性蛋白酶、木瓜蛋白酶、菠萝蛋白酶、中性蛋白酶、胰蛋白酶、胃蛋白酶)进行筛选,测定酶解产物多肽得率、光谱特性及体外抗氧化能力,分析其对过氧化氢诱导的大蒜根尖氧化损伤的保护作用。结果表明,碱性蛋白酶酶解条件下,多肽得率最高为67.78%;多肽在200~230 nm处有最大吸收峰;6种多肽在酰胺A、B、Ⅰ、Ⅱ、Ⅲ带均有吸收峰;所有多肽均具有较好地清除自由基能力,且随着多肽浓度的增加而增加。综合其抗氧化活性评价,碱性蛋白酶制备的多肽清除能力均优于其他蛋白酶;经多肽处理后大蒜根尖细胞凋亡程度下降45.90%。
关键词:蛋白酶;酶解;多肽;体外抗氧化活性;细胞凋亡
中圖分类号:TS214.2 文献标志码:A
Preparation of soybean antioxidant peptides and their protective effect against garlic root tip peroxidation
ZOU Wenguang1, YIN Lebin1,2, HE Chao1, LIU Cong1, LONG Pan1, LI Minjuan1
(1. School of Food and Chemical Engineering, Shaoyang University, Shaoyang 422000, China; 2. Key Laboratory of Soybean Product Processing and Safety Control in Hunan Province, Shaoyang 422000, China)
Abstract: In order to investigate the effects of enzymatic hydrolysis of soybean protein isolate with different proteases on the yield of polypeptide and antioxidant activity in vitro, six proteases (alkaline protease, papain, bromelain, neutral protease, trypsin, pepsin) were selected to determine the yield, spectral characteristics and antioxidant capacity of the polypeptide products of enzymatic hydrolysis. The protective effect on oxidative damage of garlic root tips induced by hydrogen peroxide was analyzed. The results show that the highest polypeptide yield was 67.78% under the condition of alkaline protease hydrolysis. The maximum absorption peak of polypeptide was at 200-230 nm. The absorption peaks of the 6 peptides were found in the amide A, B, Ⅰ, Ⅱ and Ⅲ bands. All peptides have good scavenging ability of free radicals, and the ability increases with the increase of peptide concentration. According to the evaluation of antioxidant activity, the scavenging ability of polypeptide prepared by alkaline protease is better than that of other proteases. The apoptosis of garlic root tip cells decreases by 45.90% after polypeptide treatment.
Key words: protease; enzymatic hydrolysis; polypeptide; in vitro antioxidant activity; cell apoptosis
大豆分离蛋白[1](soybean isolate protein,SPI)是经过各种物理或化学方法处理得到的一种优良蛋白质,其中含有8种人体必需氨基酸[2],其具有获取价格低廉、生物活性强等特点[3]。由于SPI具有大量的带电氨基酸和疏水氨基酸残基,因此,SPI是传递疏水生物活性物质的优良载体[4]。
生物活性肽可以通过酸水解、酶解或其他方法从蛋白质中获得。采用酶法制备的生物活性肽,具有条件温和、专一性强、副产物少等优势[5],且酶解后可以改善其功能特性[6]。不同的酶在蛋白质中酶切位点不同,肽段所呈现的功能特性也会不同,大豆多肽包括抗菌肽、抗氧化肽等[7]。大豆抗氧化肽是酶解SPI后得到的具有良好生物活性的物质;同时,也具有比酶解前蛋白更好的理化特质[8]。郑辉等[9]利用不同蛋白酶水解碧根果蛋白制备蛋白肽,研究发现碱性蛋白酶水解碧根果蛋白6 h时水解度最高为32.54%。不同蛋白酶酶解得到的多肽,具有不同的分子量以及生物活性。探究不同蛋白酶酶解产物的生物活性,可以促进多肽产品的开发利用。
通过对6种蛋白酶制备的大豆抗氧化肽进行筛选,比较其酶解后多肽的得率、光谱特性和体外抗氧化活性以评估其抗氧化能力。筛选出抗氧化能力最强的多肽,探究其对过氧化氢诱导大蒜根尖细胞的过氧化保护能力,为大豆抗氧化肽制备及多肽产品的开发提供一定的理论依据。
1材料与方法
1.1原料与试剂
SPI(河南糖柜食品有限公司);蛋白酶(食品级,河南汇泉生物科技有限公司);邻苯三酚(上海麦克林生化科技股份有限公司);水杨酸[福晨(天津)化学试剂有限公司];无水乙醇(成都金山化学试剂有限公司);其他试剂均为分析纯。
1.2主要仪器与设备
LS220A电子天平(上海天美天平仪器有限公司);UV-2600i 紫外可见分光光度计[岛津仪器(苏州)有限公司];3H16RI智能高速冷冻离心机(湖南赫西仪器装备有限公司);SCIENTZ-18N 真空冷冻干燥机(宁波新芝生物科技股份有限公司);VM-01U(HYQ-3110)涡旋混匀器(苏州捷美电子有限公司)。
1.3试验方法
1.3.1蛋白酶筛选
SPI→蛋白酶酶解→离心→超滤→真空冷冻干燥→抗氧化肽[10],不同蛋白酶处理的酶解条件见表1[11]。
1.3.2多肽得率的测定
参照郭耀华等[12]的方法进行测定。
1.3.3多肽紫外光谱测定
称取0.01 g多肽冻干粉,定容至10 mL,用紫外-可见分光光度计测定200~800 nm波长的吸光度,测定多肽紫外光谱。
1.3.4多肽红外光谱测定
参照熊喆等[13]方法进行测定。
1.3.5抗氧化活性测定
多肽清除ABTS+自由基能力,参照KUT等[14]的方法;清除DPPH自由基能力,结合文献[15]的方法;清除超氧阴离子自由基能力测定参照程超等[16]进行测定;清除羟自由基率能力结合文献[17]进行测定。
1.3.6大蒜根尖细胞凋亡程度测定
取根系生长较良好的大蒜,分为对照组(水培养6 h)、空白组(过氧化氢处理4 h、水培养2 h)、实验组(过氧化氢处理4 h、1 mg/mL多肽处理2 h)和损伤组(过氧化氢处理6 h),然后在室温下浸泡在0.25%的伊文思蓝水溶液中15 min。将根尖浸入5 mL N,N-二甲基甲酰胺中1 h后,收集浸出液,检测600 nm下吸光度,以测定细胞凋亡程度。
1.4数据处理
所有试验均进行了3次。采用IBM SPSS 19、Origin 2021等软件分析及绘图处理。
2结果与分析
2.1不同蛋白酶对多肽得率的影响
采用不同蛋白酶处理SPI后测定其多肽得率见图1。由图1可知,不同蛋白酶处理,其多肽得率由高到低为碱性蛋白酶、胰蛋白酶、中性蛋白酶、胃蛋白酶、木瓜蛋白酶、菠萝蛋白酶。碱性蛋白酶处理后多肽得率最高,67.78%;菠萝蛋白酶酶解条件下多肽得率最低,5.71%;不同蛋白酶酶解条件测定的多肽产率均有显著性差异。不同的酶具有不同的酶切位点,试验所选6种酶均属于内切酶。内切酶主要在蛋白分子的肽键位置起作用,导致蛋白分子分解为多肽片段,内部疏水基团被暴露出来[12]。菠萝蛋白酶酶解后多肽产率较低,可能是由于其作用酶切位点较多,水解程度更彻底,将大豆多肽水解成氨基酸[18];经过碱性蛋白酶的处理多肽得率较高,可能是由于其自身的特性,能够将SPI水解后得到更多分子量较小的多肽[18]。
2.2不同蛋白酶制备的多肽结构特性
2.2.1多肽紫外扫描光谱分析
蛋白质存在的不饱和结构肽键、羧基等都会对紫外光区有吸收[19-20]。不同样品的紫外波长扫描见图2。不同蛋白酶解多肽最大吸收峰出现在210~230 nm,符合大豆多肽的特征吸收峰;在260~280 nm波长处也有一定的吸收,但吸收峰较小,说明多肽中含有一定量的芳香族氨基酸;经碱性蛋白酶、胃蛋白酶和胰蛋白酶处理,其多肽峰值比其他3种更低。可能原因是这3种蛋白酶酶解后,得到的大豆多肽具有更小的分子量;且在260~280 nm处吸收峰,后者紫外光谱吸收带的最大吸收峰波长向长波方向移动。这种现象的主要原因是,肽键中的羰基π电子与相邻氨基的孤对电子发生了p-π共轭作用。由于共轭效应,使得电子在π-π*跃迁过程中产生的吸收峰发生了红移[20]。
2.2.2多肽红外光谱分析
多肽分子能够选择性吸收某些红外波长,从而导致分子中振动能级和转动能级的跃迁。因此,可分析红外光谱来探究多肽结构[10]。由图3可知,6种样品有相似的红外光譜特征吸收峰,其中,酰胺A带均在3 293.51 cm-1附近,其原因与形成氢键,导致N—H键的伸缩振动相关[10];酰胺B带在2 933.77 cm-1附近,其中,中性蛋白酶解多肽峰值表现较弱;酰胺Ⅰ带出现1 644.34~1 661.70 cm-1附近,是CO基团的特征谱带;酰胺Ⅲ带出现在1 238~1 249 cm-1附近,其原因来自C—N键的伸缩振动、N—H键的弯曲振动,以及氨基酸上—CH2基团的摇摆振动;酰胺Ⅱ带(1 500~1 600 cm-1)有明显的吸收峰,但也存在一定差异。图3中6种多肽在1 000~1 500 cm-1区域有很明显的吸收峰吸收差异,可能由于不同蛋白酶酶解后多肽侧链基团的分子构成存在显著差异,最终引起多肽之间抗氧化活性的差异[21]。
2.3不同蛋白酶制备多肽的抗氧化活性
2.3.1多肽清除ABTS+自由基的能力
ABTS+法在抗氧化试验中使用较广泛,对极性和非极性物质都可以检测[14]。由图4可知,样品对ABTS+自由基均具有清除效果,且清除能力随着样品质量浓度增加而逐渐增强。从总体的清除能力来看,碱性蛋白酶处理获得的多肽,其ABTS+自由基清除能力高于其他蛋白酶的。因此,SPI经碱性蛋白酶解后,具有更强的ABTS+自由基清除能力。
IC50可以反映清除能力,其數值越小,说明清除能力越强[22]。从表2可知,6种蛋白酶解液的IC50值之间存在显著性差异(P<0.05)。从6种蛋白酶的IC50来看,碱性蛋白酶的IC50最小,(0.568 9±0.009 8)mg/mL,菠萝蛋白酶的IC50最大,(0.598 7±0.006 1)mg/mL,说明碱性蛋白酶处理的多肽的ABTS+自由基清除能力更强,具有更强的抗氧化能力。
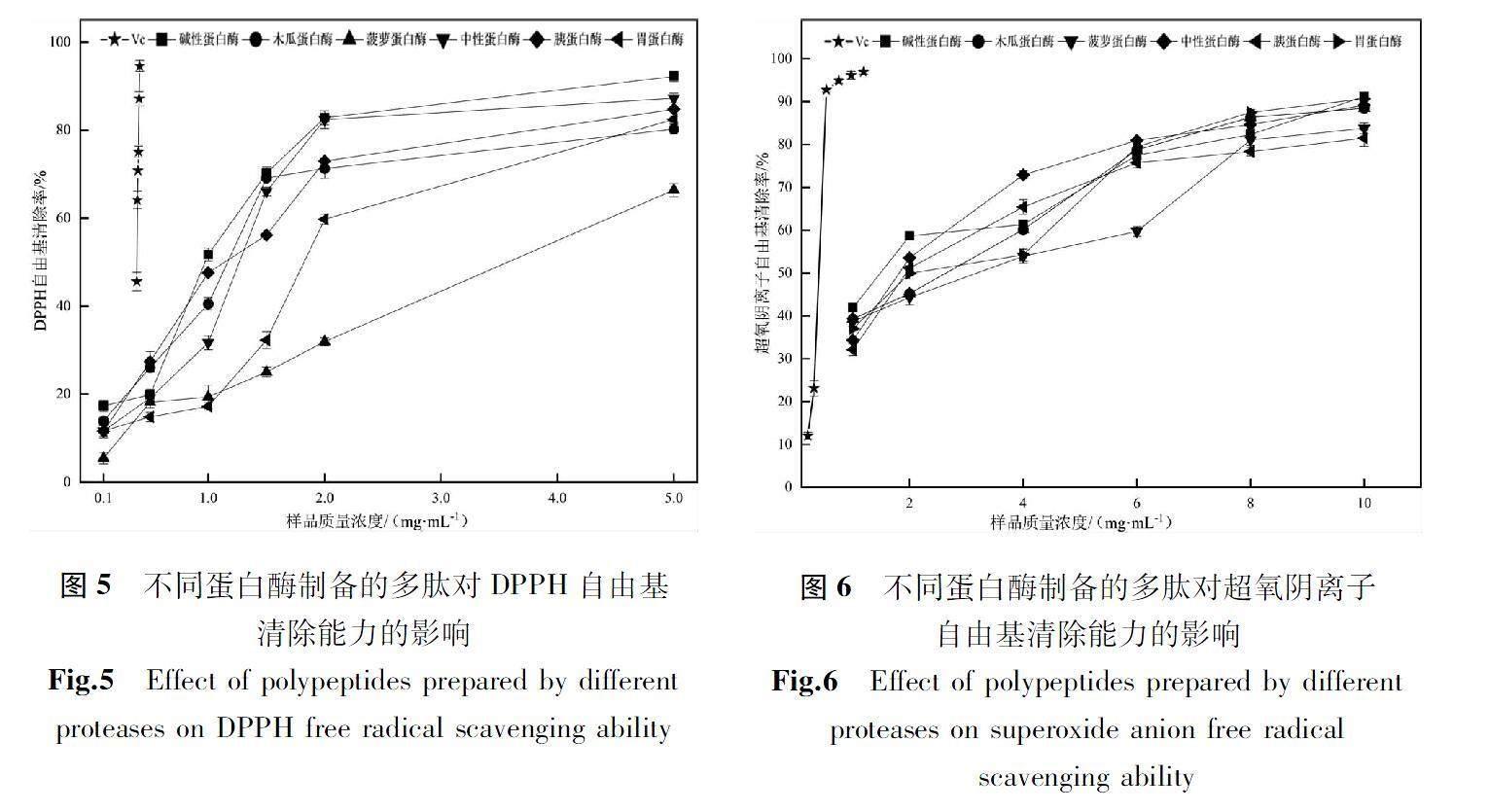
2.3.2多肽清除DPPH自由基的能力
不同样品清除DPPH自由基的能力见图5。由图5可知,样品对DPPH自由基均具有清除效果,随着样品质量浓度增加,其清除能力增强。碱性蛋白酶得到的多肽,其DPPH自由基清除能力优于其他蛋白酶;而对于菠萝蛋白酶酶解得到的多肽,它的自由基清除效果最差。可能原因是碱性蛋白酶酶解所得多肽,其N-端多为疏水性氨基酸,因而其具有较高的抗氧化活性[23]。
从表3可知,不同样品的IC50之间存在显著性差异(P<0.05),从6种蛋白酶的IC50来看,木瓜蛋白酶和胰蛋白酶酶解多肽的IC50之间无显著性差异;碱性蛋白酶的IC50最小;说明经碱性蛋白酶处理获得的多肽,有更强的DPPH自由基清除能力,具有更强的抗氧化能力。
2.3.3多肽清除超氧阴离子自由基清的能力
超氧阴离子自由基是活性氧之一,具有强烈的氧化能力[16]。由图6可知,样品对超氧阴离子自由基清除能力均随着样品质量浓度增加而逐渐增强。从总体的清除能力来看,经碱性蛋白酶与中性蛋白酶酶解的多肽,其超氧阴离子自由基清除能力优于其他蛋白酶酶解的多肽,说明碱性蛋白酶与中性蛋白酶水解产物对超氧阴离子有较好的清除作用。
从表4可知,不同样品的IC50之间存在显著性差异(P<0.05),从6种多肽的IC50来看,碱性蛋白酶处理后样品的IC50最小,(1.640 8±0.067 5)mg/mL,其清除能力最强。不同酶水解后的多肽抗氧化能力都不同,可能原因是水解后肽段的长度及本身的性质不同[17]。
2.3.4多肽清除羟自由基的能力
羟自由基是脂质氧化反应的常见中间产物,易与生物大分子发生反应[17]。如图7所示,6种蛋白酶解多肽对羟自由基的清除效果随质量浓度增加而增加。由表5可知,碱性蛋白酶和木瓜蛋白酶酶解多肽的IC50二者之间无显著性差异。但与其他蛋白酶酶解产物的IC50值存在显著性差异,说明相同质量浓度下二者的抗氧化能力无显著性差异,且其对羟自由基清除能力优于其他蛋白酶。研究显示,经酶解处理的SPI,使得样品中的活性基团或位点更加暴露,可作为氢供体还原自由基,从而终止自由基连锁反应,以此实现其抗氧化能力[24]。
2.4大豆多肽对过氧化氢诱导大蒜根尖氧化应激细胞凋亡的影响
以吸收伊文思蓝程度表示细胞凋亡程度[25]。由图8可知,不同处理组之间存在显著性差异(P<0.05)。与对照组相比,空白组、实验组和损伤组的伊文思蓝吸光度分别提高了44.91%、9.58%和59.89%;经大豆多肽处理后,吸光度降低45.90%,与对照组相近。结果表明,大豆抗氧化肽可有效降低过氧化氢诱导的细胞凋亡,原因可能是大豆抗氧化肽作为生物活性物质,可以被机体吸收后,参与机体内有关抗细胞凋亡代谢途径,有效抑制过氧化氢胁迫导致的氧化应激[25]。
3结论
1)6种蛋白酶酶解后多肽得率由高到低为碱性蛋白酶、胰蛋白酶、中性蛋白酶、胃蛋白酶、木瓜蛋白酶、菠萝蛋白酶,其中碱性蛋白酶酶解SPI的多肽得率最高为67.78%;菠萝蛋白酶酶解条件下多肽得率最低为5.71%。
2)多肽在200~230 nm处有最大吸收峰;6种多肽在酰胺A、B、Ⅰ、Ⅱ、Ⅲ带均有吸收峰,在1 000~1 500 cm-1区域有很明显的吸收峰吸收差异。
3)6种蛋白酶酶解产物的自由基清除能力,均随浓度增加而增加。从总体自由基清除率上看,碱性蛋白酶解产物对ABTS+、DPPH、超氧阴离子以及羟自由基4种自由基的清除效果均优于其他蛋白酶,其IC50值分别为0.568 9、1.010 4、1.640 8和3.936 5 mg/mL。
4)经碱性蛋白酶酶解制备的大豆抗氧化肽处理后,与对照相比,空白组、实验组和损伤组的伊文思蓝吸光度显著提高了44.91%、9.58%和59.89%;经大豆多肽处理后大蒜根尖细胞凋亡程度下降45.90%。
因此,经碱性蛋白酶酶解得到的多肽,具有更强的抗氧化活性。研究能为SPI酶解制备抗氧化肽提供一定的理论依据。
参考文献:
[1]上官玲玲, 张辉燕, 王文欣, 等. 大豆分离蛋白酶解工艺优化及在发酵调味料中的应用[J]. 食品工业科技, 2023, 44(19): 272-280.
[2]BI C H, ZHOU T, WU Z Y, et al. Acid-mediated formation of soybean isolate protein emulsion gels with soybean oil as an active component[J]. Foods, 2023, 12(9): 1754.
[3]王金玲, 王雨淅, 王梓同, 等. 大豆多肽的制备及功能性研究进展[J]. 中国酿造, 2022, 41(1): 25-31.
[4]WEN P P, XIA C, ZHANG L, et al. Effects of different dry heating temperatures on the spatial structure and amino acid residue side-chain oxidative modification of soybean isolated proteins[J].Food Chemistry, 2023, 405(Pt A): 134795.
[5]赵丛丛, 曾里, 宋娜, 等. 碱性蛋白酶提取大米水解蛋白的研究[J]. 食品与发酵科技, 2010, 46(1): 85-88.
[6]GUO H, FAN L, DING L, et al. Separation and purification of antioxidant peptide from fermented whey protein by Lactobacillus rhamnosus B2-1[J]. Food Science of Animal Resources, 2023, 43(1): 10-24.
[7]PENG B, CAI B N, PAN J Y. Octopus-derived antioxidant peptide protects against hydrogen peroxide-induced oxidative stress in IEC-6 cells[J]. Food Science & Nutrition, 2022, 10(11): 4049-4058.
[8]马福建. 酶法生产大豆抗氧化肽的工艺研究[D]. 长春: 长春大学, 2019.
[9]郑辉, 张鉴达, 宫若楠. 不同蛋白酶对碧根果蛋白肽制备的影响及抗氧化活性研究[J]. 中国食品添加剂, 2023, 34(4): 235-241.
[10]杨学为, 尹乐斌, 罗雪韵, 等. 超声辅助半仿生酶法制备大豆多肽及其抗氧化活性研究[J]. 粮食与油脂, 2023, 36(4): 126-131.
[11]芦鑫, 姜梦楠, 张丽霞, 等. 制备芝麻抗氧化肽的蛋白酶筛选[J]. 中国油脂, 2018, 43(11): 28-33.
[12]郭耀华, 尚鑫茹, 岳兰昕, 等. 3种蛋白酶对鲶鱼骨肉泥中多肽得率和钙溶出率的比较研究[J]. 食品科技, 2016, 41(2): 157-162.
[13]熊喆, 赵钰, 秦子波, 等. 超声辅助酶解促进草鱼鳞胶原肽水解进程的内在机制解析[J]. 农业工程学报, 2022, 38(16): 313-321.
[14]KUT K, STEFANIUK I, BARTOSZ G, et al. Formation of a purple product upon the reaction of ABTS radicals with proteins[J]. International Journal of Molecular Sciences, 2023, 24(10): 8912.
[15]李洁, 余有贵, 徐海月, 等. 蓝莓果酒的纯种发酵工艺优化与抗氧化特性[J]. 邵阳学院学报(自然科学版), 2023, 20(1): 74-80.
[16]程超, 薛峰, 李伟, 等. 3种处理方式对葛仙米藻膽蛋白清除超氧阴离子自由基能力的影响[J]. 食品科学, 2014, 35(13): 26-31.
[17]江含秀, 曾芳, 邬晓霞, 等. 不同品种苦荞蛋白的复合酶解及其多肽抗氧化活性[J]. 食品研究与开发, 2023, 44(9): 43-51.
[18]张文敏, 张健, 周浩纯, 等. 亚麻籽粕制备小分子抗氧化活性肽[J]. 食品科学, 2020, 41(8): 36-44.
[19]曾允灏, 陈德经, 董梦尧, 等. 大鲵肉肌基质蛋白中胶原蛋白肽和弹性蛋白肽特性[J]. 肉类研究, 2022, 36(12): 15-21.
[20]刘文颖, 冯晓文, 程青丽, 等. 小麦低聚肽的结构特征及其体外抗氧化活性[J]. 现代食品科技, 2021, 37(12): 72-79.
[21]陶琳丽, 黄伟, 杨秀娟, 等. 20种氨基酸近红外光谱及其分子结构的相关性[J]. 光谱学与光谱分析, 2016, 36(09): 2766-2773.
[22]延莎, 范亚芳, 郑龙华, 等. 乳酸菌发酵茶花粉所得提取物对4种自由基的清除能力[J]. 食品科技, 2017, 42(3): 238-243.
[23]蔡俊, 陈季旺, 王茹, 等. 多肽体外抗氧化活性测定方法的比较[J]. 食品科学, 2016, 37(11): 52-57.
[24]张海萍, 余小月, 程梦颖, 等. 羟自由基氧化对鲢鱼肌原纤维蛋白凝胶特性的影响[J]. 食品科学, 2023, 44(6): 9-16.
[25]YAO S C, HUANG W J, PAN C L, et al. Caspase-like proteases regulate aluminum-induced programmed cell death in peanut[J].Plant Cell Tissue and Organ Culture, 2016, 127(3): 691-703.
猜你喜欢
现代园艺(2017年13期)2018-01-19 02:28:09
中国中药杂志(2016年22期)2017-02-13 17:28:52
科技视界(2016年25期)2016-11-25 09:04:46
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
科技视界(2016年15期)2016-06-30 12:27:37
中国实用医药(2016年11期)2016-05-04 21:56:32
天津农业科学(2015年11期)2015-12-03 00:09:11
中国当代医药(2015年25期)2015-10-21 18:22:58
山东体育学院学报(2015年3期)2015-08-14 20:30:25
药学与临床研究(2015年4期)2015-06-05 11:35:54
