
毛乌素沙地盐池地区固定沙丘主要固沙灌木的空间分布格局与空间关联性研究
2024-04-28 05:50牛程旭张定海张志山王艳莉
生态科学 2024年1期
牛程旭, 张定海,*, 张志山, 王艳莉
毛乌素沙地盐池地区固定沙丘主要固沙灌木的空间分布格局与空间关联性研究
牛程旭1, 张定海1,*, 张志山2, 王艳莉3
1. 甘肃农业大学理学院数量生物研究所, 兰州 730070 2. 中国科学院西北生态环境资源研究院, 沙坡头沙漠研究试验站, 兰州 730000 3. 甘肃农业大学林学院, 兰州 730070
种群的空间分布格局与关联性的研究对预测种群的演替趋势和植被重建具有重要意义。基于毛乌素沙地固定沙丘上固沙灌木的地理位置和生长发育阶段数据, 采用单变量、双变量成对相关函数和Monte-Carlo拟合检验方法, 对油蒿()、杨柴()和柠条()三种优势种群进行空间分布格局和关联性分析。结果表明: (1)三个种群在小尺度下呈现聚集分布, 大尺度下呈现随机或均匀分布; 杨柴和柠条种群在研究尺度范围内主要表现为聚集分布。(2)种间关系表明: 油蒿和杨柴在小于5 m尺度下负相关, 其他尺度下基本不具有相关性。(3)种群不同生长发育阶段之间的种内关联关系表明: 在小尺度下, 3个种群的种内关系基本呈正相关; 在大尺度下, 油蒿种内关系呈负相关, 杨柴和柠条的种内关系不存在相关性。
毛乌素沙地; 固沙灌木; 点格局分析; 分布格局; 空间关联性
0 前言
自然界中生物个体、种群和群落的生存都需要一定的空间, 不同的生物个体生存所需的空间大小存在一定差异。种群是由个体组成, 个体在空间中的位置或布局, 即为种群的空间分布格局。种群的空间关联性是指不同种群在空间上的相互联结性, 同时也是种群间相互关系的一种表现形式[1–2]。分析种群的空间分布格局与空间关联性对于理解种内和种间的竞争关系、依赖关系、外界环境对种群的干扰、种群的生物学特性以及种群和环境的关系具有十分重要的意义[3–5]。
种群空间分布的类型主要分为均匀分布、随机分布和聚集分布。种群的空间分布格局与空间尺度存在密切的关系, 种群可能在较小的空间尺度下为聚集分布, 但在较大的空间尺度下却有可能呈现出随机分布或均匀分布。种群空间关联性也有3种情形, 空间正相关、空间负相关和空间无相关性[6]。种群空间分布格局与空间关联性关系密切, 聚集分布与空间正相关体现出种群内部互惠依赖的生态关系, 均匀分布与空间负相关表明种群内部存在竞争的生态关系, 随机分布与空间无相关性则反映出种群内部的生态关系不明确[7–8]。
对种群空间分布格局传统的研究方法有: 方差均值比例法[9]、以密度为基础的样方法和以距离为基础的无样方法[10]、频次检验法、分布型指数法和最近邻分析法等[11], 其中以最近邻分析法最为流行。但是, 最近邻分析法只能检验个体分布的随机性, 不能反映种群空间分布格局随着空间尺度变化的情况[12]。总体上, 空间格局传统的分析方法常常限定在特定的空间尺度上进行, 忽略了种群空间分布格局与空间尺度之间的依赖性, 不能全面准确地阐释种群空间分布格局随尺度动态变化的情况[13]。目前, 为突破传统方法的局限性, 已经出现了许多针对不同空间尺度的空间格局研究方法, 如大尺度上的趋势面分析法, 中等尺度上的双向轨迹方差法, 小尺度上的种毗连法等[14]。

1 材料与方法
1.1 研究区概况
毛乌素沙地是中国四大沙地之一, 研究区位于宁夏盐池县毛乌素沙地生态系统国家定位观测研究站(37°27′N, 107°13′E), 平均海拔1550 m, 年均降水量约为291 mm, 80%的降水主要集中在6—9月, 且多为暴雨集中降落, 固沙植被的降水利用率较低[19, 20], 年均温度8.34 ℃[21]。研究区的地形主要是高低起伏的丘陵、滩地、谷地、台地、梁地等, 大部分固定、半固定和流动沙丘主要覆盖在滩地和台地上。主要固沙灌木种群为油蒿(, 缩写为AO)、杨柴(, 缩写为HM)和柠条(, 缩写为CK)等。草本有赖草()、白草()、针茅草()等[22]。
1.2 样地调查
2014年6月沿地形在研究区(37°27′N, 107°13′E)的固定沙丘上设置调查样地, 调查样地面积为40 m×128 m, 样地包含迎风坡、背风坡、丘顶和丘底 4 种沙丘地貌类型。将调查样地划分成4 m×4 m的正方形样方, 调查样方内所有的固沙灌木, 调查指标包括固沙灌木的类型、生长发育阶段、冠幅、株高、固沙灌木所处位置的经纬度和海拔以及样地的土壤水分和表层土壤养分含量。其中, 固沙灌木的位置利用GPS-RTK定位仪(南京鼎吉测绘仪器有限公司, S86T)测量。
1.3 研究方法
1.3.1 种群空间分布格局分析
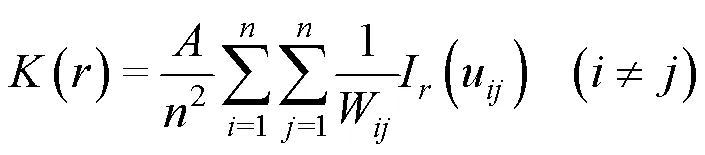
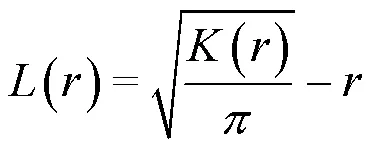
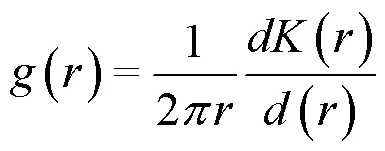
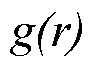
1.3.2 种群空间关联性分析


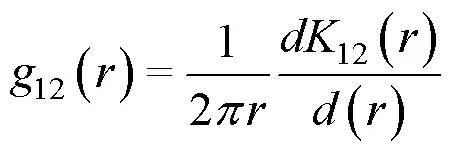
2 结果与分析
2.1 种群的结构组成
研究区固定沙丘调查样地中广泛分布着3个固沙灌木种群, 3个种群共有固沙灌木4098株。其中, 油蒿、杨柴和柠条分别有3682株、369株和48株, 占比分别为89.8%、9%和1.2%。将个体的不同生长发育阶段分为4种类型, 分别记为幼株(简记为Y)、营养株(简记为N)、生殖株(简记为R)和死株(简记为D)。3个种群不同生长发育阶段的个体数量的占种群数量的比例如图1所示, 不同生殖阶段个体的比例关系均表现出相同的种群结构, 即3种群中营养株, 生殖株、幼株和死株的占比依次减少, 营养株和生殖株的总和占到整个种群的90%以上, 但杨柴的生殖株在种群中的占比较少。同时, 需要指出的是, 样地内柠条种群中幼株仅有2株, 没有发现死株, 因此, 在后续的分析中没有分析柠条种群中幼株和死株的空间格局和空间关联性。
2.2 种群的空间分布格局
2.3 种群的空间关联性
2.3.1 不同种群的种间关联性

2.3.2 种内不同发育阶段的空间关联性
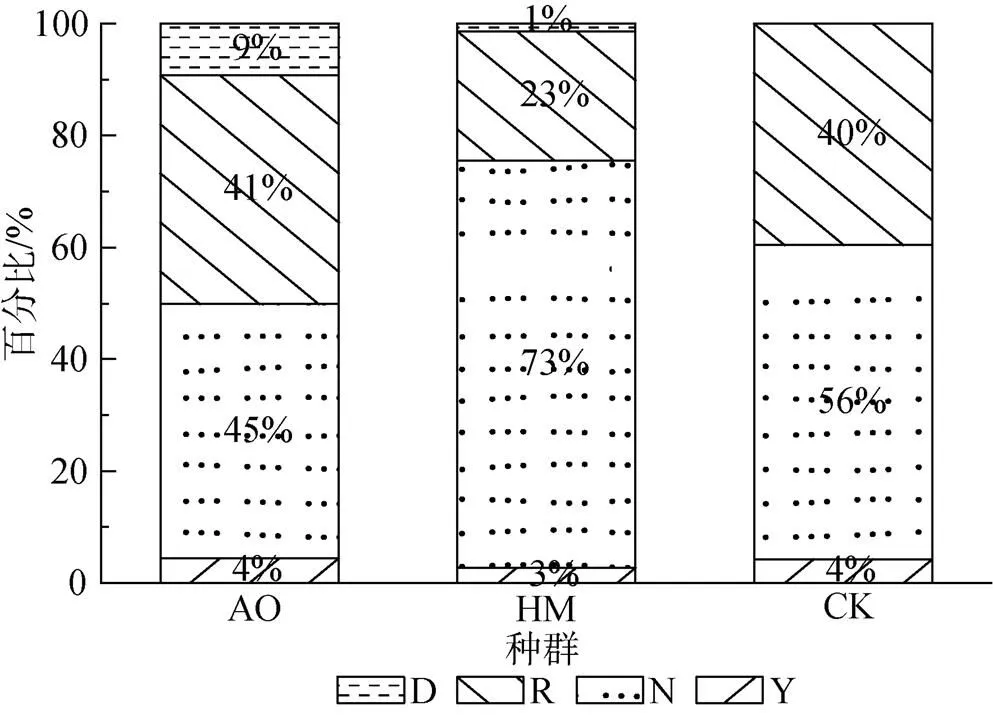
图1 毛乌素固定沙丘种群的结构组成
Figure 1 Structure composition of fixed dunes population in Mu Us sandy land
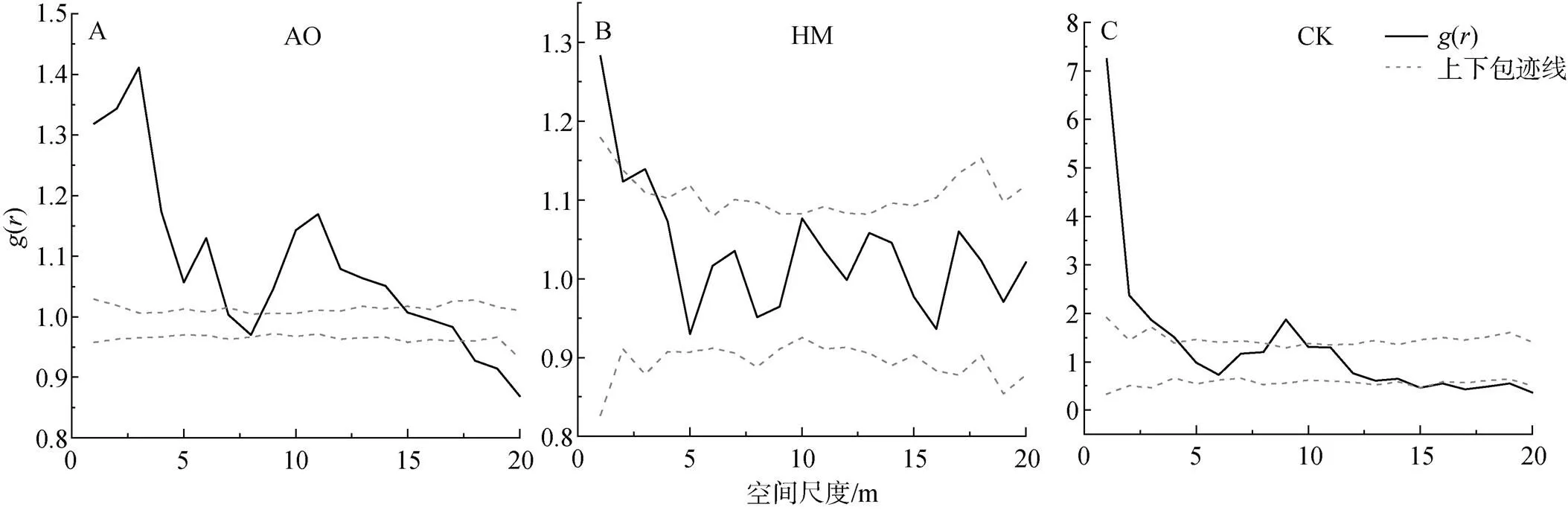
图2 基于完全随机模型(CSR)的种群的空间分布格局
Figure 2 Spatial distribution pattern of population based on completely random model

图3 种间的空间关联性
Figure 3 Interspecies spatial association
对不同生长发育阶段的柠条种群而言(图5D), 在小尺度下呈现正相关, 大尺度上呈现无关联或负关联。具体而言, 柠条种群中营养株和生殖株在小于4 m的尺度范围内呈现正相关, 在4—18 m的尺度范围内不存在相关性, 当尺度进一步增大, 大于18 m时, 两者则表现为负相关。
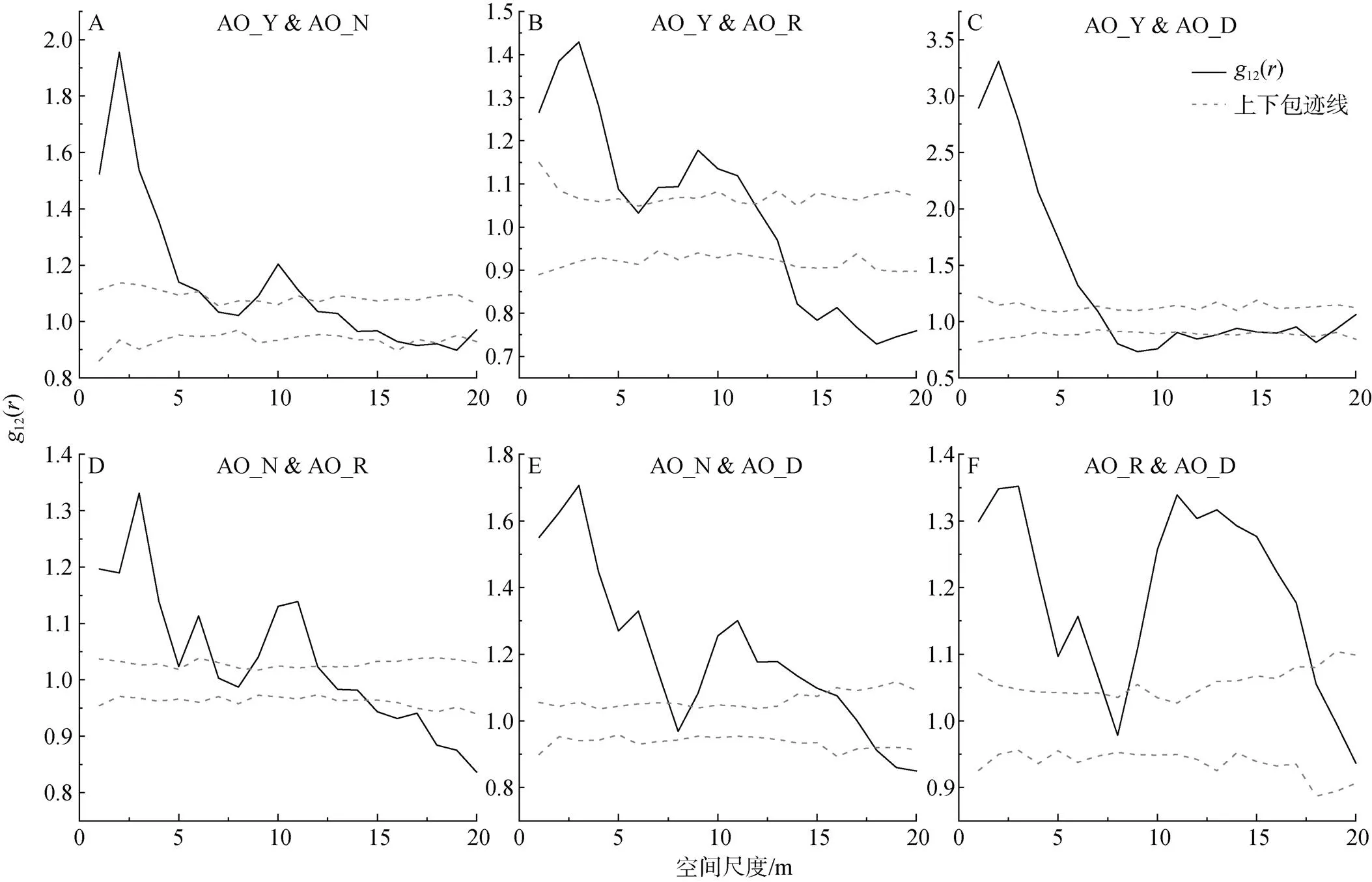
图4 油蒿不同生长发育阶段个体之间的关联性
Figure 4 Correlation between individuals at different growth stages of AO
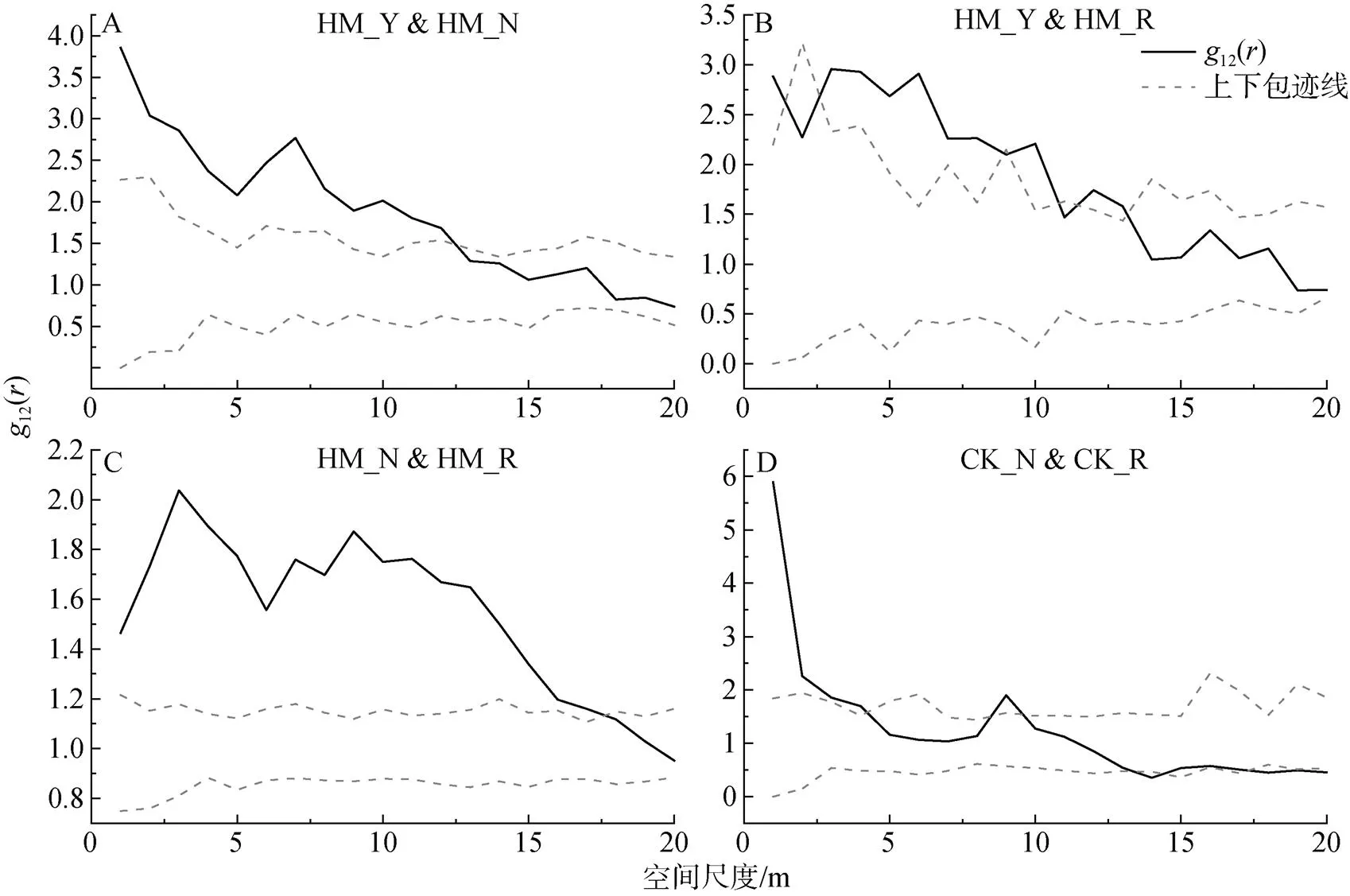
图5 杨柴和柠条不同生长发育阶段个体之间的关联性
Figure 5 Correlation between individuals at different growth stages of HM and CK
3 讨论
3.1 空间尺度与空间格局
种群的空间分布格局通常由种群的特性、生存环境、种内种间的相互关系所决定[24]。不同种群的聚集强度和空间分布情况的变化是种群为适应生态环境, 从而能够更好地生存的一种策略[25]。油蒿、杨柴和柠条种群的空间分布格局与空间尺度大小存在紧密的联系。三个种群在研究尺度范围内多表现为聚集分布, 随着尺度增大, 聚集强度逐渐降低, 最后趋近于随机分布或均匀分布。这与许多种群的空间格局的研究结果一致, 种群通常在小尺度下呈现出聚集分布, 在大尺度下多为随机或均匀分布[6, 7, 16]。产生这种现象, 可能主要与种群的生物学特性和环境因素两个方面有关[15]。一方面, 油蒿的繁殖方式主要以种子繁殖为主, 但种子繁殖会一定程度下导致播种的成活率较低, 因此在种子扩散过程中, 由于传播距离的限制和风力风速的影响, 使油蒿个体在20 m尺度下存在密切的关联关系, 呈现出聚集的分布格局[7, 26–27]。另一方面, 在小尺度下同一个种群对于环境的偏好和资源(水分、土壤、地形等)的需求基本相似, 种群会利用聚集的分布方式产生群体效应, 从而能够有效抵抗其他物种的竞争和入侵, 抵御恶劣生态环境的影响, 提高种群存活率, 维持种群稳定性[13, 28]。
随着种群不断发展, 种群对生存资源的需求不断增加, 特别是干旱生态系统中, 种群受到水分的胁迫, 其对资源利用的方式发生变化, 从而导致种内和种间竞争加剧, 处于竞争劣势的个体死亡, 种群密度降低, 个体间距逐渐增加, 最终在大尺度下种群逐渐趋向于随机或均匀分布[13]。本研究中, 油蒿种内不同发育阶段个体之间在尺度增大时普遍出现负相关的排斥作用, 导致幼年株发育受阻, 一定程度证实了种内竞争加剧的现象存在。
杨柴和柠条在研究尺度范围内主要呈现出的是随机分布, 这可能是由于杨柴和柠条的根系较深, 主要利用深层土壤水分, 受环境因素和资源竞争的影响较小, 从而主要表现为随机分布或均匀分布[29]。同时, 杨柴属于克隆生长的植物, 其具有合轴型分枝类型和游击型基株构型, 在小尺度上也会呈现聚集分布的特性[30]。
3.2 空间尺度与种间关联
种群间的相关关系, 通常是在生物学特性、种内种间的相互作用(竞争、依赖等)、生存环境等多种因素的共同作用下, 种群为适应生态环境并合理利用资源占据生态位所产生的结果[31]。在不同空间尺度下, 分析不同种群个体之间的关联性, 对于研究种群间相互作用机制和群落动态具有重要的意义。
油蒿和杨柴是在较小尺度下负相关性较强, 随着尺度增大, 两者主要表现为没有明显的关联关系。不同物种间的相关关系表现为负相关性, 可能有以下两个方面的原因: 一方面, 不同种群对环境的适应能力不同, 生态位重叠程度较小, 对于生境内资源的利用方面存在差异; 另一方面, 单位面积内资源有限, 不同种群间的对于生存资源(水分、土壤养分等)的竞争需求相同, 在数量上占绝对优势的种群对其他劣势种群的竞争排斥作用增强[31, 33]。同时油蒿耐干旱性较强, 在有性繁殖与无性繁殖方面均具有良好的生物学特性, 这使油蒿在相对较差的气候条件下, 仍然成为种间竞争的优胜者, 逐渐演替为较为稳定的种群[34]。在小尺度下, 这种较为激烈的竞争排斥作用导致油蒿与杨柴之间呈现出负相关关系, 但随着尺度增大, 油蒿与杨柴均逐渐呈现出随机或均匀分布, 种群的空间分离格局降低了种群的相遇几率, 同时生存所需的空间和资源竞争减弱, 因此油蒿与杨柴种群最终呈现出无关联关系的稳定状态。
油蒿之所以能在生态系统中占据一定优势地位,在研究区固定沙丘上与柠条等灌木形成较大数量差异, 主要归因于以下几个方面: 一是油蒿的种子产量、种子库和幼苗密度显著地高于柠条等灌木[27]; 二是固沙植被演替过程中所形成的生物土壤结皮具有较高的持水能力, 会阻止降水对深层土壤的补给, 导致根深系灌木数量减少直至退出生态系统, 巩固了油蒿的地位; 同时生物土壤结皮会阻止其他灌木种子补给到土壤种子库, 油蒿种子由于具有体积较小和见光萌发的特点, 更有利于萌发和幼苗成长[27, 35–36]; 三是油蒿比杨柴和柠条等灌木的根系分布更浅[37], 对于水分的竞争能力更强, 可以更快、更充分地利用降水[38]。
3.3 种内不同生长发育阶段之间的相关关系
种内关联性是指同一种群不同生长发育阶段个体之间在空间分布位置上的相互关联性, 其在一定程度上反映了种群的生长发育现状和种群内的相互关系[39]。种内空间正相关和负相关分别反映了种群内互利互惠和竞争排斥的生态关系, 空间无关联反映了没有明显的生态关系[6]。同一个种群在一定区域内生长发育具有相同的适应性, 其不同生长发育阶段之间的关联关系, 会受到种群数量和体积大小、生存环境、种群之间的依赖和竞争关系、历史因素以及其他因素的影响[24]。
本研究发现油蒿与杨柴不同生长发育阶段的种内关联关系在较广泛的尺度范围内表现为正相关, 随着尺度增大关联性逐渐降低, 表现为无相关, 甚至油蒿的种内关联关系出现负相关趋势。这可能与种群在小尺度下呈现出聚集分布, 大尺度下呈现出随机或均匀分布有密切联系[40]。虽然目前有研究表明, 种群的竞争压力主要是因为种内竞争产生的[41], 但油蒿与杨柴的种内关联关系在较大尺度范围内没有明显的排斥竞争的现象, 这可能是由于同一种群具有相同的生物学特性造成的。一方面, 同种植被的生境偏好和环境适应性相同, 对于资源的利用具有相似性, 产生生态位重叠, 有利于种群内部互惠互利、共存发展[40]。另一方面, 径级较小的植株对于环境资源(水分、土壤、养分、地形等)的竞争力较弱[42], 同时种群呈现聚集分布, 形成群体效应[43], 从而径级大、竞争实力强的植株在一定范围内对生境的改变使得径级较小的植株更加适合生存, 彼此交错分布, 充分共享和利用资源[13, 44]。
该研究还发现灌木(油蒿、杨柴)的生长发育阶段越接近, 个体间的正相关性越强; 反之, 植株的生长发育阶段差距较大, 个体间的正相关性就越弱, 甚至在大尺度下表现为无相关或负相关。由此可见, 形体大小相近的个体对于种群的协调发展更有利, 而形体差距较大的个体之间可能由于对资源的需求和竞争不能达到平衡稳定状态, 从而产生种内竞争,呈现出负关联关系[13]。除此之外, 柠条营养株和生殖株个体之间主要表现为无关联关系, 这可能由于柠条主要呈现为随机或均匀分布, 不同龄级之间保持一定距离, 彼此之间互不干扰、独立生长, 在长时间的群落演替过程中占据适宜的生态位, 减缓了种群内资源争夺的压力[13, 44]。
4 结论
研究基于固沙灌木个体的位置信息和生长发育阶段信息, 分析了毛乌素沙地固定沙丘上油蒿、杨柴和柠条种群的空间分布格局及种间、种内关联性。空间分布格局研究表明: 三个种群在小尺度下呈现聚集分布, 大尺度下为随机或均匀分布; 在研究尺度范围内, 杨柴和柠条主要呈现随机分布。这主要是由种子扩散机制、生境偏好、群体效应的生存策略以及密度制约造成的。
油蒿与杨柴两个种群种间关联研究表明: 油蒿和杨柴在较小尺度下负相关, 其他尺度下主要表现为无相关关系。种间负相关可能是由于生态位重叠程度小, 对于有限的资源种间竞争激烈引起的。
三个种群的种内关联研究表明: 柠条的种内关联关系基本呈无关联, 油蒿和杨柴的种内关系基本呈正相关, 随着尺度进一步增加, 会呈现负相关或无关联。除此之外, 同一种群个体的生长发育阶段越接近, 个体间的正相关性越强。
研究结果进一步加深了我们对于毛乌素沙地优势种群空间格局的认识, 有利于更好地理解毛乌素沙地固沙灌木种群的生物学特性以及形成机制, 为毛乌素沙地防风固沙建设提供了一定参考依据。本研究采用的点格局方法主要利用个体的位置和生长发育阶段信息进行分析, 对于个体内部以及个体间差异的关注度不够。因此, 未来可对样地内固沙灌木进行动态监测, 结合个体自身的生物学特性以及生境因素, 进一步阐明种群空间格局形成机制。
[1] 杨君珑, 王辉, 王彬, 等. 子午岭油松林灌木层主要树种的空间分布格局和种间关联性研究[J]. 西北植物学报, 2007, 27(4): 4791–4796.
[2] 郭忠玲, 马元丹, 郑金萍, 等. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究[J]. 应用生态学报, 2004, 15(11): 2013–2018.
[3] FANGLIANG H, LEGENDRE P, LAFRANKIE J V. Distribution patterns of tree species in a Malaysian tropical rain forest[J]. Journal of Vegetation Science, 1997, 8(1): 105–114.
[4] DRUCKENBROD D L, SHUGART H H, DAVIES I. Spatial pattern and process in forest stands within the Virginia piedmont[J]. Journal of Vegetation Science, 2005, 16(1): 37–48.
[5] CONDIT R, ASHTON P S, BAKER P, et al. Spatial patterns in the distribution of tropical tree species[J]. Science, 2000, 288(5470): 1414–1418.
[6] 张金屯. 植物种群空间分布的点格局分析[J]. 植物生态学报, 1998, 22(4): 57–62.
[7] 杨洪晓, 张金屯, 吴波, 等. 毛乌素沙地油蒿种群点格局分析[J]. 植物生态学报, 2006, 30(4): 563–570.
[8] BRISSON J, REYNOLDS J F. The effect of neighbors on root distribution in a creosote bush() population[J]. Ecology, 1994, 75(6): 1693–1702.
[9] 郑元润. 不同方法在沙地云杉种群分布格局分析中的适用性研究[J]. 植物生态学报, 1997, 21(5): 89–93.
[10] 戴小华, 余世孝. GIS支持下的种群分布格局分析[J]. 中山大学学报(自然科学版), 2003, 42(1): 75–78.
[11] 尤海舟, 贾成, 樊华, 等. 格局分析的最新方法——点格局分析[J]. 四川林业科技, 2009, 30(6): 106–110.
[12] 王本洋, 余世孝. 种群分布格局的多尺度分析[J]. 植物生态学报, 2005, 29(2): 235–241.
[13] 拓锋, 刘贤德, 刘润红, 等. 祁连山大野口流域青海云杉种群空间格局及其关联性[J]. 植物生态学报, 2020, 44(11): 1172–1183.
[14] HILL M O. The intensity of spatial pattern in plant communities[J]. The Journal of Ecology, 1973, 61(1): 225–235.
[15] 沈渭寿. 毛乌素沙地3种飞播植物种群分布格局的研究[J]. 中国沙漠, 1998, 18(4): 78–84.
[16] 刘法, 杨海龙, 高甲荣, 等. 毛乌素沙地羊柴种群点格局分析[J]. 中国水土保持科学, 2011, 9(4): 98–103.
[17] 崔强, 高甲荣, 李柏, 等. 宁夏毛乌素沙地3种常用固沙植物种群点格局分析[J]. 生态与农村环境学报, 2011, 27(3): 58–63.
[18] RIPLEY B D. Modelling spatial patterns[J]. Journal of the Royal Statistical Society: Series B (Methodological), 1977, 39(2): 172–192.
[19] 张新时. 毛乌素沙地的生态背景及其草地建设的原则与优化模式[J]. 植物生态学报, 1994, 18(1): 1–16.
[20] 郭柯, 董学军, 刘志茂. 毛乌素沙地沙丘土壤含水量特点——兼论老固定沙地上油蒿衰退原因[J]. 植物生态学报, 2000, 24(3): 275–279.
[21] 吴波, 慈龙骏. 毛乌素沙地景观格局变化研究[J]. 生态学报, 2001, 21(2): 191–196.
[22] 魏宁宁, 母艳梅, 姜晓燕, 等. 毛乌素沙地油蒿-杨柴灌丛生态系统蒸散组分分配及其影响因子[J]. 应用生态学报, 2021, 32(7): 2407–2414.
[23] LEWIS A, BRIDLE S. Cosmological parameters from CMB and other data: A Monte-Carlo approach[J]. Physical Review D, 2002, 66(10): 103511.
[24] VEBLEN T T, ASHTON D H, SCHLEGEL F M. Tree regeneration strategies in a lowland Nothofagus-dominated forest in south-central Chile[J]. Journal of Biogeography, 1979, 6(4): 329–340.
[25] HUBBELL S P. Neutral theory and the evolution of ecological equivalence[J]. Ecology, 2006, 87(6): 1387–1398.
[26] 冯丽, 张景光, 张志山, 等. 沙坡头人工固沙区油蒿繁殖分配的研究[J]. 中国沙漠, 2008, 28(3): 473–477.
[27] 王刚, 梁学功. 沙坡头人工固沙区的种子库动态[J]. 植物学报, 1995, 37(3): 231–237.
[28] 韩路, 王海珍, 彭杰, 等. 塔里木河上游天然胡杨林种群空间分布格局与动态研究[J].西北植物学报, 2007, 27(8): 1668–1673.
[29] 肖春旺, 周广胜, 马风云,. 施水量变化对毛乌素沙地优势植物形态与生长的影响[J]. 植物生态学报, 2002, 26(1): 69–76.
[30] 陈玉福, 董鸣. 毛乌素沙地根茎灌木羊柴的基株特征和不同生境中的分株种群特征[J]. 植物生态学报, 2000, 24(1): 40–45.
[31] 彭舜磊, 闫小婷, 齐光, 等. 伏牛山栓皮栎群落优势种群种间关联动态分析[J]. 森林与环境学报, 2018, 38(3): 326–333.
[32] WARD J S, PARKER G R, FERRANDINO F J. Long-term spatial dynamics in an old-growth deciduous forest[J]. Forest Ecology and Management, 1996, 83(3): 189–202.
[33] 李智叁, 李凤日. 乌兰布和沙漠天然白刺种群及主要伴生种种间关联性研究[J]. 植物研究, 2008, 28(1): 98–103.
[34] 张德魁, 王继和, 马全林, 等. 油蒿研究综述[J]. 草业科学, 2007(8): 30–35.
[35] 黄正英, GUTTERMAN Y. 油蒿与中国和以色列沙漠中的两种蒿属植物种子萌发策略的比较[J]. 植物学报, 2000, 42(1): 71–80.
[36] 马骥, 李俊祯, 孔红. 我国沙区6种蒿属植物的种子微形态特征[J]. 中国沙漠, 2002, 22(6): 586–590.
[37] 张志山, 樊恒文, 赵金龙, 等. 沙漠人工植物群落的根系分布及动态[J]. 中国沙漠, 2006, 26(4): 637–643.
[38] 唐海萍, 史培军, 李自珍. 沙坡头地区不同配置格局油蒿和柠条水分生态位适宜度研究[J]. 植物生态学报, 2001, 25(1): 6–10.
[39] 王磊, 孙启武, 郝朝运, 等. 皖南山区南方红豆杉种群不同龄级立木的点格局分析[J]. 应用生态学报, 2010, 21(2): 272–278.
[40] 张金屯, 孟东平. 芦芽山华北落叶松林不同龄级立木的点格局分析[J]. 生态学报, 2004, 24(1): 35–40.
[41] STOLL P, PRATI D. Intraspecific aggregation alters competitive interactions in experimental plantcommunities[J]. Ecology, 2001, 82(2): 319–327.
[42] 张健, 郝占庆, 宋波, 等. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J]. 应用生态学报, 2007, 18(8): 1681–1687.
[43] 赵峰侠, 尹林克. 荒漠内陆河岸胡杨和多枝柽柳幼苗种群空间分布格局及种间关联性[J]. 生态学杂志, 2007, 26(7): 972–977.
[44] 涂洪润, 李娇凤, 刘润红, 等. 桂林岩溶石山檵木种群空间格局及其关联性[J]. 应用生态学报, 2019, 30(8): 2621–2630.
Spatial distribution pattern and spatial correlation of main sand-fixing shrubs in fixed dunes in Yanchi region, Mu Us sandy land
NIU Chengxu1, ZHANG Dinghai1,*, ZHANG Zhishan2, WANG Yanli3
1.Center for Quantitative Biology, College of Science, Gansu Agricultural University, Lanzhou 730070, China 2. Shapotou Desert Research and Experimental Station, Northwest Institute of Eco-Environment and Resources, Lanzhou 730000, China 3. College of Forestry, Gansu Agricultural University, Lanzhou 730070, China
The purpose of this research on the spatial distribution pattern and correlation of populations is to predict the succession trend of populations and vegetation reconstruction which has a great significance meaning for further research investigation. Based on the geographic location and growth stage data of sand-fixing shrubs on fixed sand dunes in the Mu Us Sandy Land, the spatial distribution pattern and correlation of three dominant populations of(AO),(HM) and(CK) were analyzed by using univariate and bivariate paired correlation functions and Monte Carlo fitting test. The results are as follows. (1) The three populations were aggregated at small scale and randomly or uniformly distributed at large scale; the populations of HM and CK showed aggregation distribution within the spatial scale of the study. (2) The interspecies relationships showed thatAO and HM were negatively correlated at the scale less than 5 meters and basically uncorrelated at other scales. (3) The intraspecific correlation among different growth and development stages of the populations showed thatthe intraspecific relationships of the 3 populations were basically positively correlated at small scales. At large scales, there was a negative correlation between the intraspecifc relationships of AO,while there was no correlation between the intraspecifc relationships of HM and CK.
Mu Us sandy land; sand-fixing shrubs; point pattern analysis; spatial distribution pattern; spatial correlation
10.14108/j.cnki.1008-8873.2024.01.001
Q948.15
A
1008-8873(2024)01-001-09
2021-07-30;
2021-09-30
国家自然科学基金项目(42361016); 甘肃省科技创新计划—“西部之光”人才计划项目(22JR9KA032);甘肃省科技计划项目(21JR7RA831)
牛程旭(1997—), 女, 河北沽源人, 硕士研究生, 主要从事应用统计学研究, E-mail: 286116811@qq.com
通信作者:张定海, 男, 副教授, 硕士导师, 主要从事生态水文模型及统计学在生态学、经济学中的应用, E-mail: zhangdh@gsau.edu.cn
牛程旭, 张定海, 张志山, 等. 毛乌素沙地盐池地区固定沙丘主要固沙灌木的空间分布格局与空间关联性研究[J]. 生态科学, 2024, 43(1): 1–9.
NIU Chengxu, ZHANG Dinghai, ZHANG Zhishan, et al. Spatial distribution pattern and spatial correlation of main sand-fixing shrubs in fixed dunes in Yanchi region, Mu Us sandy land[J]. Ecological Science, 2024, 43(1): 1–9.
猜你喜欢
小猕猴学习画刊(2022年3期)2022-03-28
现代农村科技(2021年5期)2021-12-07
现代盐化工(2021年4期)2021-09-19
金秋(2021年8期)2021-07-27
音乐天地(音乐创作版)(2020年12期)2021-01-11
中国煤炭工业(2020年1期)2020-12-28
应用化工(2020年4期)2020-06-04
中国水土保持科学(2019年6期)2019-04-26
当代陕西(2019年7期)2019-04-25
铁道标准设计(2018年11期)2018-10-20
