
近自然经营对马尾松林优势树种生态位及种间关系的影响*
2024-04-12 07:10邹丰虎周雯柴宗政
西部林业科学 2024年1期
邹丰虎,周雯,柴宗政
(1.贵州大学 林学院,贵州 贵阳 550025;2.贵阳市林业局,贵州 贵阳 550081)
生态位与种间联结是衡量物种间相互关系的重要方法[1],不仅能有效表征群落稳定性,而且还能够反映群落动态变化[2]。其中,生态位反映物种在时空上的位置,对解释森林群落中物种共存和竞争有较强的指示意义[3-5]。种间联结是指物种或种群在空间分布上的相互关联性[6],对特定物种的保护具有指导作用[7],能反映林木邻近个体之间的竞争作用,从而对种群发展及群落演替产生影响。近年来,生态位及种间联结被广泛用于研究森林群落种间关系及物种共存[1,8],为森林生态系统稳定性评价及发展趋势推演提供重要科学依据[9-10]。
马尾松(PinusmassonianaLamb.)是中国南方地区种植面积最广的乡土针叶树种,对我国木材供应和森林生态系统功能发挥起到至关重要的作用[11-13]。然而,由于长期大规模采伐,现存马尾松林主要为人工纯林,存在树种单一、中幼林比例高、林分密度大、抵抗病虫害能力弱、生产力低下等问题。这些问题使得马尾松林产品难以满足市场需求,尤其是优质大径材的需求[14]。通过实践探究,将近自然经营理论与技术应用于马尾松林的经营管理中[15-16],以提高其质量和效益。研究显示近自然经营对改善马尾松林竞争关系[16]、促进林下更新[17]、优化林分结构[18],对加快人工纯林向异龄复层混交林方向发展具有较好的推动作用。近自然经营主要以目标树单株择伐的方式改变森林内部生境和树种优势度,从而使树种适应能力、竞争能力和种间关系发生变化,从而影响森林生态系统结构和功能[19]。近年来,对于马尾松林优势树种的生态位和种间关系在近自然经营下的变化的相关研究报道较少,然而,这类研究对于进一步了解近自然经营对森林群落中树种间关系的调控以及混交树种筛选与配置具有较高的指导意义。
本研究以中德财政合作贵州森林可持续经营项目中的马尾松近自然经营试验示范区为研究区域,选取近自然经营已有10余年的马尾松林为研究对象。通过样地调查,并采用生态位测定、方差比率法(VR)、χ2检验和Spearman秩相关检验等方法,探究了近自然经营对马尾松林中12个乔木优势树种在成树、幼树和幼苗这3个生长阶段的生态位及种间关系的影响。通过该项研究以期明确近自然经营技术措施是否能有效改善马尾松林的生态位和种间关系,并为后续的经营管理提供实践基础和科学依据。
1 材料与方法
1.1 研究区概况
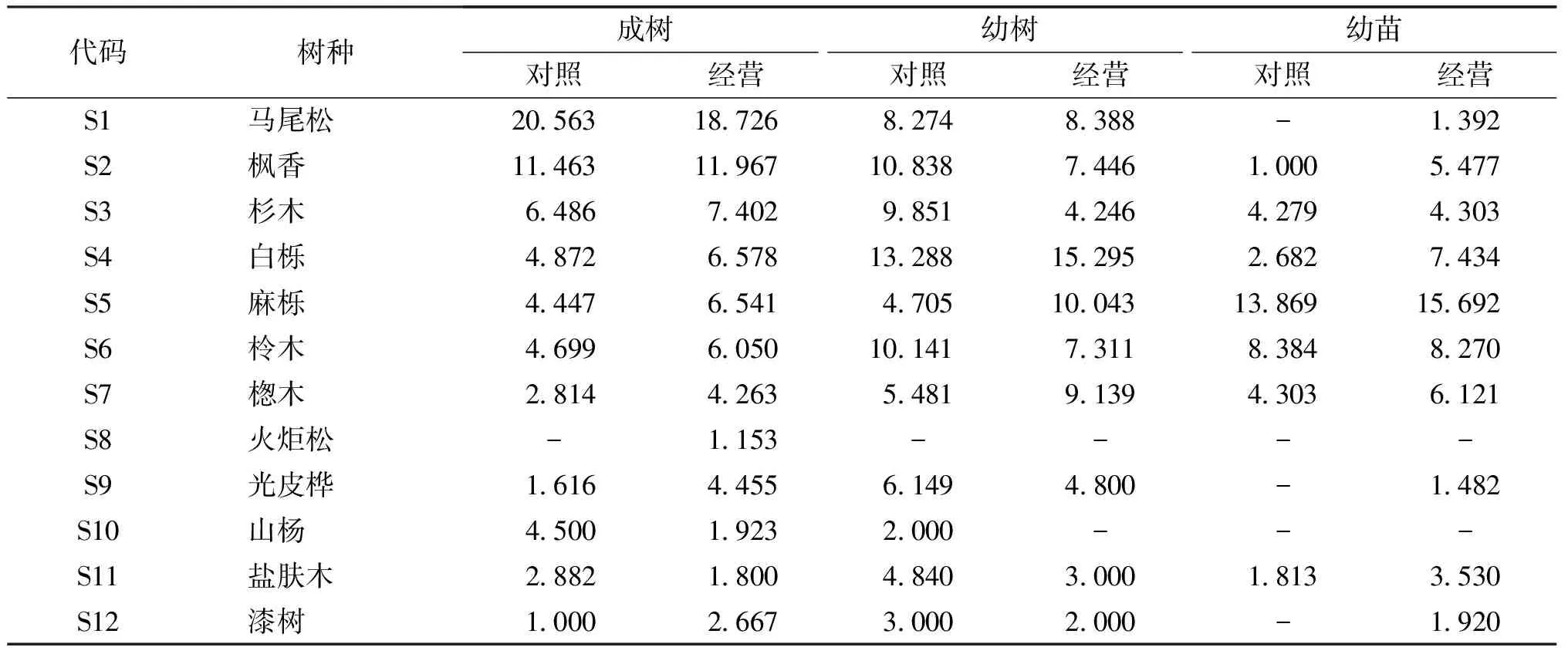
研究区位于贵阳市开阳县龙岗镇,属黔中高原区,主要地貌类型为低中山、中中山山地及丘陵,地势起伏较大,地下暗河伏流,地面山坡、河流、沟谷镶嵌分布,平均海拔约1 200 m。属亚热带湿润温和型气候,冬无严寒,夏无酷暑,平均气温15.3 ℃,无霜期年平均长达300 d以上;雨热同季,雨量充沛,年均降水量1 200 mm。土壤母质以砂页岩、石灰岩、白云岩为主,土壤类型有黄壤和石灰土,主要为酸性黄壤。森林类型以马尾松、杉木(Cunninghamialanceolata)等针叶人工林为主,另外还存在少量枫香(Liquidambarformosana)、白栎(Quercusfabri)、麻栎(Quercusacutissima)等阔叶树种。
本研究以实施近自然经营10余年的马尾松林为研究对象。近自然经营措施是基于目标树单株作业法,以林分平均胸径(d)划分马尾松林生长阶段,并据此制定相应的经营措施(表1)。

表1 马尾松林不同生长阶段的近自然经营措施
1.2 样地设置与调查
参考研究区马尾松林分布及近自然经营实施情况,按年龄梯度共计设置24组成对样地(1个经营样地和1个对照样地,两个样地具有相同的林分年龄和相近的立地条件),包含5组幼龄林、11组中龄林和8组近熟林,共计48个样地。布设圆形样地进行调查,样地半径为14.57 m,面积为667 m2。每个圆形样地内均匀布设9个同心样圆,圆心间距离为5 m,圆心用PVC管固定,其中大的样圆(4 m2)调查灌木和更新,小的样圆(2 m2)调查草本植物,另外布设4个面积为1 m2的样圆调查土壤。对样地内所有胸径≥5 cm的立木使用全站仪(TOPCON-GTS-602AF)每木定位,实测胸径、树高、冠高、枝下高、冠幅,记录林木质量、病虫害状况、生活力并进行林木分级。在4 m2样圆内对林下更新(幼树:1 cm≤胸径(DBH)<5 cm;幼苗:胸径<1 cm)及灌木的种类、株高、株数、盖度和病虫害状况等进行调查。在2 m2样圆内调查草本植物的种类、株高、株数、盖度和病虫害状况。
1.3 数据分析
1.3.1 重要值计算
重要值(IV)反映物种的相对优势程度。计算方法如下:
IV=(Ar+Pr+Fr)/3
式中:Ar为相对多度,Pr为相对显著度,Fr为相对频度。
1.3.2 生态位宽度
采用Levins提出、并由Colwell和Futuyma修改的公式来计算[20]。
式中:Bf为第f物种的生态位宽度;r为资源水平数;Pik为第i物种在第k个资源水平下的重要值占该物种在所有资源水平上重要值总和的比例。
1.3.3 生态位重叠度
根据Pianka公式计算生态位重叠[21]:
式中:Oij表示物种i和j的生态位重叠值;r为资源水平数;Pik和Pjk分别为第i和j物种在第k个资源水平下的重要值占该物种在所有资源水平上重要值总和的比例。
1.3.4 总体联结性
采用Schluter提出的方差比率(varianceratio,VR)法来测定多物种间的总体联结性,并用统计量W来检验多物种间的关联显著度,计算公式如下。
W=VR×N
式中:Pi=nI/N,ni为物种i出现的样方量,N为总样方量;S为总物种量,Tj为样方j内出现物种的总量;t为样方中物种的平均值。
1.3.5 卡方检验(χ2)
种间联结采用χ2检验进行定性研究,根据2×2列联表的χ2统计量测定成对种间的联结性,将12个优势树种是否在48个样地中出现转化为48×12的0、1二元矩阵,0表示树种未在样地中出现,1表示出现。由于χ2分布属于连续型分布,而本研究的数据为非连续性数据,因此采用Yates的连续校正公式计算[22]。
式中:N为总样地量,a为2个物种均出现的样地量,b、c为仅有1个物种出现的样地量,d为2个物种均未出现的样地量。若χ2<3.841(即P>0.05),认为2个物种独立分布,即种间联结性不显著;若3.841<χ2<6.635(即0.01 1.3.6 Spearman秩相关系数 χ2检验仅定性判断了种间联结性是否显著,并不能确定不显著的种对间是否具有联结性,也无法定量区分联结程度的大小,不能清晰地表示物种间联结性的差异性[23]。Spearman秩相关检验是基于数量数据,能更准确地反映物种间的相互关联性,从而对χ2检验进行有效补充和完善[24]。因此,在种间联结χ2检验后,进一步采用Spearman秩相关检验确定其联结程度。本研究利用12个优势乔木树种的重要值作为Spearman秩相关检验的数量指标,对种对间的线性关系进行定量分析,分析它们同时存在的可能性。 Spearman秩相关系数计算公式如下。 数据处理与制图使用R4.2.2完成,重要值计算采用forestHES包,生态位特征及种间关系分析使用spaa包,并使用ggplot2进行制图。 48个马尾松样地中共出现乔木树种36种,隶属于28科33属,乔木树种组成相对简单,见表2。 由表2可知,在成树、幼树和幼苗三个生长阶段,马尾松、枫香、杉木、白栎、麻栎、柃木(Euryajaponica)、楤木(Araliachinensis)、火炬松(Pinustaeda)、光皮桦(Betulaluminifera)、山杨(Populusdavidiana)、盐肤木(Rhuschinensis)和漆树(Toxicodendronvernicifluum)这12个树种的重要值均相对较高(表2),其中成树阶段重要值占比超过了98%,幼树阶段重要值占比超过了91%,幼苗阶段重要值占比超过了83%。这表明这12个树种在马尾松林中占据着极其重要的地位和作用,本研究中将他们作为研究区马尾松林优势树种。 马尾松的重要值在成树和幼树阶段占比较高且经营林分的重要值普遍低于对照林分;幼苗阶段的马尾松不占优势,经营林分中马尾松重要值占比仅0.309%,而对照林分未调查到马尾松幼苗。这表明近自然经营有效减弱了马尾松在成树和幼树阶段的优势度,同时一定程度上提升了马尾松幼苗的存活率。此外,枫香、杉木、白栎、柃木和楤木等树种在经营林分中的重要值高于对照林分,而麻栎、光皮桦和山杨等树种在经营林分中的重要值低于对照林分,表明近自然经营改变了树种组成及数量配置。 在成树、幼树和幼苗三个生长阶段,马尾松、枫香、杉木、白栎、麻栎、柃木、楤木共7个优势树种具有相对较高的生态位宽度,表明以上树种在研究区马尾松林中分布相对广泛(表3)。此外,大部分树种的生态位宽度在经营林分明显高于对照林分。成树阶段,除马尾松、山杨和盐肤木的生态位宽度在经营林分低于对照林分,其他树种的生态位宽度均高于对照林分;幼树阶段,除了枫香、杉木、柃木、光皮桦、山杨、盐肤木和漆树的生态位宽度在经营林分低于对照林分,其他树种的生态位宽度均高于对照林分;幼苗阶段,除了柃木的生态位宽度在经营林分低于对照林分,其他树种的生态位宽度均高于对照林分。可见,近自然经营能够有效提升树种的生态位宽度,增强树种的适生能力。 表3 马尾松林不同生长阶段优势树种生态位宽度 成树阶段的对照林分中,12个优势树种共形成55个种对,其中49个种对存在生态位重叠,生态位重叠度平均值为0.474(图1)。其中9对处于0.80~1.00之间,占比16.36%;14对处在0.40~0.80之间,占比25.45%;26对处于0.00~0.40之间,占比47.27%。成树阶段的经营林分中,12个优势树种共形成66个种对,其中53个种对存在生态位重叠,生态位重叠度平均值为0.449。其中4对处于0.80~1.00之间,占比6.06%;23对处在0.40~0.80之间,占比34.8%;26对处于0.00~0.40之间,占比39.4%。其中生态位重叠较大的种对分别是马尾松—山杨(1.00),枫香—光皮桦(0.90),麻栎—盐肤木(1.00),麻栎—漆树(1.00)。对比前者,经营过的林分种对间生态位重叠度减小了0.025。 图1 马尾松林不同生长阶段优势树种的生态位重叠度 幼树阶段的对照林分中,共形成55个种对,其中51对存在生态位重叠,生态位重叠度平均值为0.397。其中3对处在0.80~1.00之间,分别是马尾松—麻栎(1.00),枫香—杉木(0.86),白栎—柃木(0.82);14对处在0.40~0.80之间,占比25.5%;34对处在0.00~0.40之间,占比61.8%。幼树阶段的经营林分中,其中4对相互独立存在,5对处于0.80~1.00之间,分别为杉木—漆树(1.00),白栎—楤木(0.90),白栎—漆树(0.99),麻栎—漆树(1.00),楤木—盐肤木(0.92);12对处于0.40~0.80之间,占比21.8%;34对处于0.00~0.40之间。经营林分总种对数与对照林分一致,但平均值为0.356,相比前者减小了0.041。 幼苗阶段的对照林分中仅有7个树种共形成21个种对,其中15对存在生态位重叠,生态位重叠值在0.00~0.88之间,平均值为0.266。其中仅1对生态位重叠值达到0.88,为楤木—柃木;3对处于0.40~0.80之间;11对处在0.00~0.40之间,占比52.38%。幼苗阶段的经营林分中共有10个树种形成了45个种对,其中38个种对存在生态位重叠,平均值为0.238 5。其中2对处于0.80~1.00之间,分别为白栎—盐肤木(0.83),麻栎—盐肤木(1.00);7对处于0.40~0.80之间,占比15.6%;29对处于0.00~0.40之间,占比64.4%。经营与对照林分相比,总种对数增加了114%,但平均生态位重叠度减小了0.027 5。 可见,不同生长阶段对照林分中树种间生态位重叠程度均较高,特别是成树阶段,其中马尾松与其他乔木树种尤为突出,资源竞争比较激烈,近自然经营明显降低了各优势树种种对间的生态位重叠度,提高了生态位分化程度。 表4 马尾松林不同生长阶段优势树种的总体关联性 基于Spearman秩相关检验显示(图2、表5),成树阶段对照林分中,正相关的种对有33对,占比60.00%,其中极显著与不显著的种对数分别为2、31,分别占比3.60%、56.40%;负相关种对有22对,相关性都不显著,正负关联比为1.50。经营林分中,正相关的种对有32对,占比48.48%,其中极显著、显著和不显著的种对数分别为1、3和28,分别占比1.52%、4.55%和42.42%;负相关种对有34对,占比51.51%,其中显著和不显著的种对分别有2、32,分别占比3.03%、48.48%,正负关联比为0.94。以上结果表明,近自然经营后成树的种间关系被干扰,种间联结性减弱,相互独立性增强。 图2 马尾松林不同生长阶段优势树种种间的Spearman秩相关系数 表5 各优势树种不同生长阶段相关性的Spearman检验结果 幼树阶段对照林分中,正相关的种对有34对,占比61.82%,其中极显著、显著和不显著的种对数分别为2、1和31,分别占比3.64%、1.82%和56.36%;负关联的有21对,占比38.18%,其中极显著、显著和不显著的种对数分别为1、1和19,分别占比1.82%、1.82%和34.55%,正负关联比为1.62。经营林分中,正相关的种对有26对,占比57.78%,其中显著、不显著的正对数分别为2、24,分别占比4.44%和53.33%;呈负相关的种对有19个,占比42.22%,其中显著和不显著的种对数分别为2和17,分别占比4.44%和37.78%,正负关联比为1.37。以上结果表明,幼树阶段对照和经营林分的种间联结性差异不大,表明近自然经营对幼树的种间关系影响不明显。 幼苗阶段对照林分中,正相关的种对有8对,占比38.10%,其中极显著、显著和不显著的种对数分别为1、2和5,分别占比4.76%、9.52%和23.81%;呈负相关的种对有13个,占比61.90%,相关性皆不显著,正负关联比为0.62。经营林分中,幼苗种间呈正相关的种对有24个,占比53.33%,其中极显著、显著和不显著的种对数分别为1、2和21,占比2.22%、4.44%和46.67%;呈负相关的种对有21个,占比61.90%,相关性皆不显著,正负关联比为1.43。以上结果表明,经营林分中的幼苗的种对数比对照林分增加了24对,表明近自然经营有效改善了幼苗的生存和生长,使幼苗种间联结性更加复杂,群落更加稳定。 重要值和生态位宽度均反映了树种在群落中的相对优势程度和作用地位[25-26]。研究区马尾松以人工林居多,导致马尾松成树和幼树阶段的重要值和生态位宽度均较高(表2)。而幼苗阶段,马尾松重要值和生态位宽度明显不占优势,表明马尾松存在天然更新困难的问题,这主要是由于马尾松为喜光树种,幼苗阶段的生长需要良好的光照环境保证其生长,而研究区马尾松普遍存在密度过大且郁闭度高的问题,导致林下光照缺乏,马尾松幼苗难以生存[27]。这也是对照林分中未发现马尾松幼苗的原因。而经营林分,通过抚育采伐对主林层进行密度调控,伐后林内产生的林隙或林窗有效改善了林下光照环境,创造了适宜马尾松幼苗生长的光照环境,这也是经营林分中马尾松幼苗数量高于对照林分的主要原因。同时,林下光照环境的改善也为其他喜光树种的幼苗创造了较好的生境条件,因而经营林分中大部分幼树幼苗的生态位宽度也普遍高于对照林分。近自然经营通过抚育采伐有效降低了成树和幼树阶段马尾松的生态位宽度,提升了马尾松幼苗的生态位宽度和大部分优势树种的生态位宽度。由此可见,适度的抚育采伐有利于保持优势种群在群落中较大生态位宽度和较强的生态适应性。贾呈鑫卓等[19]对思茅松(P.kesiyavar.langbianensis)天然林生态位研究和巫志龙等[28]对天然次生林生态位研究也有力支持了本研究的结论。 大部分优势树种间生态位重叠程度不是很高,但马尾松与其他树种均存在生态位高度重叠,生态位重叠度处于0.54~1.00之间,且平均值大于0.7,主要是由于研究区马尾松林多为人工纯林,马尾松占比较高,树种单一。对照林分中成树、幼树和幼苗3个阶段的生态位重叠度平均值为0.474、0.397和0.266,经营林分中则分别为0.449、0.356和0.238 5,表明实施近自然经营后,各优势树种间生态位重叠度有所减小,降低了种间竞争,提高了生态位分化程度。这与蒋子涵等人的结论一致[29-31]。减少的生态位重叠度值主要分布于马尾松与其他树种形成的种对间,这主要是由于研究区的近自然经营以采伐马尾松林木为主,一定程度上减小了马尾松在林分中的优势度,从而减小了马尾松与其他树种生态位重叠程度。 群落总体关联性大多呈现正关联,表明群落树种正向多物种、多龄级和多层次发展,但极显著正联结种对较少,人工林群落树种间竞争不明显,这与姜俊等[8]对马尾松人工林种间联结性研究一致。Spearman秩相关检验结果也表明不同生长阶段经营和对照林分种对间存在一定差异但并不显著,且正负关联种对占比差异也不明显。这意味着该群落正处于演替初期,整体结构并不稳定,群落内物种还处在不断变化中,表明近自然经营对马尾松林种间关系的改善具有一定效果但并不理想,种间关系的改善还需要更长时间的演替及更具有针对性的措施。 本研究结果表明,马尾松林近自然经营过程中,仅通过以抚育采伐为主的密度调控实现种间关系的优化其作用是有限的,多项研究及生产实践均已证实,通过林下补植或套种阔叶树种加速针叶纯林向针阔混交林转化实现树种结构调整是有效途径,而适宜混交树种的筛选是优化种间关系和调整树种结构的关键。根据近自然经营的基本思想,在人工造林或营林过程中,根据原生植被分布规律,选择现有立地条件下适宜生长的乡土树种,科学筛选混交树种,培育异龄、复层、混交的林分结构是保证森林稳定健康发展的关键[32]。同时,种间联结可作为森林经营中树种配置的量化依据[8],对探究林分树种搭配、混交模式设计等方面具有重要的指导意义。因此,结合我们的研究,可以确定枫香和麻栎、白栎等栎类树种是研究区马尾松林较为适宜的混交树种,其原因有以下3个方面:(1)是研究区广泛分布的乡土树种,适应能力较强;(2)在马尾松林中具有较大的生态位宽度且幼苗、幼树和成树阶段均有分布,能与马尾松和谐相处建立良好的种间关系;(3)3个树种都为高大乔木且长寿命树种,可以与马尾松长期共存。基于此,本研究建议针对研究区马尾松纯林化程度过高且密度过大的这类林分,应在密度调控的基础上,结合林下补植枫香和麻栎、白栎等栎类树种,加速群落演替,尽快达到改善种间关系和提升群落稳定性的目的。
2 结果与分析
2.1 不同生长阶段优势树种的重要值
2.2 不同生长阶段优势树种的生态位宽度
2.3 不同生长阶段优势树种的生态位重叠度

2.4 不同生长阶段优势树种的总体联结性


2.5 不同生长阶段优势树种的种间联结性Spearman秩相关性分析


3 讨论与结论
猜你喜欢
植物研究(2023年5期)2023-09-09
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
防护林科技(2020年6期)2020-08-12
绿色科技(2019年5期)2019-11-29
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
杂草学报(2015年2期)2016-01-04
