
太湖河蚬种群组成及生长特征研究
2024-04-28 05:09刘洋李大命陆建明刘燕山唐晟凯杨家新张彤晴
生态科学 2024年1期
刘洋 , 李大命, 陆建明, 刘燕山, 唐晟凯, 杨家新, 张彤晴
太湖河蚬种群组成及生长特征研究
刘洋1, 2, 李大命2, *, 陆建明3, 刘燕山2, 唐晟凯2, 杨家新1, 张彤晴2
1. 南京师范大学海洋科学与工程学院, 南京 210023 2. 江苏省淡水水产研究所江苏省内陆水域渔业资源重点实验室, 南京 210017 3. 江苏省太湖渔业管理委员会办公室, 苏州 215104
为掌握太湖河蚬种群的大小组成、年龄结构及其生长特征, 在太湖设置6个采样点, 通过采集太湖河蚬样品, 测量河蚬的壳长、壳宽、壳高和体重, 判定河蚬的年龄, 建立河蚬壳长、体重的生长方程。研究结果表明: (1) 河蚬壳长、壳宽、壳高及体重范围分别为5.45—40.34 mm、6.68—24.89 mm、7.15—37.41 mm、0.15—19.27 g, 其均值分别为(20.17±0.60) mm、(13.57±0.33) mm、(19.13±0.53) mm和(4.09±0.32) g。(2) 河蚬种群由0+至4+龄共5个年龄组组成, 其中0+至2+龄的个体数量占比91.85%, 3+至4+龄的个体数量占比8.15%。(3) 河蚬的壳长与体重的关系式为= 2.0×10–32.477(2=0.954), 表明河蚬呈异速生长。(4) 河蚬的壳长生长方程为t= 41.99×[1––0.27(t+1.47)], 体重生长方程为t= 19.28×[1––0.27(t+1.47)]2.477, 拐点年龄为2.22龄, 对应的壳长和体重分别为26.48 mm和6.16 g。基于研究结果得出, 太湖河蚬种群个体的小型化、低龄化明显, 资源开发利用强度偏大, 需要采取措施保护太湖河蚬资源。
河蚬; 种群组成; 年龄结构; 生长方程; 太湖
0 前言
河蚬()俗称沙蜊、金蚶、黄蚬等, 隶属于软体动物门、瓣鳃纲(Lamellibranchina)、真瓣鳃目(Eulamellibrqanchina)、蚬科(Corbiculidae)、蚬属()[1]。河蚬为滤食性生物, 广泛分布于世界各地的江河、湖泊、沟渠等水域, 且常成为大型底栖动物群落的优势种。河蚬不仅是水生态系统的重要组成部分, 而且在水生态系统中具有重要的生态作用和功能[2-5]。河蚬肉质味道鲜美, 营养丰富, 且具有多种药效功能, 已被开发成多种调味及保健产品[6-8]。河蚬也是我国重要的出口创汇水产品之一, 多年来国际市场对河蚬的需求量持续增加, 导致对河蚬天然资源的捕捞强度日益加剧。另外, 加之水体环境污染、栖息地破坏等多种不利因素的影响, 造成我国湖泊河蚬资源量急剧衰退, 分布范围缩小, 捕捞个体越来越小, 严重制约了河蚬资源的可持续发展利用[9-13]。
迄今, 关于河蚬的研究主要集中在繁殖生物学[14-15]、生态毒理学[16]、营养及药用价值[6-8]、遗传多样性[17-18]及资源时空分布与环境因子等方面[19-20],而有关河蚬种群生长方面的研究较少。太湖是我国第三大淡水湖泊, 河蚬是太湖底栖动物群落的优势种群, 营养丰富, 经济价值高, 且对控制太湖富营养化及修复水体生态环境具有重要意义[4, 21]。目前有关太湖河蚬资源的时空分布及演替已有较多报道[4, 9, 11-12], 但尚未见太湖河蚬种群年龄结构及生长方面的研究, 而种群的年龄结构及生长规律是开展渔业资源评估及制定渔业资源保护措施的基础。本研究通过采集太湖河蚬样品, 分析河蚬的种群组成和年龄结构, 构建河蚬的生长方程及揭示其生长规律, 研究结果可为太湖河蚬资源的科学保护与合理利用提供重要参考依据。
1 材料与方法
1.1 河蚬样品采集和处理
本研究在太湖的梅梁湾、竺山湾、贡湖湾、湖心、河蚬保护区及西太湖水域共设置6个采样点(图1), 分别于2019年5月和10月在6个采样点开展河蚬资源调查。河蚬采集工具为渔民捕捞河蚬用的划耙, 耙宽70 cm, 耙齿间距6 mm, 每个采样点划耙拖行10 m。将划耙内的泥沙及其它杂物去除, 用湖水将河蚬冲洗干净, 活体、避光带回实验室处理。
1.2 河蚬测量
用游标卡尺测量河蚬的形态指标: 壳长(壳体前后方向的最大距离,)、壳宽(两个壳瓣间的最大距离,)和壳高(壳体北部与腹部方向的最大距离,)[22], 数据精确到0.01 mm。用电子天平测量河蚬湿重(), 数据精确到0.01 g。河蚬贝壳的生长线由有密带和疏带组成, 一般将密带向疏带过度的最后一个完整的生长环记为生长线。在显微镜下观察贝壳表面的生长环, 并辅以贝壳表面的凸凹变化准确识别生长线, 根据生长线的条数鉴定河蚬年龄[23]: 贝壳表面有1条生长环的个体, 生长环为当年形成, 记为0+龄个体; 贝壳表面有2条生长环, 记为1+龄个体, 依次类推。记录所有河蚬个体的年龄, 统计河蚬的年龄组成及不同壳长组年龄的分布。
1.3 数据处理和分析
1.3.1 河蚬壳长-体重关系拟合
采用幂函数建立壳长-体重方程:=×L,为重量(g),为壳长(mm), 参数为生长的条件因子, 参数为生长因子[24]。
1.3.2 河蚬生长状况
采用年相对增长率()和生长指标()分析河蚬的生长状况[25], 其中= (R–R–1)/R–1×100%,= (lgR–lgR–1)/0.4343,R为t龄时的壳长(mm)或重量(g),R–1为t-1龄时的壳长或重量。
Figure 1 Sampling sites ofin Lake Tai
1.3.3 河蚬的生长方程构建
利用Von Bertalanffy生长方程建立河蚬壳长、体重与年龄的关系[25]。壳长和体重生长方程分别为L=∞[1––k(t–to)],W=∞[1––k(t–to)],L和W为龄时的体长和重量,∞和∞为渐近体长和渐近重量, k为生长参数,o为理论生长起点年龄。
数据的统计分析及绘图采用Excel 2010和SPSS19.0,<0.05为差异显著水平。
2 结果与分析
2.1 河蚬种群组成
2.1.1 种群的大小组成及数量分布
共采集和测量了638个河蚬样品的壳长、壳宽、壳高及重量, 其中壳长范围为5.45至40.34 mm, 均值为(20.17±0.60) mm; 壳宽范围为6.68至24.89 mm,均值为(13.57±0.33) mm; 壳高范围为7.15至37.41 mm,均值为(19.13±0.53) mm。体质量范围为0.15至19.27 g, 均值为(4.09±0.32) g。
壳长、壳宽、壳高及河蚬重量组成见图2至图5。壳长在15至20 mm的个体数量占比最大(31.66%),壳长小于25 mm的个体数量占比为80.56%, 壳长大于30 mm的个体数量占比为6.58%; 壳宽在10至12 mm的个体数量占比最大(29.78%), 壳宽小于16 mm的个体数量占比为77.12%, 壳宽大于20 mm的个体数量占比为4.70%; 壳高小于15 mm的个体数量占比最大(27.90%), 壳高小于27 mm的个体数量占比为92.78, 壳高大于31 mm的个体数量占比为2.51%; 河蚬体重在1至4 g的个体数量占比最大(57.37%), 体重小于10 g的个体数量占比为95.29%, 体重大于12 g的个体数量占比为2.82%。可以看出, 小型个体在河蚬种群中占据明显优势。
2.1.2 种群年龄结构
太湖河蚬的年龄结构如图6所示。可以看出, 太湖河蚬种群由0+至4+龄共5个年龄组构成, 其中2+龄和0+龄个体的数量较多, 占比分别为33.23%和31.66%, 其次是1+龄个体, 占比为26.95%。3+和4+龄的个体数量较少, 占比分别为4.70%和3.45%, 可以看出, 低龄个体在河蚬种群中占据明显优势。
进一步分析河蚬各壳长组的年龄分布情况(图7), 可以看出, 当壳长小于15 mm时, 0+龄的个体占据优势, 占比为98.90%; 当壳长大于15 mm小于20 mm时, 1+龄个体占据优势, 占比为60.40%; 当壳长大于20 mm小于等于30 mm时, 2+龄个体占据优势, 占比为72.60%至75.60%; 当壳长大于30 mm小于35 mm时, 3+和4+龄个体占据优势, 占比均为41.20%; 当壳长大于35 mm时, 4+龄个体占比为100%。
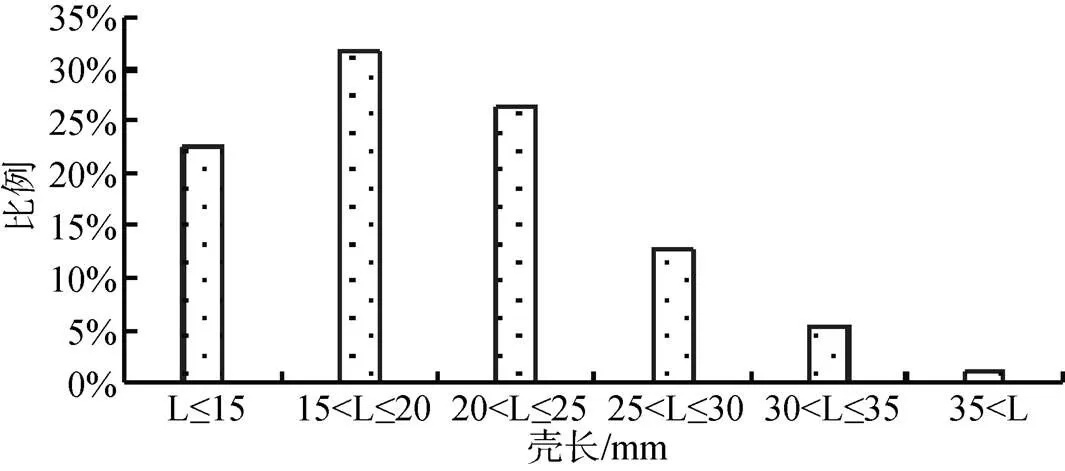
图2 太湖河蚬壳长组成
Figure 2 Shell length distribution ofin Lake Tai

图3 太湖河蚬壳宽组成
Figure 3 Shell width distribution ofin Lake Tai

图4 太湖河蚬壳高组成
Figure 4 Shell heigth distribution ofin Lake Tai
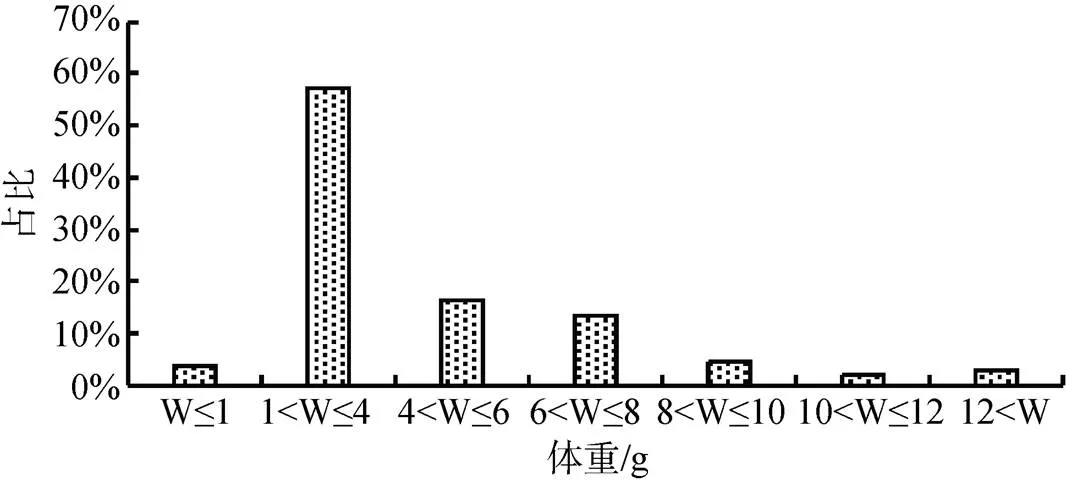
图5 太湖河蚬体重组成
Figure 5 Wet weight distribution ofin Lake Tai
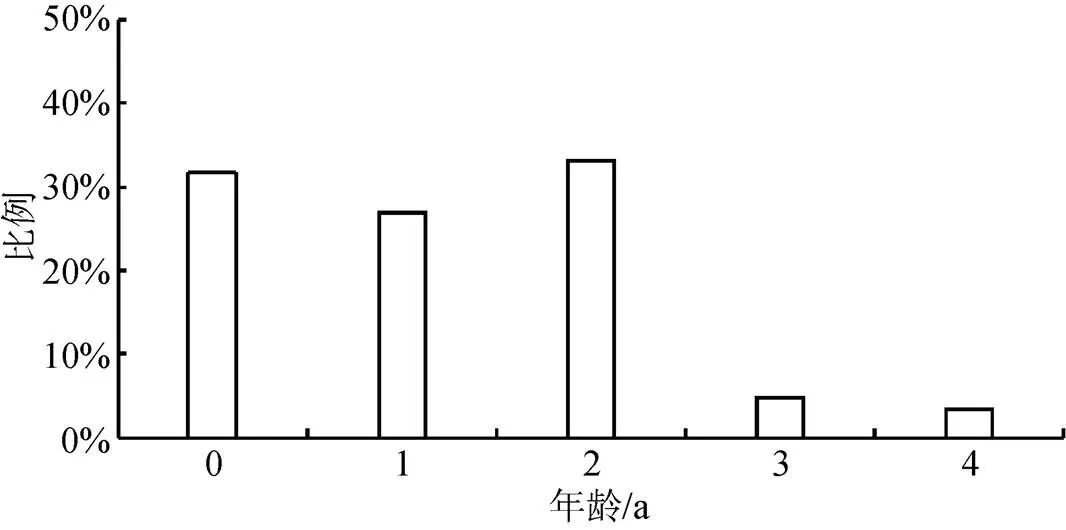
图6 太湖河蚬年龄分布
Figure 6 Age distribution ofin Lake Tai
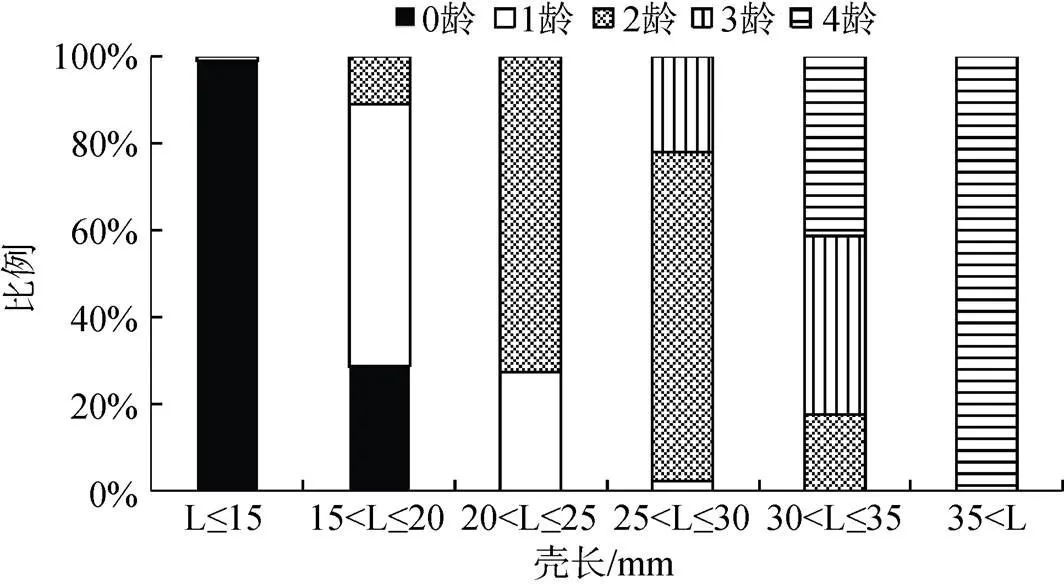
图7 河蚬各壳长组的年龄分布
Figure 7 Age frequency distribution ofin different body length groups
2.2 河蚬生长特征
2.2.1 壳长与体重的关系
河蚬体长与体重的关系式为:= 2.0×10–32.477(=638,2=0.954) (图8)。可以看出, 体长与体重具有幂函数相关性, 幂指数为2.477, 统计检验表明, 幂指数显著小于“3”(<0.05), 说明太湖河蚬呈异速增长模式。
2.2.2 生长状况
利用年相对生长率和生长指标指示太湖河蚬的生长状况, 结果如表1所示。可以看出, 随着年龄的增加, 壳长和体重的年相对生长率及生长指标呈现出逐渐下降的趋势。从0+至1+龄, 河蚬壳长和重量的年相对增长率以及生长指标最大; 从1+至2+龄, 河蚬的生长速度有小幅下降, 之后呈持续明显下降趋势, 说明河蚬生长与年龄阶段密切相关, 其中0+至2+龄是河蚬的快速生长期, 2+至3+龄为稳定生长期, 3+至4+龄为衰老期。
2.2.3 Von Bertalanffy生长方程
采用线性回归法获得河蚬生长方程的参数:∞= 41.99 mm,∞= 19.28 g, k= 0.27,0= –1.47, 建立河蚬壳长和体重的Von Bertalanffy生长方程, 分别为:L= 41.99×[1––0.27(t+1.47)](2=0.945)和W= 19.28×[1––0.27(t+1.47)]2.477(2=0.935)。河蚬的壳长和体重生长曲线如图9和图10所示, 可以看出, 随着时间增长, 河蚬壳长和体重不断增加并趋于最大值。
对壳长生长方程分别求一阶导数和二阶导数, 分别得到河蚬壳长的生长速度和生长加速度方程: dL/d=11.34×e–0.27(t+1.47)和2L/d2=–3.06×–0.27(t+1.47), 并绘制其壳长生长速度曲线和生长加速度曲线(图11和图12)。可以看出, 河蚬壳长的生长速度和生长加速度曲线没有生长拐点, 壳长的生长速度随年龄增加而逐渐下降, 并逐渐趋于0; 河蚬壳长的生长加速度小于0, 且随年龄增长逐渐上升。
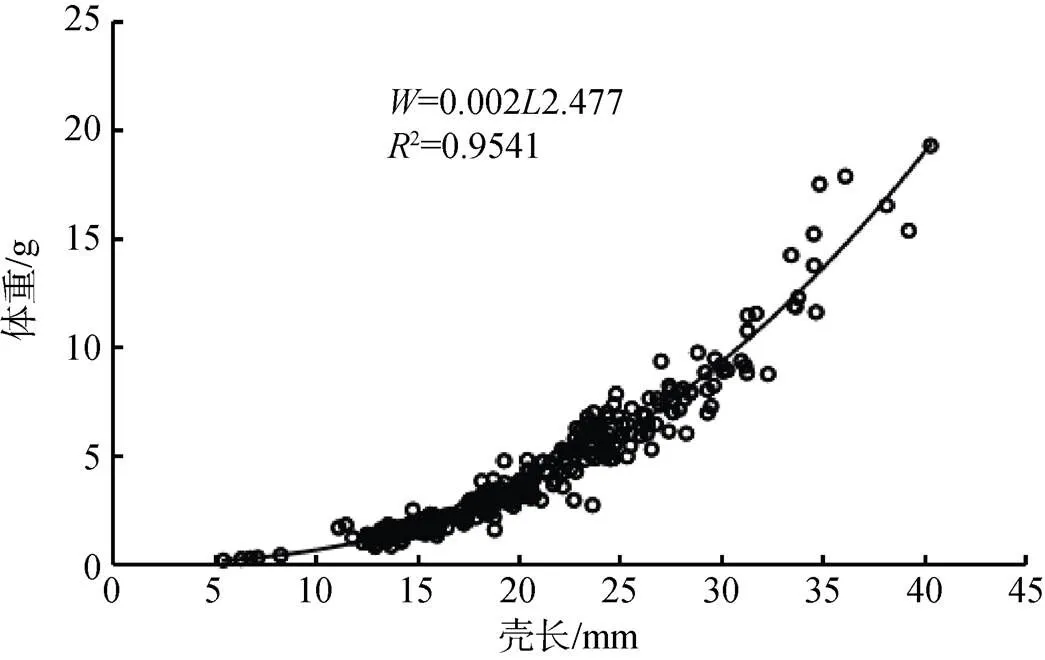
图8 河蚬壳长与体重的关系
Figure 8 Relationships between shell length and body weight of
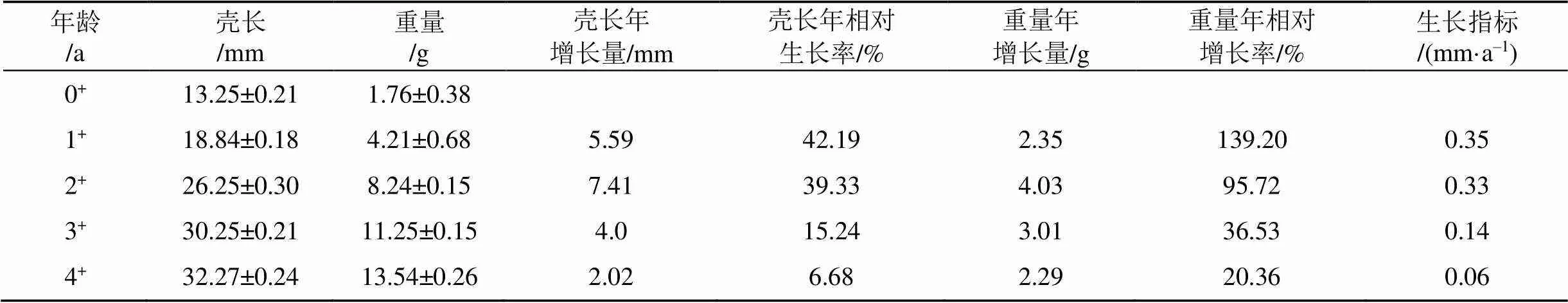
表1 太湖河蚬的生长状况
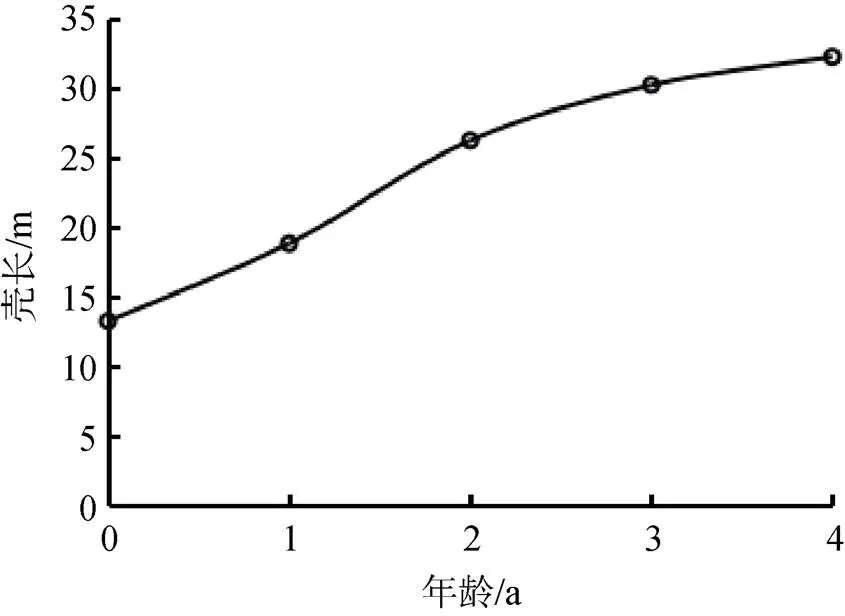
图9 河蚬的壳长生长曲线
Figure 9 Growth curve offor shell length
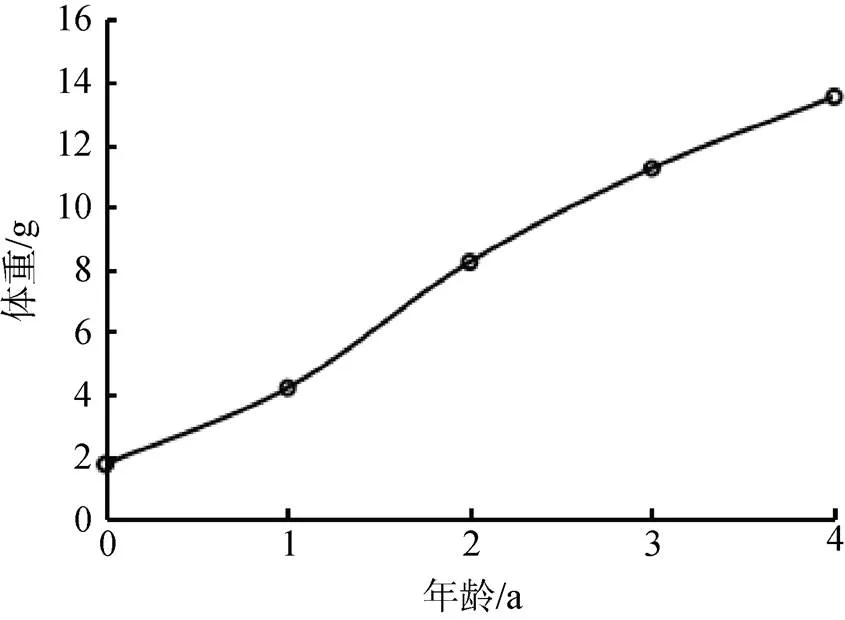
图10 河蚬的重量生长曲线
Figure 10 Growth curve offor wet weight
对体重的生长方程分别求一阶导数和二阶导数, 分别得到体重的生长速度和生长加速度方程:dW/d= 12.89×–0.27(t+1.47)×[1––0.27(t+1.47)]1.47和2W/d2=3.48×–0.27(t+1.47)×[1––0.27(t+1.47)]0.47×[2.47×–0.27(t+1.47)–1], 并绘制其体重生长速度曲线和生长加速度曲线(图13和图14)。可以看出, 河蚬体重生长速度呈现先增加后下降趋势, 生长加速度呈逐渐下降趋势。经计算, 河蚬的体重生长拐点年龄为2.22龄, 对应的拐点壳长和体重分别为26.48 mm和6.16 g。
3 讨论
本研究对太湖河蚬种群的个体大小和年龄进行测量和鉴定, 结果显示, 河蚬的壳长范围为5.45至40.34 mm, 均值为20.17 mm, 其中壳长小于20 mm的个体数量占比为54.23%, 壳长小于生长拐点对应壳长(26.48 mm)的个体数量占比为85.27%, 表明太湖河蚬种群的个体组成呈现出明显的小型化特点。同时发现, 河蚬种群由0+至4+龄共5个年龄组构成, 其中0+至2+龄的个体数量占比达91.85%, 表明太湖河蚬的年龄结构呈现出明显的低龄化特点。由此可以得出, 太湖河蚬种群的个体组成具有典型的小型化、低龄化特征, 与已报道的太湖河蚬种群组成相一致[9, 11]。太湖河蚬个体的小型化、低龄化可能受以下因素影响: (1)过度捕捞。河蚬是重要的水产资源, 经济价值较高, 市场需求大, 驱使渔民对河蚬资源过度捕捞, 进而造成河蚬资源过度开发利用, 且捕捞河蚬的渔具捕捞效率高, 选择性差; (2)环境污染。太湖富营养化严重, 蓝藻水华频发, 污染严重,河蚬对污染物的富集能力强, 导致河蚬直接死亡或生长受到抑制; (3)栖息地丧失。非法采砂、水利工程导致河蚬栖息地丧失及河蚬死亡, 造成河蚬分布范围缩小, 且种群数量在短时间内难以恢复。太湖河蚬资源调查结果表明, 近年来河蚬的分布范围明显缩小, 资源密度和生物量降低, 捕获量急剧下降, 与河蚬个体的小型化、低龄化变化具有密切关系[4, 9, 11-12]。除太湖外, 我国诸多湖泊(洪泽湖、淀山湖、巢湖、大通湖, 鄱阳湖、洞庭湖)的河蚬个体普遍存在小型化、低龄化现象, 且资源量急剧减少[10, 13, 26-30], 应当引起渔业主管部门的高度重视, 并采取措施加强河蚬资源保护。
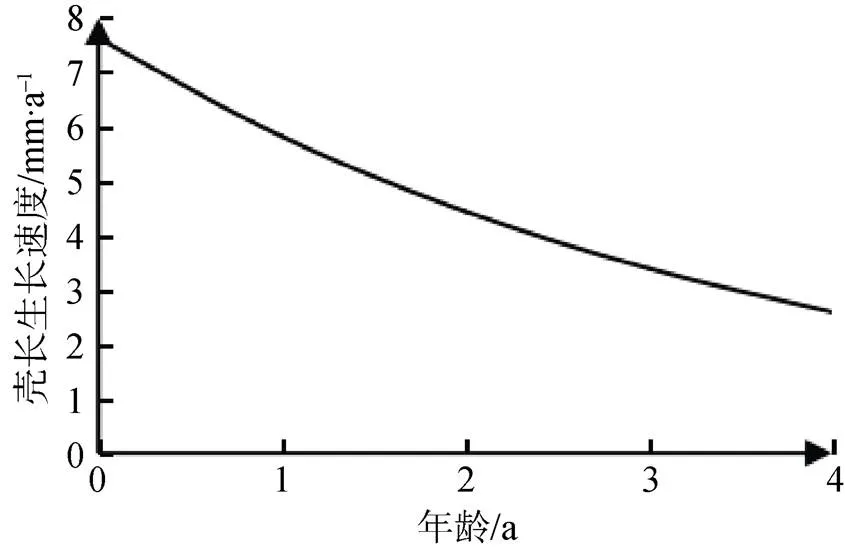
图11 河蚬壳长的生长速度
Figure 11 Grwoth rate curve offor shell length
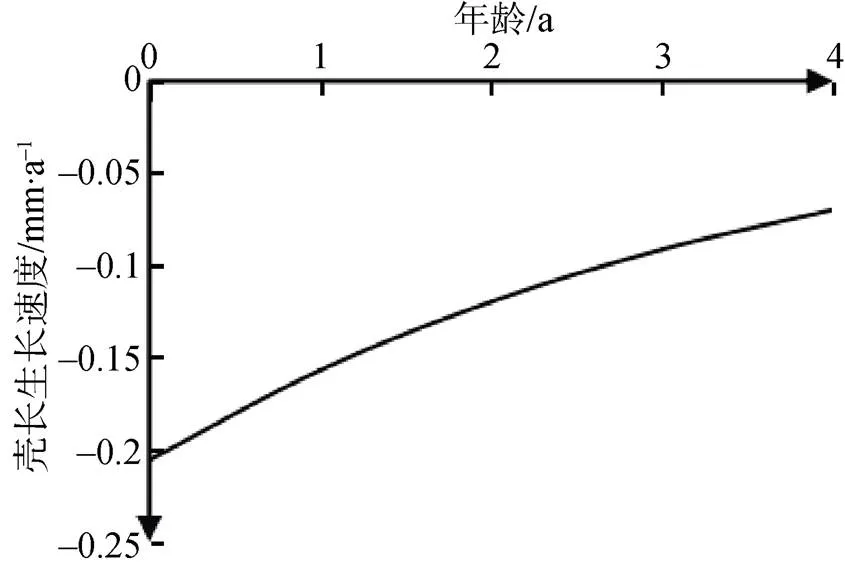
图12 河蚬壳长的生长加速度
Figure 12 Grwoth acceleration curve offor shell length
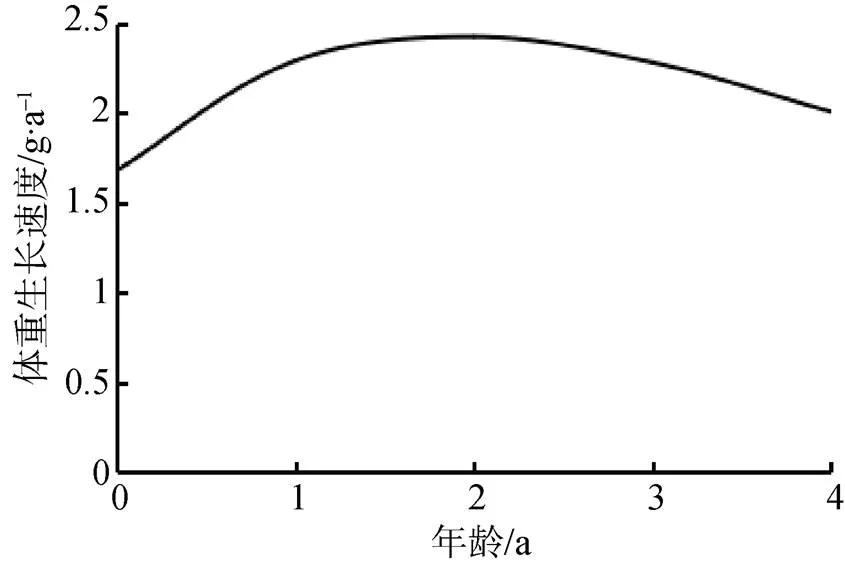
图13 河蚬体重的生长速度
Figure 13 Grwoth rate curve offor weight
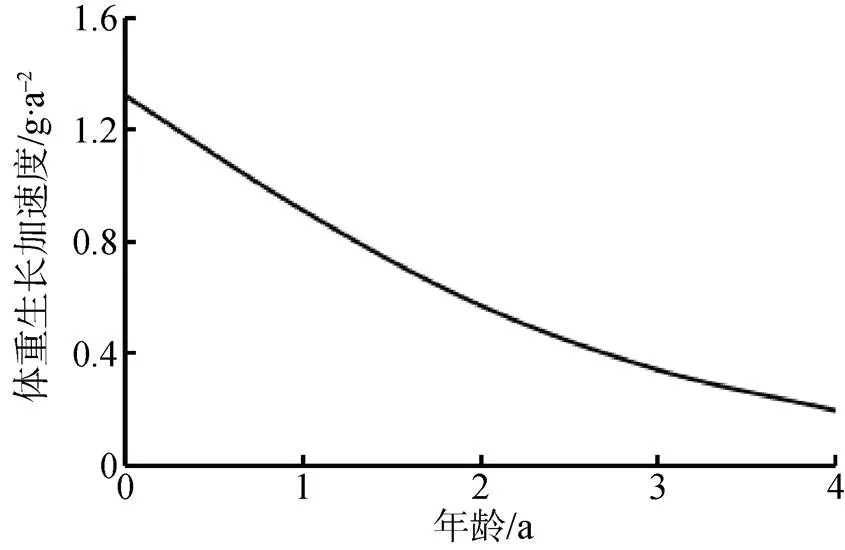
图14 河蚬体重的生长加速度
Figure 14 Grwoth acceleration curve offor weight
本研究结果显示, 太湖河蚬体长和体重的幂指数方程的幂指数为2.477, 且显著小于3, 说明河蚬为异速生长[26]。已有研究表明太湖、洪泽湖、大通湖的河蚬均表为异速生长模式[12, 26-27, 29]。一方面, 受捕捞工具的选择性, 极小的河蚬个体没有被捕获, 而河蚬的体长和体重关系与个体大小相关, 河蚬生长过程经历异速生长-等速生长-异速生长模式[12]。另一方面, 河蚬生长过程中营养条件及环境因子的剧烈波动也是导致河蚬异速增长的重要原因[31]。其次, 河蚬繁殖也会影响其生长模式, 这是因为在河蚬性腺组织发育的不同阶段肥其满度会发生显著变化[32-33]。研究表明, 太湖河蚬繁殖季节主要集中在5至7月份, 性成熟最小规格为1.2cm, 处于繁殖季节河蚬体重增长速率高于非繁殖季节河蚬, 未性成熟个体生长速率大于性成熟个体[9, 12]。根据壳长和体重年相对生长率和生长指标判断, 0+至2+龄河蚬的生长率和生长指标最大, 指示河蚬处于快速生长期; 2+至3+龄河蚬的生长率和生长指标呈小幅下降趋势, 指示河蚬进入稳定生长期; 3+至4+龄河蚬的生长率和生长指标急剧下降, 指示河蚬进入衰老期, 这与洪泽湖河蚬具有相似的生长规律[26]。有研究表明, 河蚬生长速率的变化与个体大小密切相关, 且随着体重的增加而降低, 河蚬幼年的新陈代谢强度大于老年[34]。
迄今, Von Bertalanffy生长方程是是渔业资源管理及评估中应用最为广泛的模型, 基于该模型可以估测鱼类的生长拐点及理论最大体长和体重, 为渔业资源保护和生产提供指导[35]。同样, Von Bertalanffy生长方程也被广泛用于描述贝类生长规律。比如, 利用该方程获得了长牡蛎()的生长拐点为5月龄[36], 洪泽湖河蚬的生长拐点为2.73龄, 相对应的壳长和体重分别为26.28 mm和6.69 g[26]。本研究通过对河蚬的体重生长方程求一阶和二阶导数, 获得河蚬的生长拐点年龄为2.22 龄, 相对应的壳长和体重分别为26.48 mm和6.16 g, 这为制定太湖河蚬的开捕规格提供了重要参考。有研究指出, 太湖河蚬长至1.2 cm即可达到性成熟, 据此制定的太湖商品蚬的规格仅为1.5 cm, 明显小于其生长拐点对应的大小[9]。因此, 基于河蚬发育阶段制定起捕规格不利于河蚬资源保护。另外, 有研究表明不同年龄阶段的河蚬在营养成分上也存在明显差异, 高龄河蚬的含肉量、蛋白质含量以及必需氨基酸含量等重要营养指标均相对较高[37]。综合考虑, 太湖河蚬的起捕规格应增加至26 mm, 这样既可以充分利用河蚬的快速生长期, 发挥其生态作用, 也有利于保护河蚬资源, 并提高河蚬的营养价值。
4 结论
(1)太湖河蚬的理论最大壳长为41.99 mm, 理论最大重量为19.28 g。河蚬的壳长范围为5.45—40.34 mm, 均值为(20.17±0.60) mm, 体质量范围为0.15—19.27 g, 均值为(4.09±0.32) g。河蚬种群由0+至4+龄共5个年龄组构成, 太湖河蚬的种群组成具有明显的小型化、低龄化特点。
(2)太湖河蚬的体长与体重的幂指数关系为= 2.0×10–32.477, 生长为异速增长模式, 且具有明显的阶段性, 其中0+至2+龄处于快速生长期, 2+至3+龄处于稳定生长期, 3+龄至4+龄处于衰老期。
(3)太湖河蚬的壳长生长方程为L= 41.99× [1––0.27(t+1.47)], 体重生长方程为W= 19.28×[1––0.27(t+1.47)]2.477, 体重生长拐点年龄为2.22龄, 对应的壳长和体重分别为26.48 mm和6.16 g, 河蚬的起捕规格应不低于26 mm。
[1] 刘月英, 张文珍, 王跃先. 中国经济动物志: 淡水软体动物[M]. 北京: 科学出版社, 1979: 119–123.
[2] VAUGHN C C, HAKENKAMP C C. The functional role of burrowing bivalves in freshwater ecosystems[J]. Freshwater Biology, 2001, 46(11): 1431–1446.
[3] ILARRI M I, FREITAS F, COSTA-DIAS S, ANTUNES C, et al. Associated macrozoobenthos with the invasive Asian clam[J]. Journal of Sea Research, 2012, 72: 113–120.
[4] 许浩, 蔡永久, 汤祥明, 等. 太湖大型底栖动物群落结构与水环境生物评价[J]. 湖泊科学, 2015, 27(5): 840–852.
[5] 朱利明, 肖文胜, 周东, 等. 淀山湖大型底栖动物群落结构及其与环境因子的关系[J]. 水生态学杂志, 2019, 40(2): 55–65.
[6] 庄平, 宋超, 章龙珍. 长江口河蚬营养成分的分析与评价[J]. 营养学报, 2009, 31(3): 304–306.
[7] 王宇, 刘东红. 河蚬功能性成分的研究现状与展望[J].食品与发酵工业, 2010, 36(6): 122–124.
[8] 王春, 张玮伯, 孙卫军, 等. 河蚬营养成分的提取及其生理功能研究进展[J]. 食品安全质量检测学报, 2021, 12(1): 242–248.
[9] 曹文明, 周刚, 盛建明, 等. 太湖河蚬资源现状及演变[J]. 南京林业大学学报: 自然科学版, 2000, 24(S1): 125–128.
[10] 邓道贵, 李洪远, 胡万明, 等. 巢湖富营养化对河蚬和环棱螺分布及种群密度影响[J]. 应用生态学报, 2005, 16(8): 1502–1506.
[11] 蔡永久, 龚志军, 秦伯强. 太湖软体动物现存量及空间分布格局(2006—2007年)[J]. 湖泊科学, 2009, 21(5): 713–719.
[12] 蔡炜, 蔡永久, 龚志军, 等. 太湖河蚬时空格局[J]. 湖泊科学, 2010, 22(5): 714–722.
[13] 陈彦, 戴小杰, 田思泉, 等. 上海淀山湖内河蚬的分布与种群生长的初步研究[J]. 上海海洋大学学报, 2013, 22(1): 81–87.
[14] 王庆志, 常亚青. 大洋河河蚬的繁殖生物学研究[J].大连水产学院学报, 2010, 25(1): 8–13.
[15] 赵雪琳, 郑小东, 孙同秋, 等. 黄河三角洲河蚬()的繁殖生物学研究[J]. 海洋与湖沼, 2012, 43 (5): 1008–1015.
[16] 邱昕晔, 俞爽, 刘红玲. 河蚬()在生态毒理学研究中的应用与评价[J]. 生态毒理学报, 2016, 11(1): 80–93.
[17] 丁怀宇, 姜虎成, 冯建彬, 等. 河蚬微卫星引物筛选及洪泽湖野生群体遗传结构分析[J]. 水产学报, 2011, 35(11): 1624–1632.
[18] 李大命, 张彤晴, 唐晟凯, 等. 基于线粒体COI序列的洪泽湖河蚬()遗传多样性和种群结构分析[J]. 海洋 与湖沼, 2015, 11(6): 1339–1346.
[19] 李德亮, 张婷, 肖调义, 等. 湖南省大通湖河蚬()现存量及其时空分布[J]. 湖泊科学, 2013, 25(5): 743–748.
[20] 张彤晴, 唐晟凯, 李大命, 等. 洪泽湖河蚬空间分布及资源量研究[J]. 江苏农业科学, 2017, 45(20): 180–187.
[21] 朱小龙, 谷娇, 靳辉, 等. 太湖河蚬()对富营养水体水质的改善作用[J]. 湖泊科学, 2015, 27(3): 486–492.
[22] SOUSA R, FREIRE R, RUFINO M, et al. Genetic and shell morphological variability of the invasive bivalve(Müller, 1774) in two Portuguese estuaries[J]. Estuarine, Coastal and Shelf Science, 2007, 74 (1-2): 166–174.
[23] 蔡英亚, 张英, 魏若飞. 贝类学概论[M]. 上海: 科学技术出版社, 1995: 193–194.
[24] 陈大刚. 渔业资源生物学[M]. 北京: 中国农业出版社, 1997: 1–100.
[25] 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995: 18–48.
[26] 刘燕山, 张彤晴, 唐晟凯, 等. 洪泽湖河蚬种群生长方程估算及其应用[J]. 水生态学杂志, 2017, 38(4): 82–89.
[27] 毕婷婷, 侯刚, 张胜宇, 等. 基于壳长频率分析的洪泽湖河蚬渔获物年龄结构、生长与死亡参数估计[J]. 水生生物学报, 2014, 38(4): 797–800.
[28] 刘勇江. 鄱阳湖淡水双壳类的资源状况及优势种繁殖特性[D] . 南昌: 南昌大学, 2007: 1–67.
[29] 张健. 大通湖河蚬的周年生产量及繁殖特性研究[D]. 长沙: 湖南农业大学, 2016: 1–51.
[30] 王丑明, 张屹, 石慧华, 等. 洞庭湖大型底栖动物群落结构和水质评价[J]. 湖泊科学, 2016, 28(2): 395–404.
[31] 李伟. 河蚬胁迫对太湖河蚬生长的影响[D]. 重庆: 重庆三峡学院, 2020: 1–43.
[32] 王庆志, 常亚青. 大洋河河蚬的繁殖生物学研究[J]. 大连水产学院学报, 2010, 25(1): 8–13.
[33] 吴丛迪, 张建琦, 夏思宇, 等. 洪泽湖河蚬繁殖高峰期的生物学研究[J]. 基因组学与应用生物学, 2020, 39(5): 2028–2032.
[34] 刘其根, 沈和定, 周洪琪, 等. 河蚬的耗氧率和排氨率[J]. 上海水产大学学报, 1999, 8(4): 298–302.
[35] 李大命, 唐晟凯, 刘燕山, 等. 滆湖鲢、鳙种群生长特征及起捕规格[J]. 江苏农业科学, 2021, 49(6): 134–139.
[36] 楼宝. 太平洋牡蛎生长特征研究[J]. 浙江海洋学院学报(自然科学版), 2000, 19(3): 303–306.
[37] 唐晟凯, 张彤晴, 崔晓翠, 等. 洪泽湖不同年龄河蚬营养成分的分析与评价[J]. 江苏农业科学, 2018, 46 (18): 196–199.
Study on the population composition and growth characteristics ofin the Tai Lake
LIU Yang1, 2, LI Daming2,*, LU Jianming3, LIU Yanshan2, TANG Shengkai2, YANG Jiaxin1, ZHANG Tongqing2
1. College of Marine Science and Engineering , Nanjing Normal University, Nanjing 210023, China 2. Freshwater Fisheries Research Institute of Jiangsu Province, Key Laboratory of Fisheries Resources and Environment in Inland water, Nanjing 210017, China 3. Taihu Lake Fisheries Administration Committee Office of Jiangsu Province, Suzhou 215104, China
In order to understand population size composition, age structure and growth characteristics ofin Tai Lake, 638 individuals ofcollected from six sampling sites in Tai Lake were analyzed. The shell length, shell width, shell heigth and wet weight ofwere measured and the ages of all individuals were determined. Furthermore, the growth equations of shell length and wet weight ofwere established. The research results are as follows.(1) The shell length, shell width, shell height and wet weight ofranged from 5.45 to 40.34 mm, from 6.68 to 24.89 mm, from 7.15 to 37.41 mm and from 0.15 to 19.27 g, respectively, the corresponding average values were (20.17±0.60) mm, (13.57±0.33) mm, (19.13±0.53) mm and (4.09±0.32) g, respectively. (2) Thepopulation consisted of five age groups (0+- 4+), in which the number of individuals aging from 0+to 2+yr accounted for 91.85%, and the number of individuals aging from 3+to 4+yr accounted for 8.15%. (3) The relationship between shell length and wet weight was expressed as= 2.0×10–32.477(2=0.954) which indicated thatgrew at an allometric rate. (4)The relationships between shell length and age were described by Von Bertalanffy equations:t=41.99×[1–e–0.27(t+1.47)], and the relationships between wet weight and age were described by Von Bertalanffy equations:t=19.28×[1–e–0.27(t+1.47)]2.477. The growth inflection point ofoccurred at age 2.22 yr, with the corresponding shell length of 26.48 mm and wet weight of 6.16 g. The present results suggested that small size and low age individuals dominated thepopulation and the intensity of resource utilization was too high. Therefore, it is urgent to take measures to protect the natural resources ofin Tai Lake.
; population composition; age structure; growth equation; Tai Lake
10.14108/j.cnki.1008-8873.2024.01.010
Q112
A
1008-8873(2024)01-081-07
2021-08-03;
2021-12-10基金项目:江苏省自然科学基金面上项目(BK20191488); 江苏省水生生物资源重大专项(ZYHB16-3); 2019年淡水渔业资源监测(2019-SJ-018-2)
刘洋(1996—), 男, 江苏靖江人, 硕士研究生, 主要从事底栖动物研究, E-mail: ly964394951@163.com
通信作者:李大命, 男, 博士, 研究员, 主要从事渔业资源资源监测与评估研究, E-mail: ldm8212@126.com
刘洋, 李大命, 陆建明, 等. 太湖河蚬种群组成及生长特征研究[J]. 生态科学, 2024, 43(1): 81–87.
LIU Yang, LI Daming, LU Jianming, et al. Study on the population composition and growth characteristics ofin the Tai Lake[J]. Ecological Science, 2024, 43(1): 81–87.
猜你喜欢
今日农业(2022年15期)2022-09-20
小哥白尼(野生动物)(2021年9期)2022-01-17
数学小灵通(1-2年级)(2020年6期)2020-06-24
红土地(2018年7期)2018-09-26
玩具世界(2017年4期)2017-07-21
数学小灵通·3-4年级(2017年5期)2017-06-05
宝藏(2017年2期)2017-03-20
中亚信息(2016年3期)2016-12-01
小学生作文(中高年级适用)(2016年3期)2016-11-11
金融经济(2015年16期)2015-08-21
