
山羊口疮病毒四川株007基因克隆及其编码蛋白分析
2024-03-25 02:37张风雨姜雯玉
四川畜牧兽医 2024年3期
张风雨,杨 璐,姜雯玉,黄 忍
(成都威斯津生物医药科技有限公司,四川 成都 610218)
羊口疮病毒(ORFV)是痘病毒科、脊椎动物痘病毒亚科、副痘病毒属的典型代表[1]。ORFV具有高度的嗜上皮性,主要通过破损的皮肤及黏膜感染,是一种引起绵羊、山羊及人发病的急性高度接触性人兽共患病[2]。临床上主要以感染动物的唇、鼻孔、口腔黏膜、乳房、阴部等部位皮肤形成红斑、水疱、脓疱、丘疹和疣状痂皮为特征[3]。已有研究表明,羊口疮病毒为线性双链DNA 病毒,基因组大小为134~139 kb,其中包含末端基因区(ORFs001-008和112-134)和中间核心区(ORFs009-111),编码134个基因[4]。早有研究报道ORFV 编码dUTPase[5],dUTPase 由ORFV中483 bp的基因序列编码产生,该蛋白常以融合形式表达[6]。dUTPase 是一种普遍存在的酶,它催化dUTP 水解生成脱氧尿苷单磷酸(dUMP)和焦磷酸(PPI),从而降低细胞中dUTP/dTTP 的比值,阻止尿嘧啶进入DNA,降低发生错配的几率[7]。dUMP经甲基化变为胸腺嘧啶脱氧核糖核苷三磷酸(dTTP),dTTP 是胸腺核苷酸生物合成的前体。研究表明,当胞内dUTPase 酶活性不足以支持病毒进行有效复制时,编码dUTPase 基因的病毒则可以上调胞内dUTPase 酶活性,确保病毒正常复制[8]。本研究对山羊口疮病毒四川分离株007基因(dUTPase)进行PCR扩增、克隆,并利用生物信息学软件对该基因编码的蛋白结构进行预测,为进一步研究ORFV dUTPase蛋白在ORFV致病中的作用机制奠定基础。
1 材料与方法
1.1 材料 山羊口疮病毒四川分离株SC-LZ1和pET-32a(+)载体由西南民族大学动物医学实验室保存;DNA提取试剂盒、PCR试剂、胶回收试剂盒、Plasmid DH5α感受态细胞购至Takara 公司;限制性内切酶BamHI和XhoI购自NEB公司。
1.2 方法
1.2.1 ORFV 007基因的扩增 根据GenBank 中ORF strain SJ1(KP010356.1)dUTPase 基因序列,用Primer 5.0 软件设计一对带有酶切位点的特异性引物。上游引物:5′-CGCGGATCCATGGAG TTCTGCTGCACG-3′,下游引物:5′-CCGCTCGAG TTAAAGAGCTACATTTTACAATTTTGAT-3′。预期目的基因片段大小为507 bp,引物由生工生物工程(上海)股份有限公司合成。根据DNA 提取试剂盒说明提取山羊口疮病毒四川分离株SCLZ1 的DNA。按以下体系进行基因扩增:DNA模板2 μL,上下游引物各1 μL,PreMix 酶12.5 μL,ddH2O 8.5 μL,反应体系总体积为25 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性30 s,64 ℃退火30 s,72 ℃延伸45 s,共35个循环;72 ℃延伸10 min;16 ℃保存。
1.2.2 pET-32a(+)-007重组质粒的构建 将所扩增的目的基因经1.5%琼脂糖凝胶电泳后,采用胶回收试剂盒对目的条带进行胶回收。分别用限制性内切酶BamHI 和XhoI 对胶回收产物和pET-32a(+)进行双酶切(双酶切体系见表1)。在37 ℃水浴条件下酶切30 min,酶切产物采用纯化试剂盒进行纯化回收。将酶切纯化产物与pET-32a(+)在16 ℃金属浴中连接过夜(连接体系见表2)。连接产物转化入DH5α感受态细胞,再均匀涂布于含100 μg/mL AMP 的LB 平板上,再倒置于37 ℃恒温培养箱中过夜。次日,挑取疑似阳性单菌落至含有100 μg/mL AMP的LB液体培养基内,37 ℃160 r/min 增菌培养过夜。采用Plasmid Miniprep Kit质粒提取试剂盒提取质粒,对质粒进行PCR 鉴定和双酶切鉴定。将鉴定结果为阳性的质粒送往生工生物工程(上海)股份有限公司测序。将测序结果为阳性的质粒命名为pET-32a(+)-007。

表1 双酶切体系

表2 连接体系
1.2.3 ORFV 007基因序列分析 将测序所得序列在NCBI上进行同源性比较,并利用Mega7.0构建系统进化树。
1.2.4 dUTPase蛋白结构及功能预测 采用在线软件ExPasy 服务器上ProtParam 工具对基因编码的氨基酸序列进行理化性质分析;利用ExPasy服务器的ProtScale工具,计算基于K-D法的蛋白质疏水性;使用TMHMM Server v.2.0进行跨膜区分析;使用Signal P 3.0 server 进行信号肽预测;利用NPSA和CBS在线软件对该蛋白进行二级结构预测;利用NCBI 在线软件对蛋白保守域进行分析进而预测其功能;利用SWISS-MODEL 进行同源建模,预测其三级结构。
2 结果
2.1 ORFV 007 基因的扩增 以提取的山羊口疮病毒四川分离株SC-LZ1的DNA为PCR扩增模板,扩增产物经1.5%琼脂糖凝胶电泳,结果成功扩增出约500 bp条带,与预期扩增条带大小相符(图1)。
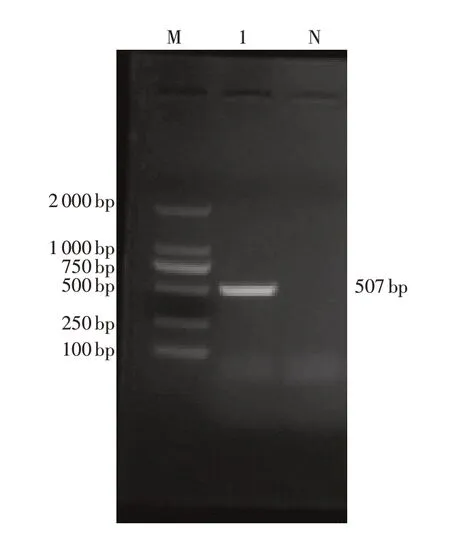
图1 ORFV 007 PCR扩增结果
2.2 重组质粒的双酶切鉴定 由图2可知,重组质粒双酶切产物与预期结果相符。

图2 pET-32a(+)-007重组质粒的双酶切鉴定
2.3 ORFV 007基因序列分析
2.3.1 ORFV 007 基因全序列 ORFV 007 基因全长为483 bp,测序结果显示扩增基因无缺失。
2.3.2 ORFV 007基因的遗传进化树 将本研究扩增测序的ORFV 007 基因与NCBI 上收录的所有ORFV 007 基因进行比对分析,并构建遗传进化树。结果该基因全序列与NCBI 收录的编号为MF4890951 基因的同源性最高,达98.96%;构建的系统进化树结果显示本试验分离毒株 与 MF489089.1、MF489095.1、MF489093.1、MF489088.1、MF489090.1 及MF489094.1 在同一分支,说明亲缘关系较近,但又单独为一小支,存在一定的变异(图3)。

图3 ORFV 007基因序列系统进化树
2.4 ORFV 007 编码的dUTPase 蛋白结构预测利用ExPasy 服务器上的ProtParam 工具对该蛋白进行理化性质分析,结果显示:ORFV 007 编码161个氨基酸,相对分子量为16 945.11,理论PI值为5.27,消光系数为11 960,不稳定系数为21.53,脂肪系数为82.19,总平均亲水性为-0.016;该蛋白包含了除Pyl 和Sec 两个氨基酸之外的其余18种常见氨基酸,其中Gly 含量最高,为13.1%,Met和Trp 含量最低,均为0.6%,其他氨基酸含量在1.9%~9.4%;带正电荷氨基酸残基总数为14 个,带负电荷氨基酸残基总数为18 个。利用ExPasy的ProtScale 程序计算基于K-D 法的蛋白质疏水性,结果未发现典型疏水区域。使用TMHMM Server v.2.0进行跨膜区分析,结果未发现典型跨膜区域。使用Signal P 3.0 server 进行信号肽预测,结果显示此蛋白不存在信号肽,不是分泌蛋白。二级结构预测显示ORFV 007编码蛋白主要由无规卷曲和延伸链构成,其中无规卷曲含量最多,为60.62%,位于1-7、14-33、35-46、57-61、67-71、80-82、86-90、99-103、107-110、120-124、130-154、160 区域;延伸链结构占蛋白质的33.12%,位于8-13、34、47-56、62-66、72-79、83-85、91-98、104-106、125-128、155-159区域;除上述结构外,还有少部分α螺旋结构,占蛋白质的5.62%,位于111-119 区域。CBS 在线预测显示,此段蛋白质存在12 个磷酸化位点(丝氨酸9 个、苏氨酸2个、酪氨酸1个);在第17位氨基酸位置,含有一个潜在的N-连接糖基化位点;NCBI 在线预测显示,此蛋白质保守域活性位点的保守特征由9个氨基酸残基组成,这9个残基分别位于65-75、80-95区域;此段蛋白三聚体界面的保守特征由29 个氨基酸残基构成,分别位于25-50 和80-115 区域;dUTPase 蛋白氨基酸序列中包含五个保守结构域,分别是PHA02703、dut、dUTPase、Dut、trimeric_dUTPase。用SWISS-MODEL 预 测该蛋白三级结构(图4),QMQE为0.77,QMEAN为0.31,与模板3ehw.1.A 氨基酸序列的相似性为70.21%,属于dUTP焦磷酸酶。

图4 dUTPase蛋白三维结构预测图
3 讨论与结论
3.1 讨论 dUTPase 广泛存在于各类哺乳动物、昆虫、植物、细菌以及病毒内,对生物体生命发展有重要意义。ORFV dUTPase 是dUTPase 超家族中的常见成员,与参与催化dUTP转化为dUMP过程中的其他超家族成员相比,dUTPase 与它们具有相同的四级结构,但在合成dTTP 过程中,仅有dUTPase 可以通过转储制作将dUTP 转化为dUMP[9],并且还能消除过量dUTP,从而降低dUTP与dTTP比例,进一步降低尿嘧啶的DNA合成过程,减少在合成过程中出现错配[7]。除上述已经确定的功能外,已有研究暗示dUTPase 可调节病毒毒力和基因组完整性[10];亦有研究表明病毒dUTPase 在调节宿主免疫中起着重要作用,可促进Ι型干扰素(IFN)的生成[11]。
3.2 结论 本试验以山羊口疮病毒四川分离株SC-LZ1 的DNA 为模板,采用自行设计的ORFV dUTPase 特异性引物,经PCR 成功扩增获得dUTPase 完整基因,通过构建pET-32a(+)-007重组表达质粒并测序分析,发现该基因全序列与NCBI 收录的编号为MF4890951 的基因同源性最高,为98.96%;构建的系统进化树显示该分离毒株 与 MF489089.1、MF489095.1、MF489093.1、MF489088.1、MF489090.1 及MF489094.1 在同一分支,说明亲缘关系较近,但其又单独为一小支,存在一定的变异。利用各类生物软件对该蛋白结构进行分析,结果编码蛋白中Gly含量最高,且二级结构中无规卷曲所占比例最大;在蛋白质二级结构中,就结构稳定性而言,α螺旋>β折叠>无规卷曲,但就功能而言,无规卷曲往往充当蛋白质和酶的活性中心,综合对蛋白活性位点的预测分析,推测其酶活性中心位于氨基酸序列65-75及80-95;疏水性区域预测结果显示此段氨基酸无疏水性区域,氨基酸疏水性预测值越大则此段氨基酸含有跨膜区域的可能性越大,跨膜区预测结果也显示此段氨基酸无跨膜区域,所以跨膜区分析结果与疏水性区域分析结果一致,进一步证明该蛋白不是细胞信号传导有关的膜受体蛋白。
猜你喜欢
今日农业(2019年10期)2019-06-26
食品科学(2018年10期)2018-05-23
山西医科大学学报(2017年11期)2017-12-01
食品研究与开发(2016年9期)2016-06-13
哈尔滨商业大学学报(自然科学版)(2016年1期)2016-04-22
西南医科大学学报(2015年1期)2015-08-22
农村百事通(2015年8期)2015-05-19
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
云南畜牧兽医(2014年5期)2014-02-28
