
怒江干热河谷木棉种子大小的变异格局
2024-04-28 05:10陈凯杨艳徐玲赵永福张宏玉
生态科学 2024年1期
陈凯, 杨艳, 徐玲, 赵永福, 张宏玉
怒江干热河谷木棉种子大小的变异格局
陈凯1, *, 杨艳2, 徐玲1, 赵永福1, 张宏玉1
1. 保山学院资源环境学院, 保山 678000 2. 保山市质量技术监督综合检测中心, 保山 678000
为揭示怒江干热河谷木棉种子大小的变异格局, 在该气候区的不同生境采集木棉种子, 测定种子生物量、直径用于量化种子大小, 并借助回归模型分析种子大小随环境梯度的变化。结果显示: (1)在怒江干热河谷, 木棉种子大小呈正态分布; (2)在不同干热区, 种子大小依次为轻度干热区﹥中度干热区﹥重度干热区; (3)木棉种子随海拔升高而呈变小的趋势; (4)木棉种子大小的变异性随海拔(或干旱强度)上升不断减小。结论表明, 木棉种子大小受到水热条件的选择压力, 在稳定的水热环境中木棉种子更大, 其大小的变异性也更高。
怒江干热河谷; 木棉; 种子大小变异; 干热区; 海拔
0 前言
怒江干热河谷是横断山区人口聚集与农业发展中心之一, 对滇西南社会经济发展具有重要意义[1]。受喜马拉雅隆升、焚风效应、人类干扰等因素的综合影响, 怒江干热河谷的气候较同纬度地区温度高、降雨少、湿度低, 因而植被稀疏、水土流失严重, 是典型的生态脆弱区[2]。
种子大小代表植物为子代储存的能量多少, 是关键植物功能性状[3]。大种子能适应竞争激烈的稳定环境, 而小种子在波动生境中具有优势[4], 因此种子大小变异反映植物在不同生境下的生态适应策略, 也是植物多样性形成与生态系统功能维持的重要驱动因子[5]。已有的证据表明, 植物种子大小存在巨大变异。在不同植物间种子大小变异可达11个数量级, 比如: 最小的兰科植物种子为1×10–7g, 而双椰子()的种子重达27 kg[6]。种子大小也存在一定程度的种内变异, 如葱芥()的种子大小可相差8倍[7], 万寿菊()的种子大小变异可达29倍[8]。值得注意的是, 种子大小变异可呈现一定的环境梯度格局[9], 比如: 在青藏高原随着海拔升高, 植物种子逐渐变小[10]; 在北美洲的针叶林, 裸子植物种子随降雨量增加呈增大的趋势[11]。尽管已有大量种子大小变异的研究, 但这些研究主要关注森林、草原等典型生态系统, 而极端生境(如干热河谷)下种子大小变异的研究较少。
木棉()是怒江干热河谷最高大的乔木树种, 对该区域的水土保持具有重要作用[12]。木棉在怒江干热河谷广泛分布, 从怒江江畔至山腰的咖啡地都散布有野生居群。目前发现木棉种子大小存在较大变异, 这种变异进一步影响种子萌发方式, 对种群天然更新和世代维持具有重要意义[13-14]。然而, 目前尚不清楚, 怒江干热河谷木棉种子大小变异呈现何种格局?本研究拟在怒江干热河谷进行木棉种子采集, 探究种子大小在不同生境的变异规律, 以期揭示木棉种子的生态适应策略。
1 材料与方法
1.1 怒江干热河谷区划
怒江干热河谷位于E98°50′—98°55′、N24°40′—25°50′(图1), 海拔跨度约为400—1500 m, 全年干热少雨, 是中国西南乃至全球独特的气候景观。为将怒江干热河谷按干旱程度进一步区划, 从WorldClim (v1.4, http: //www.worldclim.org/)下载怒江干热河谷降雨量图层, 根据该区域年均降雨量的第66(1477 mm)、33(1246 mm)百分位数, 将河谷划分为: 轻度(降雨量> 1477 mm)、中度(1246 mm ≤降雨量≤ 1477 mm)及重度(降雨量< 1246 mm)干热区。
1.2 木棉种子采集与处理
2020年4—5月, 分别在怒江干热河谷的轻度、中度及重度干热区由南向北采集木棉种子, 每间隔约20 km设一取样点(图1), 每个取样点采集3个木棉居群, 每个居群随机选取5株个体, 每株摘取2颗成熟的果实(尖端自然炸裂), 同时记录采样点的海拔。最终在轻度、中度和重度干热区分别采集到15份、12份、9份木棉果实。果实摘取后放入布袋中, 于当天带回实验室, 清理掉果皮、绒毛及杂物后将种子置于20 ℃、相对湿度15%的干燥间30 d。
1.3 种子大小测定
种子大小包括种子的质量(生物量)与体积[6, 10]。用万分之一的电子天平称量木棉种子生物量, 每个居群随机选取35粒种子, 逐一对每粒种子称重。木棉种子为圆形, 可用直径衡量其体积; 用数显游标卡尺测量种子直径, 每个居群随机测定35粒。
1.4 计算变异系数
用标准差对平均数的比值表示变异系数[14]。分别计算每个居群种子生物量、直径的变异系数。
1.5 数据分析
用单因素方差分析(One-way ANOVA)分别比较不同干热区种子生物量、直径大小及变异系数的差异; 若方差分析显著, 则进一步用忠实显著性差异(Tukey’s HSD)检验法进行多重比较。用简单线性回归(Simple Linear Regression)分析种子生物量、直径随海拔的变化规律。数据分析与作图在R 3.6.2中完成[15]。
2 结果与分析
2.1 木棉种子大小的分布
木棉种子生物量最小值、均值和最大值分别为: 0.008 g、0.031 g和0.060 g, 标准差达到0.009 g (图2A); 种子的最小、平均及最大直径分别为: 2.780 mm、3.753 mm及4.830 mm, 标准差达到0.299 mm (图2B)。木棉种子生物量、直径大小都近似正态分布(生物量= 0.980,直径= 0.989; 图2)。
2.2 木棉种子大小在不同干热区的变异格局
单因素方差分析显示, 在轻度、中度及重度干热区之间木棉种子生物量(= 90.590,< 0.001; 图3A)、直径(= 22.320,< 0.001; 图3B)的大小差异都极显著。多重比较的结果表明, 轻度干热区的木棉种子生物量显著大于中度(差值 = 0.004 g,< 0.001; 图3A)、重度(差值 = 0.008 g,< 0.001; 图3A)干热区的种子生物量, 中度干热区的种子生物量显著大于重度干热区(差值 = 0.004 g,< 0.001; 图3A); 同样轻度与中度干热区的木棉种子直径显著大于重度干热区的种子直径(差值 = 0.131 mm,< 0.001; 差值 = 0.103 mm,< 0.050; 图3B), 但轻度与中度干热区的种子直径差异不显著(差值 = 0.028 mm,= 0.731; 图3B)。
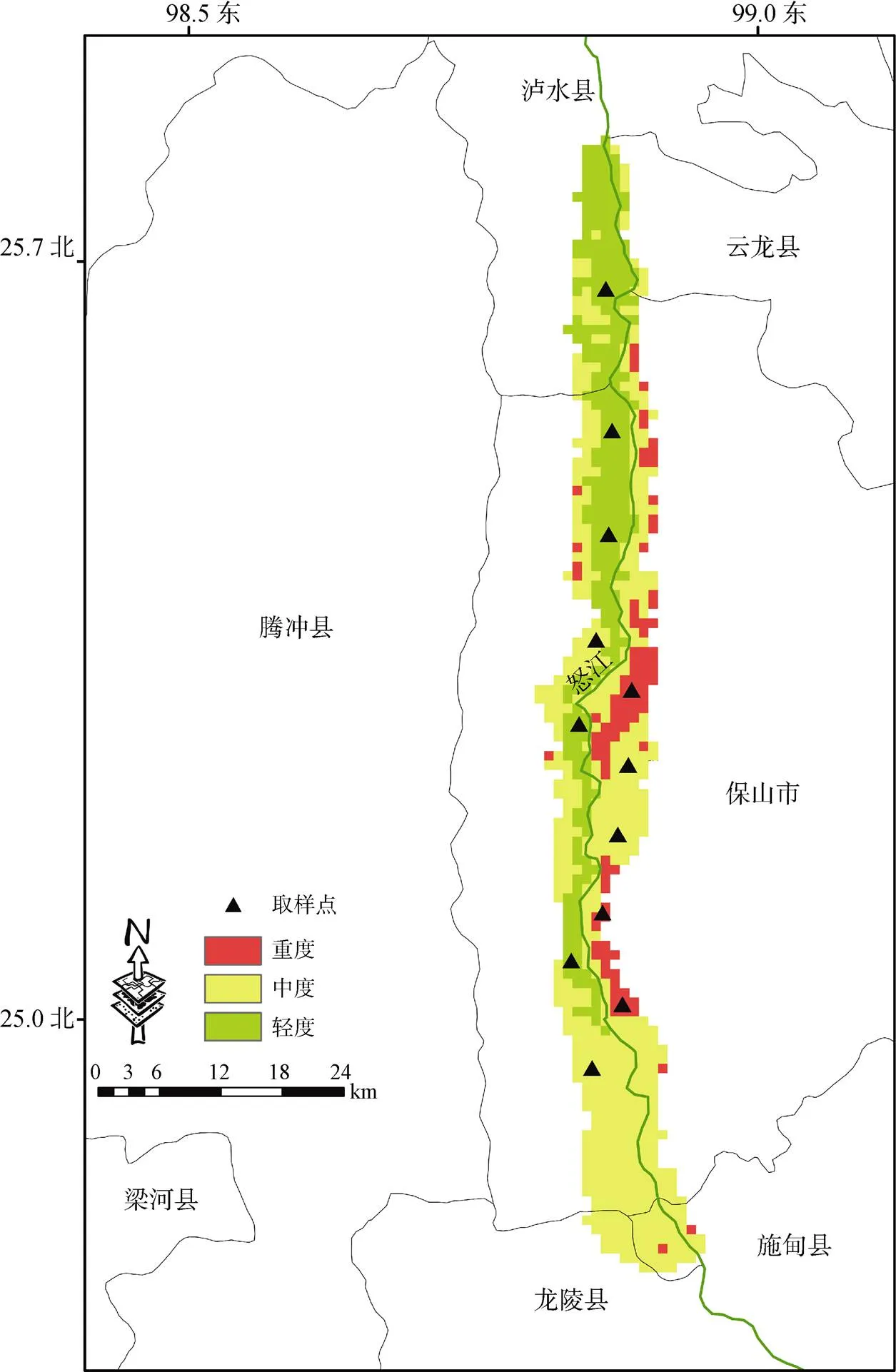
图1 木棉种子的采集样点(黑色三角形)
Figure 1 Location of populations ofseeds sampled in the study (black triangles)
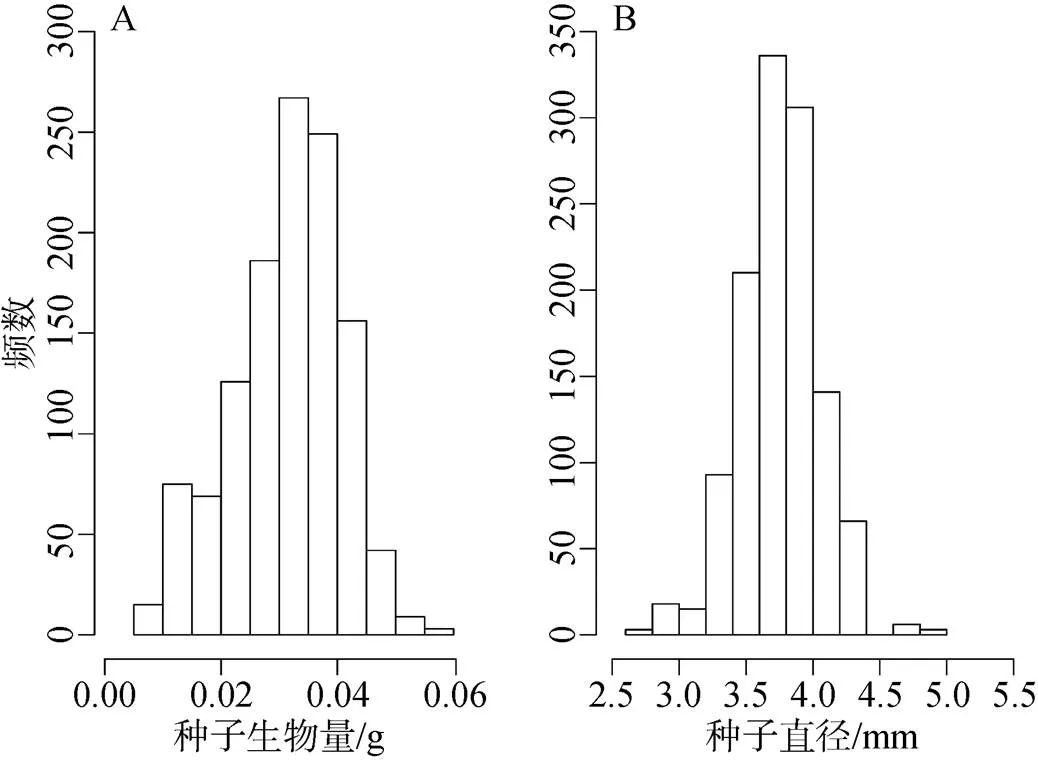
图2 木棉种子生物量(A)与直径(B)的分布
Figure 2 Distributions of seed mass (A) and diameter (B) in
轻度、中度及重度干热区之间木棉种子生物量的变异系数差异极显著(= 23.250,< 0.001; 图4A), 其中轻度、中度干热区的种子生物量变异系数显著大于重度干热区种子生物量的变异系数(差值 = 0.175,< 0.001; 差值 = 0.091,< 0.050; 图4A), 但轻度与中度干热区之间生物量变异系数差异不显著(差值 = 0.084,= 0.053; 图4A)。种子直径的变异系数在轻度、中度及重度干热区之间差异不显著(= 3.852,= 0.058; 图4B)。

注: 相同的小写字母表示多重比较(Tukey’s HSD)检验出两两差异不显著。
Figure 3 Comparisons of seed mass (A) and diameter (B) amongpopulations distributed at different dry-hot regions
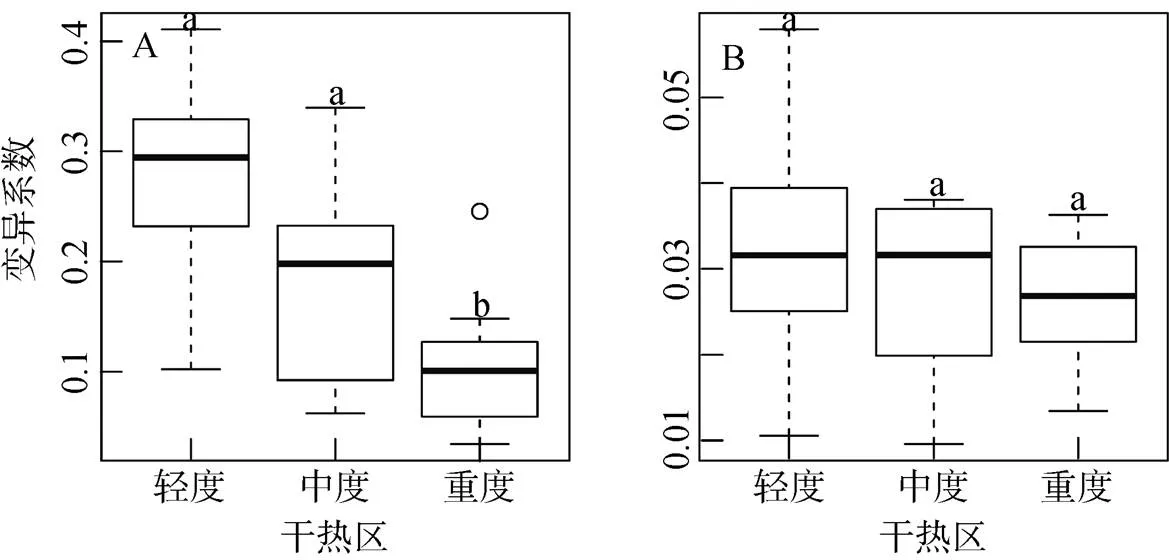
注: 相同的小写字母表示多重比较(Tukey’s HSD)检验出两两差异不显著。
Figure 4 Comparisons of seed mass (A) and diameter (B) variation coefficients amongpopulations distributed at different dry-hot regions
2.3 木棉种子大小沿海拔梯度的变异格局
简单线性回归分析显示, 木棉种子生物量与海拔存在显著的直线回归关系(回归系数 = –1.475×10–5,< 0.050), 海拔能解释36.5%的种子生物量变异(图5A); 但木棉种子直径大小随海拔变化不显著(回归系数 = –1.974×10–4,= 0.234; 图5B)。
随着海拔升高, 木棉种子生物量的变异系数显著减小(回归系数 = –3.293×10–4,< 0.001), 海拔能解释38.9%的变异系数变化(图6A); 而种子直径的变异系数随海拔上升也呈降低的趋势(回归系数 = –1.581×10–5,= 0.058; 图6B)。
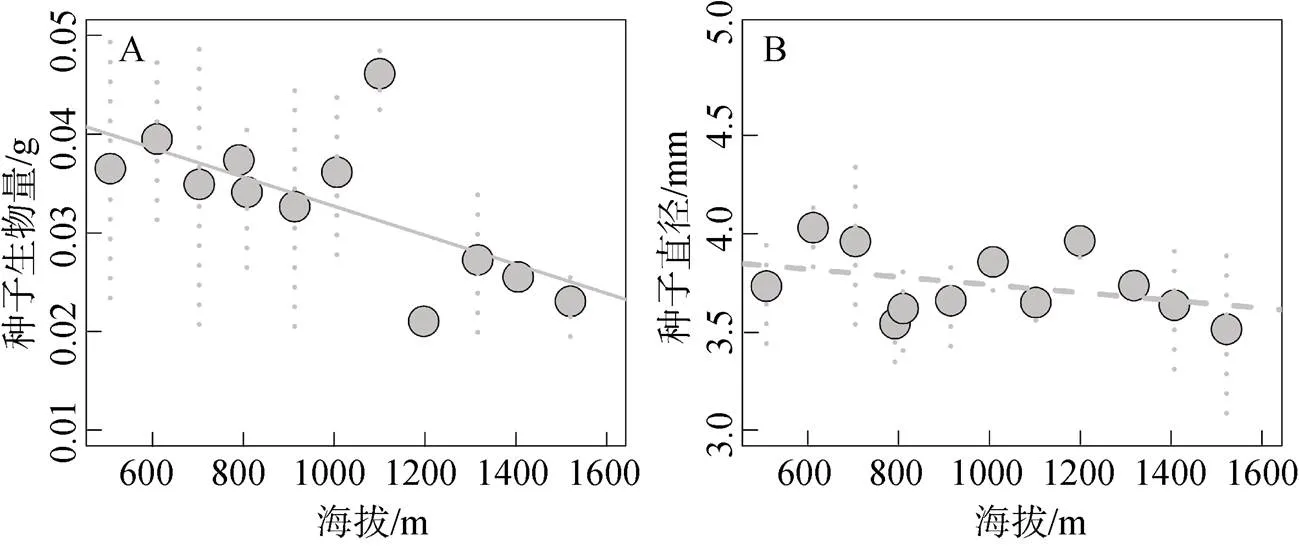
图5 木棉种子生物量(A)与直径(B)大小沿海拔梯度变异
Figure 5 Seed mass (A) and diameter (B) variations inalong elevational gradients
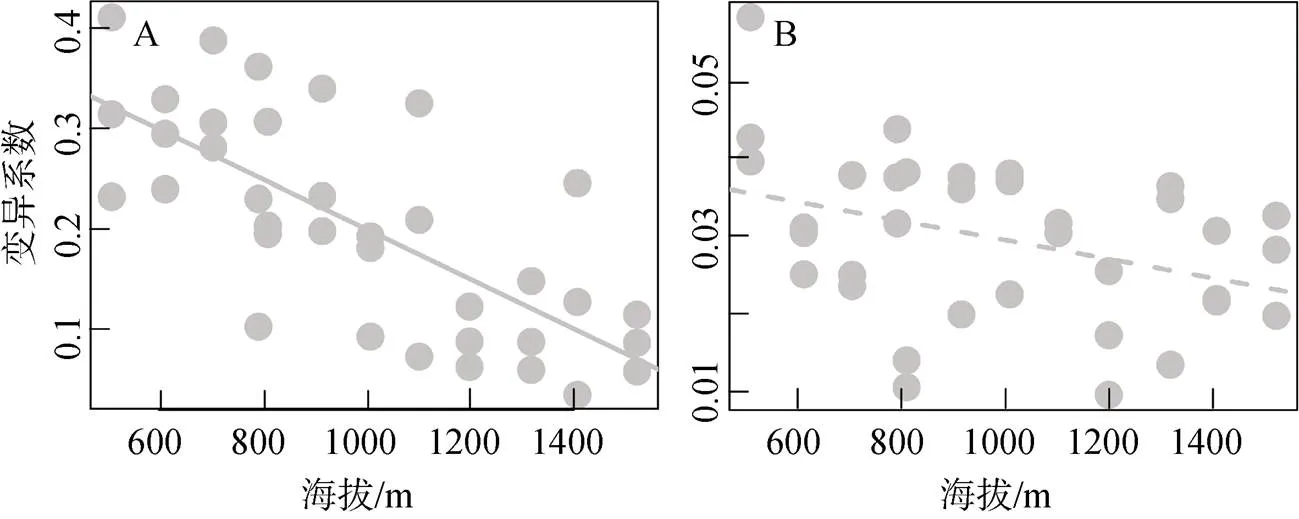
图6 木棉种子生物量(A)与直径(B)的变异系数沿海拔梯度变化
Figure 6 Variation coefficients of seed mass (A) and diameter (B) change inalong elevational gradients
3 讨论
怒江干热河谷的木棉种子大小呈现正态分布, 中等大小的种子数量多, 特别大/小的种子数量少。这与Moles等(2007)在全球种子大数据分析中[16]、Edwards等(2015)对种子生物量统计建模的研究结果一致[17]。种子大小呈正态分布, 暗示着过小或过大的种子受到更大的自然选择压力, 这些种子适合度低, 更可能被自然环境所过滤[18]。种子太小携带的能量过少, 导致幼苗在生活史早期不能获得足够营养物质, 更容易夭折; 而太大的种子不利于扩散(尤其是木棉等靠风力传播的种子)且在土壤种子库中寿命更短, 萌发后幼苗离母株更近, 可能遭受更大的竞争压力。
在怒江干热河谷, 年降雨量相对多的轻度干热区其木棉种子大于中度、重度干热区的种子。水分是干热区植物生长发育的重要限制因子[19], 轻度干热区的木棉较其他分布区的木棉能获得更多水分, 故种子更大。长期遭受水分胁迫的植物可提早繁殖并迅速结束生活史, 而繁殖期的植物处于缺水状态, 会加速果实成熟以逃避胁迫环境[20]; 上述这些响应方式都可能缩短种子发育的周期导致种子变小。Liu等(2013)在加拿大北方针叶林的研究与本研究的结果一致, 他们发现裸子植物的种子生物量随降雨量增加而变大, 降雨能解释近30%的种子大小变异[11]。但Malhado等(2015)在亚马逊热带雨林发现降雨量与植物种子大小无显著相关性, 原因可能是热带雨林全年多雨, 水分并不是植物生长发育的制约因子[21]。总之, 降雨是否影响植物种子大小, 取决于植物所属的生态系统类型。
随着海拔升高, 木棉种子呈不断变小的趋势。对于这种海拔格局, 有两种可能的解释: ①能量限制假说认为, 海拔升高植物由环境中获得的能量减少, 其净初级生产力降低, 因而种子变小[22]; ②虽然怒江干热河谷年均温整体较高, 但江畔与山腰温度相差约6 ℃(海拔上升100 m, 温度下降约0.6 ℃), 木棉生长的最适温度可能与江畔温度更接近, 随着海拔上升, 木棉的生境温度逐渐偏离最适温度, 所以种子呈变小的趋势。然而, 目前种子大小的海拔格局的研究结果存在不一致性。杜鹃属()植物种子随海拔上升而变小[23], 但风毛菊属()的种子随海拔上升而变大[24]。植物种子大小的海拔格局可能与类群的温度生态幅有关, 若海拔升高生境温度偏离植物最适温度, 种子大小与海拔呈负相关, 反之则二者呈正相关。
木棉种子大小的变异性随海拔(或随干旱强度)上升不断减小。与低海拔生境(或轻度干热区)相比, 高海拔生境(或高度干热区)的种子面临更大的自然选择压力, 很多大种子被环境所过滤, 种子大小的取值带谱更窄, 故变异性更小[18]。Májeková等(2016)同样发现, 草本植物在适宜生境中其生物量大小的变异性更高[25]。植物性状的种内变异与塑性基因或整个基因组有关[26], 因而种子大小的变异性可能是环境与遗传背景共同作用的结果, 但目前尚不清楚哪个因子对变异性起主导作用。
4 结论
怒江干热河谷的木棉种子大小呈正态分布, 中等大小的种子适合度最高; 轻度干热区的木棉种子最大, 水分是怒江干热河谷木棉种子大小的重要限制因子; 在怒江干热河谷, 木棉种子大小随海拔升高不断变小; 木棉种子大小的变异性随海拔和干旱强度上升也呈现不断减小的趋势。
[1] TANG Ya, XIE Jiasui, SUN Hui. Revisiting sustainable development of dry valleys in Hengduan Mountains region[J]. Journal of Mountain Science, 2004, 1(1): 38–45.
[2] FANG Shu, ZHAO Yonghua, HAN Lei, et al. Boundaries and characteristics of arid regions in mountain valleys in Southwestern China[J]. Mountain Research and Development, 2018, 38(1): 73–84.
[3] PÉREZ-HARGUINDEGUY N, DÍAZ S, GARNIER E, et al. New handbook for standardised measurement of plant functional traits worldwide[J]. Australian Journal of Botany, 2013, 61(3): 167–234
[4] WESTOBY M. A leaf-height-seed (LHS) plant ecology strategy scheme[J]. Plant and Soil, 1998, 199: 213–227.
[5] CHEN Kai, BURGESS K, YANG Xiangyun, et al. Functional trade-offs and the phylogenetic dispersion of seed traits in a biodiversity hotspot of the Mountains of Southwest China[J]. Ecology and Evolution, 2018, 8(4): 2218–2230.
[6] HARPER J L, LOVELL P H, MOORE K G. The shape and sizes of seeds[J]. Annual Review of Ecology and Systematics, 1970, 1(1): 327–356.
[7] SUSKO D J, LOVETT-DOUST L. Patterns of seed mass variation and their effects on seedling traits in(Brassicaceae)[J]. American Journal of Botany, 2000, 87(1): 56–66.
[8] HOLLAND J N, CHAMBERLAIN S A, WAGUESPACK A M, et al. Effects of pollen load and donor diversity on seed and fruit mass in the,(Cactaceae)[J]. International Journal of Plant Sciences, 2009, 170(4): 467–475.
[9] 于顺利, 陈宏伟, 李晖. 种子重量的生态学研究进展[J]. 植物生态学报, 2007, 31(6): 989–997.
[10] QI Wei, GUO Shuqing, CHEN Xuelin, et al. Disentangling ecological, allometric and evolutionary determinants of the relationship between seed mass and elevation: insights from multiple analyses of 1355 angiosperm species on the eastern Tibetan Plateau[J]. Oikos, 2014, 123(1): 23–32.
[11] LIU Jushan, BAI Yuguang, LAMB E G, et al. Patterns of cross-continental variation in tree seed mass in the Canadian boreal forest[J]. Plos One, 2013, 8(4): e61060.
[12] 孔国辉, 刘世忠, 吴彤, 等. 油页岩废渣场26种木本植物光合作用和生长的差异[J]. 热带亚热带植物学报, 2006, 14(6): 467–476.
[13] 赵高卷, 徐兴良, 马焕成, 等. 红河干热河谷木棉种群的天然更新[J]. 生态学报, 2016, 36(5): 1342–1351.
[14] 高柱, 王小玲, 伍建榕, 等. 西南地区木棉居群种子变异及萌发特性研究[J]. 中南林业科技大学学报, 2015, 35(10): 6–12.
[15] R Development Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, 2006.
[16] MOLES A T, ACKERLY D D, TWEDDLE J C, et al. Global patterns in seed size[J]. Global Ecology and Biogeography, 2007, 16(1): 106–116.
[17] EDWARDS W, MOLES A T, CHONG C. Generalised extreme value distributions provide a natural hypothesis for the shape of seed mass distributions[J]. PLoS One, 2015, 10(4): e0121724.
[18] DIAZ S, KATTGE J, CORNELISSEN J H, et al. The global spectrum of plant form and function[J]. Nature, 2016, 529(7585): 167–171.
[19] 闫帮国, 刘刚才, 樊博, 等. 干热河谷植物化学计量特征与生物量之间的关系[J]. 植物生态学报, 2015, 39(8): 807–815.
[20] GUPTA A, RICO-MEDINA A, CAO-DELGADO A I. The physiology of plant responses to drought[J]. Science, 2020, 368(6488): 266–269.
[21] MALHADO A C M, OLIVEIRA-NETO J A, STROPP J, et al. Climatological correlates of seed size in Amazonian forest trees[J]. Journal of Vegetation Science, 2015, 26(5): 956–963.
[22] Baker H G. Seed weight in relation to environmental conditions in California[J]. Ecology, 1972, 53(6): 997–1010.
[23] WANG Yongji, WANG Jianjian, LAI Liming, et al. Geographic variation in seed traits within and among forty-two species of(Ericaceae) on the Tibetan plateau: relationships with altitude, habitat, plant height, and phylogeny[J]. Ecology and Evolution, 2014, 4(10): 1913–1923.
[24] WU Gaolin, TIAN Fuping, REN Guohua, et al. Seed mass increase along altitude within four(Asteraceae) species in Tibetan plateau[J]. Polish Journal of Ecology, 2011, 59(3): 617–622.
[25] MÁJEKOVÁ M, BELLO F D, DOLEŽAL J, et al. Plant functional traits as determinants of population stability[J]. Ecology, 2016, 95(9): 2369–2374.
[26] NICOTRA A B, ATKIN O K, BONSER S P, et al. Plant phenotypic plasticity in a changing climate[J]. Trends in Plant Science, 2010, 15(12): 684–692.
Variation patterns ofseed size in Nujiang Dry-hot Valley
CHEN Kai1*, YANG Yan2, XU Ling1, ZHAO Yongfu1, ZHANG Hongyu1
1. School of Environment and Resources Sciences, Baoshan University, Baoshan 678000, China 2. Baoshan Comprehensive Inspection Center of Quality and Technical Supervision, Baoshan 678000, China
In order to reveal the variation patterns of(Bombacaceae) seed size in Nujiang Dry-hot Valley,seeds were collected from different habitats of Nujiang Dry-hot Valley. Then seed biomass and diameter were measured to quantify seed size. The regression models were used to analyze seed size variations along environmental gradients in Nujiang Dry-hot Valley. The results are as follows. (1)seed size (including mass and diameter) showed normal distributions in Nujiang Dry-hot Valley. (2)Seed size had significant differences among three different dry-hot regions, with values of mild dry-hot region > moderate dry-hot region > severe dry-hot region. (3) Seed size decreased with increasing elevation. (4) Seed size variability also decreased with increasing elevation and drought intensity. These results suggest thatseeds are under hydrothermal selective pressures, and populations ofdistributed in stable hydrothermal habits produce larger seeds and also have higher seed size variability.
Nujiang Dry-hot Valley;; seed size variation; dry-hot region; elevation
10.14108/j.cnki.1008-8873.2024.01.011
Q948.1
A
1008-8873(2024)01-088-06
2021-08-06;
2021-10-07
国家自然科学地区基金项目(32160078); 云南省科技厅地方高校联合专项(202001BA070001-091); 国家自然科学地区基金项目(32060638)
陈凯(1985—), 男, 湖南邵阳人, 博士, 副教授, 主要从事植物生态学研究, E-mail: kchen1985@163.com
通信作者:陈凯
陈凯, 杨艳, 徐玲, 等. 怒江干热河谷木棉种子大小的变异格局[J]. 生态科学, 2024, 43(1): 88–93.
CHEN Kai, YANG Yan, XU Ling, et al. Variation patterns ofseed size in Nujiang Dry-hot Valley[J]. Ecological Science, 2024, 43(1): 88–93.
猜你喜欢
红领巾·萌芽(2023年8期)2023-07-21
小哥白尼(趣味科学)(2021年5期)2021-08-13
云南画报(2021年4期)2021-07-22
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
歌海(2021年6期)2021-02-01
幼儿园(2020年15期)2020-11-24
艺术家(2020年5期)2020-07-09
Zoological Research(2019年3期)2019-06-28
海峡姐妹(2019年5期)2019-06-18
