
田蓟苷对大鼠A7R5 血管平滑肌细胞增殖、迁移、表型转化及Notch1信号通路调控的影响
2024-03-25 06:13:42李雅琪马晓莉赵云丽王新春
中成药 2024年3期
李雅琪,马晓莉,赵云丽,李 静,袁 勇,王新春*
(1.石河子大学药学院,新疆 石河子 832002; 2.石河子大学医学院第一附属医院,新疆 石河子 832008)
血管平滑肌细胞(vascular smooth muscle cell,VSMCs)是主动脉壁中膜的重要细胞成分[1],血管损伤后,由收缩型转为合成型,即VSMCs 的表型转化,表型转化的特征体现为细胞异常增殖和迁移。VSMCs 在动脉内膜聚集是动脉粥样硬化(atherosclerosis,AS) 过程的基础,在AS 的发生发展过程中,VSMCs 表型转化失调,由血管中膜向内膜迁移并过度增殖[2],分泌更多的细胞外基质和炎症因子[3]。由此可见,VSMCs 表型转化是动脉粥样硬化的关键性起始步骤。Notch 信号通路在VSMCs 分化、增殖、迁移及表型转化中是一个关键的“调节器”,经典的Notch1 信号通路通过激活下游靶基因Hes1、Hes5[4-5],影响细胞增殖、凋亡和分化[6]。
田蓟苷是新疆特色植物香青兰DracocephalumMoldavica L.黄酮类化合物中含量最高的活性单体[7]。课题组前期研究发现,田蓟苷可调节脂质代谢,减少炎症介质生成,对AS 具有保护作用[8-10]。VSMCs 在AS 斑块形成的发展中起着至关重要的作用,而田蓟苷发挥抗AS 的作用与VSMCs表型转化之间的关系尚不清楚。因此,本研究采用氧化低密度脂蛋白(oxidized low density lipoprotein,ox-LDL) 诱导刺激VSMCs 表型转化,探讨田蓟苷调控Notch1 信号通路对VSMCs 表型转化的影响,为田蓟苷抗AS 提供理论依据。
1 材料
1.1 细胞 大鼠A7R5 血管平滑肌细胞,货号FH0419,购自上海富衡生物科技有限公司。
1.2 药物 田蓟苷(纯度大于98%),由新疆药物研究所提供,称取5 mg 田蓟苷粉末,加入到1 mL DMEM 中,配制成5 mg/mL 母液,给药时用含10%胎牛血清的DMEM 培养基稀释到所需剂量。γ-分泌酶抑制剂DAPT (美国APExBIO 公司,批号A8200); 氧化低密度脂蛋白 (ox-LDL,广州弈源生物科技有限公司,批号YB-002); 辛伐他汀(北京索莱宝科技有限公司,批号IS0170),临用时均用含10%胎牛血清的DMEM 培养基稀释至所需剂量。
1.3 试剂 0.1% 结晶紫水溶液、BCA 蛋白定量试剂盒(北京索莱宝科技有限公司,批号G1064、PC0020); FBS胎牛血清(美国HyClone 公司,批号10100147); DMEM 高糖液体培养基(美国Gibco 公司,批号11965092); 0.25%胰酶-EDTA 消化液、ECL 超敏发光试剂盒、cDNA 反转录试剂盒(美国Thermo Fisher Scientific 公司,批号25200-056、P0018A、K1622); TRIzol 试剂 (美国Invitrogen 公司,批号15596026); CCK-8 试剂盒(日本DojinDO 公司,批号CK04); Transwell 聚碳酸酯膜嵌套(美国Corning 公司,批号3422); QuantiNova SYBR Green PCR Kit (德国QIANGEN 公司,批号208054); Hes1 兔单克隆抗体、RBPJκ 兔单克隆抗体(英国Abcam 公司,批号ab108937、ab180588); Notch1 兔单克隆抗体 (美国Cell Signaling Technology 公司,批号3608S); GAPDH 小鼠单克隆抗体、HRP 标记山羊抗小鼠IgG、HRP 标记山羊抗兔IgG (北京中杉金桥生物技术有限公司,批号10494-1-AP、140193、141987)。
1.4 仪器 Forma II 3110 水套式CO2培养箱、Pico-17 型微量高速离心机 (美国Thermo Fisher Scientific 公司);CKX53 型常规倒置显微镜 (日本Olympus 公司); DW-86L100J-80 ℃超低温冰箱(青岛海尔电冰箱有限公司);M200 Pro 型多功能酶标仪(瑞士Tecan 公司); VE-180 型垂直电泳及电转仪(上海天能科技有限公司); ChemiDoc XRS+型凝胶成像系统(美国Bio-Rad 公司); Rotor-Gene Q型实时荧光定量-聚合酶链式反应 (PCR) 仪 (德国QIANGEN 公司)。
2 方法
2.1 细胞培养 将A7R5 血管平滑肌细胞接种于含10%胎牛血清、0.1% 青-链霉素的DMEM 高糖培养基,置于37 ℃、5% CO2培养箱中培养,细胞生长融合时用0.25%胰酶-EDTA 消化液传代,传代后取对数生长期细胞进行实验。
2.2 造模及分组 将A7R5 细胞分为对照组(未加药物处理,用含10% 胎牛血清的DMEM 培养基进行常规培养)、模型组(50 μg/mL ox-LDL 作用24 h 后,等体积含10%胎牛血清的DMEM 培养基作用24 h)、田蓟苷组(50 μg/mL ox-LDL 作用24 h 后,10 μg/mL 田蓟苷作用24 h)、DAPT组(50 μg/mL ox-LDL 作用24 h 后,10 μmol/L DAPT 作用24 h)、辛伐他汀组(50 μg/mL ox-LDL 作用24 h 后,10 μmol/L 辛伐他汀作用24 h)。
2.3 CCK8 法检测细胞活性 将A7R5 细胞接种到96 孔板中,按照“2.2” 项下分组进行处理,分别测定0 (细胞贴壁后不做任何处理)、24 (给予ox-LDL 造模24 h)、48 h(ox-LDL 造模24 h+各组药物24 h) 细胞活性,每组6 个复孔。处理结束后每孔加入含10% CCK-8 试剂的无血清培养基100 μL,在37 ℃培养箱中孵育1.5 h,在450 nm 波长下测定光密度(OD) 值,细胞增殖活性与OD 值成正比。
2.4 Transwell 法检测细胞迁移数 将A7R5 细胞接种于6孔板中,按照“2.2” 项下分组进行处理,收集各组细胞于无血清培养基中,铺入Transwell 小室的上室中,将小室放入24 孔板,下室加入600 μL 含15% FBS 的培养基,培养24 h后,取出小室,弃去孔中培养液,PBS 浸洗2 次后,用棉签轻轻擦掉上层未迁移细胞,甲醇固定30 min,适当风干,0.1%结晶紫溶液染色40 min,PBS 浸洗3 次后,棉签轻轻拭去上室水分,将上室的纤维膜撕下,置于载玻片上,封于中性树胶中,盖上盖玻片晾干,于200 倍显微镜下随机选5 个视野观察细胞,拍照,计数。
2.5 RT-qPCR 法检测细胞SM22α、α-SMA、Notch1、Hes1、Hes5、Jagged-1 mRNA 表达 将A7R5 细胞接种于6 孔板中,按照“2.2” 项下分组进行处理,收集各组细胞,采用TRIzol 法提取细胞总RNA,测定其浓度和纯度后,使用逆转录试剂盒逆转录为cDNA,按试剂盒说明书进行扩增反应,以GAPDH为内参,结果以2-ΔΔCT法计算目的基因mRNA 相对表达量。引物由生工生物工程(上海) 股份有限公司合成,序列见表1。

表1 引物序列
2.6 Western blot 法检测细胞Notch1、RBP-Jκ、Hes1 蛋白表达 将A7R5 细胞接种于6 孔板中,按照“2.2” 项下分组进行处理,弃掉培养基,PBS 洗3 次后,向各组孔中加入150 μL 预冷的含1%PMSF 的裂解液,冰上裂解15 min,4 ℃、12 000 r/min 离心10 min,取上清得到细胞总蛋白溶液,BCA 测定蛋白浓度。经10% SDS-PAGE 凝胶电泳,湿法转膜,5%脱脂奶粉溶液封闭1 h,然后分别加入一抗Notch1 (1 ∶1 000)、RBP-Jκ (1 ∶ 1 000)、Hes-1 (1 ∶500)、GAPDH (1 ∶4 000) 4 ℃孵育过夜,TBST 洗膜4次,加入HRP 标记山羊抗小鼠IgG 或抗兔IgG 二抗(1 ∶20 000) 室温孵育1 h,TBST 洗膜4 次,ECL 化学发光液法显影,通过Image J 1.6.0 软件对条带进行灰度值分析。
2.7 统计学分析 采用SPSS 26.0 软件进行处理,数据以(±s) 表示,组间比较采用单因素方差分析。P<0.05 为差异有统计学意义。
3 结果
3.1 田蓟苷对A7R5 细胞增殖的影响 与对照组比较,模型组细胞增殖能力增强(P<0.01); 与模型组比较,田蓟苷组、DAPT 组和辛伐他汀组细胞增殖能力减弱 (P<0.01),见表2。
表2 田蓟苷对A7R5 细胞增殖的影响(±s,n=6)
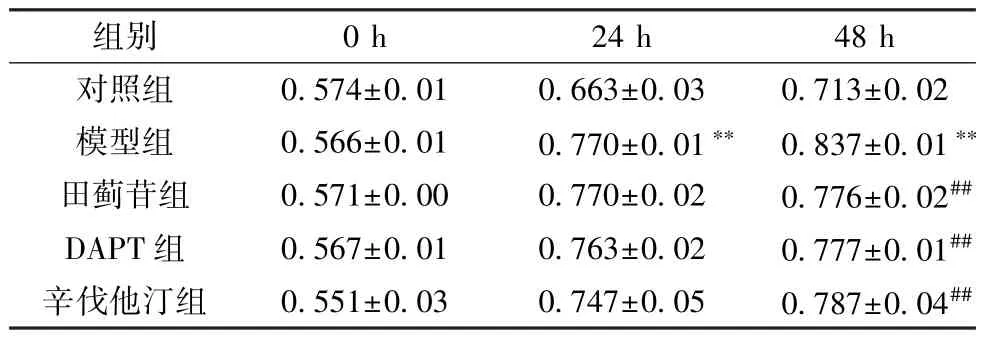
表2 田蓟苷对A7R5 细胞增殖的影响(±s,n=6)
注: 与对照组比较,**P<0.01; 与模型组比较,##P<0.05。
组别0 h24 h48 h对照组0.574±0.010.663±0.030.713±0.02模型组0.566±0.010.770±0.01** 0.837±0.01**田蓟苷组0.571±0.000.770±0.020.776±0.02##DAPT 组0.567±0.010.763±0.020.777±0.01##辛伐他汀组0.551±0.030.747±0.050.787±0.04##
3.2 田蓟苷对A7R5 细胞迁移的影响 与对照组比较,模型组细胞迁移能力增强(P<0.05); 与模型组比较,田蓟苷组、DAPT 组和辛伐他汀组细胞迁移能力减弱(P<0.05),见图1。

图1 田蓟苷对A7R5 细胞迁移的影响(×200,±s,n=5)
3.3 田蓟苷对A7R5 细胞SM22α、α-SMAmRNA 表达的影响 与对照组比较,模型组细胞α-SMAmRNA 表达降低(P<0.01),SM22αmRNA 表达呈下降趋势,但差异不显著(P>0.05); 与模型组比较,田蓟苷组、DAPT 组和辛伐他汀组细胞α-SMAmRNA 表达均升高 (P<0.01),SM22αmRNA 表达呈增加趋势,但差异不显著 (P>0.05),见表3。
表3 田蓟苷对A7R5 细胞SM22α、α-SMA mRNA 表达的影响(±s,n=5)
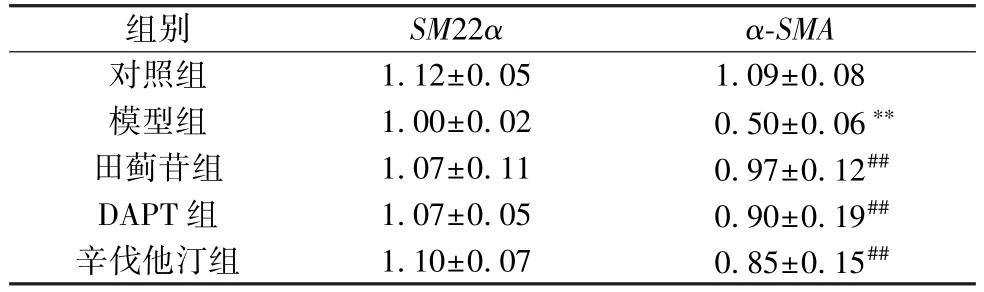
表3 田蓟苷对A7R5 细胞SM22α、α-SMA mRNA 表达的影响(±s,n=5)
注: 与对照组比较,**P<0.01; 与模型组比较,##P<0.05。
组别SM22αα-SMA对照组1.12±0.051.09±0.08模型组1.00±0.020.50±0.06**田蓟苷组1.07±0.110.97±0.12##DAPT 组1.07±0.050.90±0.19##辛伐他汀组1.10±0.070.85±0.15##
3.4 田蓟苷对A7R5 细胞Notch1、Hes1、Hes5、Jagged-1 mRNA 表达的影响 与对照组比较,模型组细胞Notch1、Hes1、Hes5 和Jagged-1 mRNA 表达升高(P<0.01); 与模型组比较,田蓟苷组和DAPT 组细胞Notch1、Hes1、Hes5、Jagged-1 mRNA 表达降低(P<0.05,P<0.01),辛伐他汀组Hes1、Jagged-1 mRNA 表达降低(P<0.05,P<0.01),Notch1、Hes5 mRNA 表达无明显变化(P>0.05),见表4。
表4 田蓟苷对A7R5 细胞Notch1、Hes1、Hes5、Jagged-1 mRNA 表达的影响(±s,n=5)

表4 田蓟苷对A7R5 细胞Notch1、Hes1、Hes5、Jagged-1 mRNA 表达的影响(±s,n=5)
注: 与对照组比较,**P<0.01; 与模型组比较,#P<0.05,##P<0.05。
组别Notch1Hes1Hes5Jagged-1对照组0.61±0.070.63±0.060.53±0.100.60±0.06模型组1.02±0.25**1.04±0.07**1.01±0.17**1.01±0.14**田蓟苷组0.64±0.17##0.82±0.15#0.59±0.04##0.64±0.09##DAPT 组0.61±0.03##0.70±0.11##0.45±0.22##0.39±0.10##辛伐他汀组0.89±0.170.71±0.07##0.76±0.250.68±0.08#
3.5 田蓟苷对A7R5 细胞Notch1、RBP-Jκ、Hes1 蛋白表达的影响 与对照组比较,模型组细胞Notch1、RBP-Jκ、Hes1 蛋白表达升高(P<0.05,P<0.01); 与模型组比较,田蓟苷组细胞Notch1、RBP-Jκ、Hes1 蛋白表达降低(P<0.05,P<0.01),DAPT 组和辛伐他汀组Hes1、RBP-Jκ蛋白表达降低(P<0.05,P<0.01),Notch1 蛋白表达无明显变化(P>0.05),见图2。
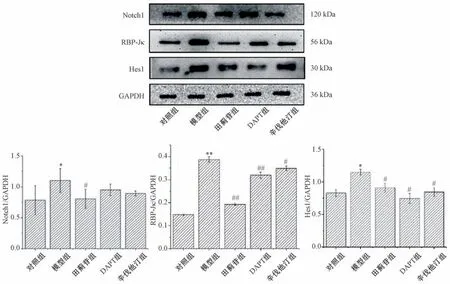
图2 田蓟苷对A7R5 细胞Notch1、RBP-Jκ 和Hes1 蛋白表达的影响(±s,n=5)
4 讨论
VSMCs 位于血管中膜上,在正常状态下,VSMCs 表现为高度分化的收缩型,增殖和迁移能力较差; 当血管受损或ox-LDL 刺激时,VSMCs 由收缩型转化为异常增殖的合成型,此过程称为表型转化。合成型的VSMCs 迁移到内膜吞噬胆固醇,导致脂质积累和泡沫细胞形成[11],斑块稳定性下降,进一步加速AS 的发展。因此,控制VSMCs 表型转化对防治AS 具有重要意义。
本研究CCK-8 和Transwell 结果表明,田蓟苷可抑制ox-LDL 诱导的细胞异常增殖和迁移。α-SMA 和SM22α 作为收缩型VSMCs 的标志[12-13],当细胞处于增殖、修复或病变状态时,该基因表达减少。本研究结果显示,ox-LDL 刺激使α-SMAmRNA 表达降低,表明VSMCs 向合成型转化,田蓟苷处理可上调α-SMAmRNA 表达,抑制ox-LDL 诱导的VSMCs 的表型转化。SM22αmRNA 表达无明显变化,可能是因为SM22α 表达具有时空性特点,在VSMCs 发育过程中,SM22α 的表达晚于α-SMA[14-15]; 并且平滑肌SM22α 表达的激活有从量变到质变的过程,表现出时间、剂量储积性[16-17]。以上结果表明,田蓟苷可抑制VSMCs 的异常增殖、迁移和表型转化。
Notch 信号通路由Notch 配体、受体、下游信号转导分子和核内应答过程组成,与细胞的增殖和分化密切相关[18]。其中,受体Notch1 被广泛关注,Notch1 信号通路能调控平滑肌细胞的分化[19-21],配体Jaggde1 在平滑肌细胞成熟的过程中发挥作用[22],Hes1 和Hes5 是下游靶基因。Notch 信号通路在生物进化过程中高度保守,组成部分的缺失会引起生理功能障碍,为了保证通路的完整性,本研究测定了包括受体Notch1、配体Jaggde1、靶基因Hes1 和Hes5 在内的通路基因。结果显示,ox-LDL 诱导A7R5 细胞表型转化后,Notch1 通路相关基因Notch1、Hes1、Hes5、Jagged-1 mRNA 和Notch1、RBP-Jκ、Hes1 蛋白表达均上调,而田蓟苷处理可逆转表达的上调,且在基因水平的作用与DAPT 相当。值得注意的是,DAPT 抑制RBP-Jκ、Hes1 蛋白表达,对Notch1 蛋白的抑制作用不明显,其原因可能是由于DAPT 是γ-分泌酶抑制剂[23],而Notch1 信号通路的转导途径是Notch1 与配体结合形成胞内域,胞内域在γ-分泌酶的作用下被裂解、释放,活化的胞内域易位到细胞核与RBP-Jκ 结合形成复合物,进而激活下游靶蛋白Hes1 和Hes5 的表达[20,24],因此DAPT 对单独的Notch1 蛋白的抑制作用有限,而不影响对下游的RBP-Jκ、Hes-1 蛋白的抑制作用。鉴于DAPT 对Notch1 通路中胞内域的这种抑制过程,故而在蛋白表达层面测定通路蛋白Notch1,形成复合物的关键蛋白RBP-Jκ 和下游靶蛋白Hes-1。
综上所述,田蓟苷对ox-LDL 诱导的A7R5 细胞有确切的保护作用,其机制与抑制VSMCs 的异常增殖、迁移、表型转化,调控Notch1 信号通路有关。然而,对于田蓟苷在体内是否具有同样的作用机制还有待进一步的研究。
猜你喜欢
航天电子对抗(2022年4期)2022-10-24 13:38:30
中国科技纵横(2018年2期)2018-11-29 18:45:44
中成药(2018年7期)2018-08-04 06:04:26
现代园艺(2017年21期)2018-01-03 06:41:32
中国卫生标准管理(2015年17期)2016-01-20 09:26:45
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
医学研究杂志(2015年5期)2015-06-10 06:43:26
中国当代医药(2015年17期)2015-03-01 02:03:39
现代检验医学杂志(2015年5期)2015-02-06 01:42:20
中国卫生标准管理(2015年13期)2015-01-26 21:05:38
