
铁死亡机制及其在食管癌中的作用研究*
2024-03-18 00:38:30李析濛唐月王涛关徐涛李松伟
中医学报 2024年3期
李析濛,唐月,王涛,关徐涛,李松伟,3
1.河南中医药大学,河南 郑州 450046; 2.河南中医药大学第一附属医院,河南 郑州 450000;3.河南中医药大学第二附属医院/河南省中医院,河南 郑州 450002
食管癌是临床常见的消化系统肿瘤,据统计,其在全球的发病率和死亡率分别位于各种恶性肿瘤的第8位和第6位,且我国中北部是食管癌的高发地区,发病数约占全世界的50%[1]。食管癌主要包括食管腺癌(esophageal adenocarcinoma,EAC)和食管鳞状细胞癌(esophageal squamous cell carcinoma,ESCC)两个亚型,其中ESCC是最常见的组织亚型[2]。食管癌临床常见的治疗方式包括单纯手术治疗和新辅助化学治疗,但手术治疗容易伴发食道梗阻和狭窄等并发症,新辅助化学治疗容易出现肿瘤细胞的化疗耐药,治疗效果并不理想[3]。因此,积极寻求治疗食管癌的新的有效途径显得尤为必要。近年来,随着科学研究的深入,铁死亡(ferroptosis)作为一种新型的对铁离子依赖的细胞程序性死亡方式,越来越受到关注[4]。研究发现,铁死亡与癌症的发生、发展及病情转归关系密切[5]。铁死亡的激活可以对肿瘤细胞的增殖产生抑制,可能为肿瘤防治提供新的方向。本文对铁死亡的发生机制及其与食管癌疾病进展的关系进行综述。
1 铁死亡概述
2003年,Dolma等[6]发现一种名为Erastin的小分子化合物,能够以一种非凋亡的模式选择性杀死大鼠肉瘤病毒(rat sarcoma virus,RAS)基因突变的肿瘤细胞,而且此种死亡模式没有出现细胞核的形态改变、DNA的断裂以及半胱天冬酶(cysteinyl aspartate specific proteinase,Caspase)激活,caspase抑制剂对Erastin诱导的细胞死亡亦无显著抑制作用,而脂质过氧化物抑制剂和铁螯合剂的使用却能发挥明显的抑制作用。这一研究结果表明,在经典细胞凋亡模式外还存在一种全新的细胞死亡形式。2012年,Dixon等[4]首次将这种由Erastin所介导的新型调节性细胞死亡方式命名为铁死亡。铁死亡的主要特点为铁依赖的高活性氧(reactive oxygen species,ROS)积累和脂质过氧化物(lipid peroxide,LPO)堆积,这使细胞原有的氧化还原平衡遭到破坏,最终导致细胞死亡。铁死亡在细胞学上表现为细胞膜丧失完整性,线粒体变小甚至消失,以及其外膜的破裂。
2 铁死亡的调控机制
细胞铁死亡的过程主要包括3个重要环节:铁超载、LPO累积以及谷胱甘肽过氧化物酶4(glutathione peroxidase 4,GPX4)活性下调。见图1。越来越多的研究表明,铁代谢异常、脂质代谢异常和氨基酸代谢异常是诱发铁死亡的关键因素,本篇主要从这三方面对铁死亡的调节机制进行概述。
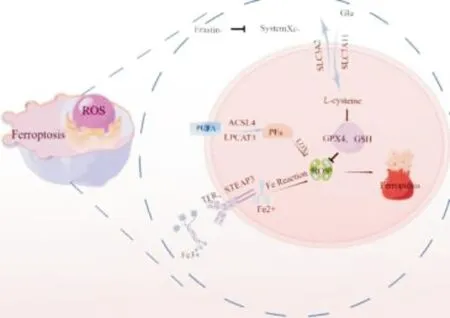
图1 铁死亡调控机制示意图
2.1 铁代谢细胞铁是诱导铁死亡发生的关键。铁在人体中主要以两种形式出现,即Fe3+和Fe2+。食物中的Fe3+在胃肠道被还原为Fe2+,Fe2+在十二指肠和空肠上段被吸收。人体循环中的铁主要是Fe3+,Fe3+与细胞膜转铁蛋白受体1(transferrin receptor 1,TfR1)结合后进入细胞,在细胞内铁还原酶金属还原酶家族3(six-transmembrane epithelial antigen of the prostate 3,STEAP3)的作用下被还原为Fe2+。二价金属转运蛋白1(divalent metal transporter 1,DMT1)将一部分Fe2+释放到细胞质的动态铁池中,余下的Fe2+由膜铁转运蛋白氧化为Fe3+出胞,继续参与机体内铁的再循环[7-8]。正常生理情况下,动态铁池可以维持机体铁代谢的稳定状态。当出现铁超载现象时,血清转铁蛋白接近饱和,Fe2+在细胞内积累,与H2O2反应生成羟自由基,即芬顿反应,使细胞内ROS含量升高,ROS与细胞膜上多不饱和脂肪酸(polyunsaturated fatty acids,PUFAs)发生过氧化反应,导致细胞膜结构被破坏,从而促进细胞铁死亡[9-10]。研究发现,在RAS基因突变的铁死亡敏感细胞中,TfR1表达增加和铁蛋白表达降低是导致铁超载的主要原因,铁螯合剂脱铁胺可以通过抑制铁超载从而阻碍铁死亡,而供应外源性铁可刺激铁死亡,沉默TFR1或使用铁螯合剂脱铁胺可逆转Erastin诱导的细胞铁死亡[11-12]。研究还发现,核转录因子红细胞系相关因子-2(nuclear factor-erythroid 2-related factor 2,Nrf2)、热休克蛋白B1(heat shock protein β1,HSBP1)等基因可通过限制 ROS 产生和下调TFR1介导的铁摄取过程,从而缓解铁过载并抑制细胞铁死亡,进而对铁死亡起到负性调节作用[13-14]。铁反应元件结合蛋白2(iron-responsive element-binding protein 2,IREB2)是影响铁代谢的主要转录调节因子,IREB2基因沉默后铁蛋白受刺激而表达增加,导致细胞内铁含量下降,进而减弱了erastin诱导的铁死亡[4]。由此可见,对细胞铁的摄取、贮存和利用进行调控,是铁死亡调控的关键环节。
2.2 脂质代谢研究表明,LPO异常聚集是铁死亡发生的中心环节,铁死亡是PUFAs的磷脂过度氧化所引起的细胞死亡,PUFAs是铁死亡中LPO积累的关键物质[15]。PUFAs是组成细胞膜脂质双分子层的重要成分之一,是铁死亡脂质过氧化的主要底物,其极易受到ROS损伤,产生强烈的氧化应激反应[16]。细胞内PUFAs的含量积聚程度将影响细胞脂质过氧化的进度,进而影响着铁死亡发生的敏感性[17-18]。因此,脂质合成代谢异常也会间接地影响铁死亡。研究发现PUFAs能在乙酰辅酶A 合成酶长链家族成员4(acyl-CoA synthetase long-chain family member 4,ACSL4)和溶血磷脂酰胆碱酰基转移酶3(lysophosphatidylcholine acyltransferase 3,LPCAT3)这两个关键酶的共同作用下生成磷脂酰乙醇胺(phosphatidylethanolamines,PEs),PEs是诱发细胞铁死亡的关键磷脂,可继而被一种含铁的蛋白酶脂加氧酶氧化为LPO,过量的LPO导致细胞膜损伤,最终可触发铁死亡[19-21]。研究人员发现,在多种肿瘤细胞中,ACSL4和LPCAT3表达明显升高,而且其表达水平与这些肿瘤细胞对铁死亡的敏感性呈正相关[22]。同时,ACSL4 和LPCAT3有助于铁死亡过程中细胞内脂质过氧化物的积累,故ACSL4和LPCAT3基因敲除或功能抑制时可降低PUFAs的合成,进而增强细胞对铁死亡的抵抗[18,23]。相反,当PUFAs 的主要代表物花生四烯酸和烟油酸盐含量充足时,铁死亡诱导剂erastin会使过氧化脂质增多,进而加快铁死亡[16]。这些证据均提示脂质代谢异常对铁死亡的发生发挥着关键作用,PUFAs是铁死亡过程中脂质过氧化的重要靶点。
2.3 氨基酸代谢GPX4是一种重要的脂修复酶,也是已知的仅有的能够直接清除LPO的酶,可以将LPO还原为脂质醇,防止LPO堆积[24]。细胞膜上存在胱氨酸/谷氨酸反向转运体(System Xc-),其为轻链SLC7A11和重链SLC3A2共同构成的异二聚体。System Xc-可使胞内的谷氨酸(glutamate)与胞外的胱氨酸(cystine)以11的比例进行交换。胱氨酸进入细胞内被还原为半胱氨酸(L-cysteine),可作为合成谷胱甘肽(glutathione,GSH)的原料。GSH是由谷氨酸、半胱氨酸和甘氨酸所构成的三肽化合物,是重要的抗氧化剂,对自由基的清除和保持细胞内外氧化还原稳态起着重要作用,是GPX4降解LPO的协同分子。当GSH的合成被抑制或GPX4缺乏时,LPO发生蓄积,细胞膜被破坏,从而促进铁死亡的发生[24]。研究证实通过对GPX4的抑制可以诱导小鼠肿瘤异种移植瘤中的铁死亡;同时,GPX4间接失活后,细胞对铁死亡的敏感性会增加[25-26]。由此可见,在铁死亡调控过程中,GPX4发挥着关键作用。当细胞膜上的System Xc-减少或失活时,细胞无法正常摄取足量的胱氨酸,使GSH合成减少,从而促进铁依赖的ROS的积累,进而促进了铁死亡的发生,铁死亡激活剂erastin和柳氮磺吡啶即是通过抑制System Xc-的活性进而发挥作用[27]。研究证实,SLC7A11的表达增加能明显抑制铁死亡的发生,而抑制SLC7A11的表达能显著提高细胞对铁死亡的敏感性[28-29];抑癌基因p53可以通过抑制SLC7A11转录水平的方式抑制System Xc-的功能,从而进一步调控铁死亡[30]。由此可见,在铁死亡的发生发展过程中,system Xc-发挥着重要作用。因此,寻找靶向GSH代谢过程的药物,对于铁死亡的研究意义重大。
3 铁死亡与食管癌
近年来的研究表明,乳腺癌、肾癌、肺癌、肝癌、胶质细胞瘤和前列腺癌等的发生发展都与铁死亡有关,铁死亡在抑制肿瘤生长和杀死肿瘤细胞方面发挥了重要作用[31-32]。铁死亡的产生机制与细胞代谢联系紧密,包括许多关键分子和信号通路,调控这些重要分子和信号通路会改变细胞对铁死亡的敏感度。通过合理的方式诱导或控制铁死亡,将使多种病症得到改善,特别是恶性肿瘤。铁死亡现象最初是在RAS突变的癌细胞中被发现的。大量发生RAS突变的不同类型的癌细胞对铁死亡的诱导较为敏感。研究表明,RAS的激活可以通过激活TfR1和抑制膜铁转运蛋白(FPN)来提高细胞内铁的摄入,同时减少细胞内铁的外排[33-34]。简而言之,可利用对铁代谢这一途径的调节,使肿瘤细胞对铁死亡更为敏感。铁死亡已成为许多癌症治疗药物和手段必不可少的环节,然而,铁死亡与食管癌相关报道仍较少。
3.1 机制方面铁死亡是一种以细胞内铁和ROS积累为特征的非凋亡细胞死亡,是由氧化和抗氧化系统失衡引起的氧化损伤过程。氧化应激在许多癌症发病机制中发挥着重要作用。研究发现,ESCC患者血清中超氧化物歧化酶、谷胱甘肽过氧化物酶、谷胱甘肽还原酶活性水平低于正常健康人群,而血清丙二醛(malonic dialdehyde,MDA)水平显著高于对照组,表明ESCC的发病与氧化应激增加和抗氧化酶活性降低有关[35],提示食管癌的发生发展与铁死亡存在一定的关系。
NRF2是主要的细胞抗氧化和铁代谢调节因子。研究发现,在EC组织和细胞中,NRF2显著表达,有利于肿瘤细胞的生长[36]。同时,在ESCC活检标本中,NRF2的表达与ESCC的化疗效果和预后呈负相关,NRF2高表达的ESCC比NRF2低表达的ESCC更耐放化疗,生存率更低[37-38]。Brusatol是一种靶向NRF2抑制剂,能够显著抑制NRF2转录活性并下调NRF2靶基因,促进脂质过氧化和铁死亡,进而抑制EAC细胞生长,同时增加食管腺癌细胞对顺铂的敏感性[39]。由此进一步表明,NRF2在ESCC中具有促肿瘤作用,探究针对NRF2信号传导的靶向治疗将是NRF2高表达的ESCC治疗的有效举措。
SLC7A11是半胱氨酸/谷氨酸转运蛋白的组成部分,在肿瘤生长中起关键作用。在ESCC中,NRF2作为转录因子可刺激SLC7A11的表达上调,上调的SLC7A11有利于细胞内GSH的合成,进一步增强了对铁死亡的抵抗力[40]。研究表明,ESCC患者中NRF2和SLC7A11在ESCC组织中过度表达,相互呈正相关,并且与较差的生存率和治疗结果有关[40]。SLC7A11介导的铁死亡抑制诱导NRF2相关的放射抗性,突出了 NRF2/SLC7A11/铁死亡轴在未来ESCC治疗中的优势。NRF2调控的SLC7A11可通过抑制铁死亡调节ESCC的放射敏感性[40]。目前,一些SLC7A11抑制剂已被陆续发现,如erastin、柳氮磺吡啶、顺铂等[41],SLC7A11作为EC治疗靶点必将越来越受到重视。
P53是一种重要的抑癌基因,其基因突变或失活可促进细胞分裂、增殖,致使肿瘤形成。P53基因突变在EC中很常见[42-43]。P53可以通过调节氧化代谢干预癌症中的铁死亡[44]。最近的一项研究发现,辐射诱导的P53激活抑制了SLC7A11的表达,导致EC细胞脂质过氧化和铁死亡[45]。
GPX4能分解小分子过氧化物并削弱脂质过氧化物毒性,防止LPO堆积,防止脂质过氧化,抑制铁死亡[27]。HMOX1在铁代谢调节途径中同样起着重要作用。人体内每天所需的铁大部分来自血红素分解形成的铁。HMOX1是参与血红素分解代谢的关键酶,能使血红素分解为CO、Fe2+和胆绿素。HMOX1的激活介导铁离子从血红素中排出,诱导了铁死亡的出现[46]。GPX4的上调和血红素加氧酶(heme oxygenase 1,HMOX1)的下调是ESCC的不良预后因素[47]。
DnaJ/Hsp40同源物B亚家族成员6(DNAJB6)水平与ESCC患者淋巴结转移呈负相关。过表达DNAJB6a在体外和体内均显示出肿瘤抑制作用。此外,DNAJB6a的过度表达伴随着GPX4和磷酸化AKT(p-AKT)蛋白水平的显著降低[47]。研究证实,Zeste同源物2增强子(EZH2)通过miR-125b-5p/SLC7A11轴抑制erastin诱导的ESCC细胞铁死亡,与ESCC患者的不良预后显著相关[48]。DNAJB6EZH2通过铁死亡在ESCC的演变中起重要的抑癌作用。
3.2 治疗方面上述研究证实铁死亡与食管癌的发病关系密切,食管癌细胞铁死亡的发生对食管癌的进展有抑制作用,诱导食管癌细胞铁死亡对食管癌的防治意义重大。目前研究表明,一些铁死亡诱导剂似乎是可行的食管癌治疗手段。
最近,一种胶质毒素相关的铁死亡途径引起学者们的关注。胶质毒素是从海洋来源真菌烟曲霉中分离出来的次生代谢产物,已在多种癌症中显示出抗肿瘤特性。SUV39H1是胶质毒素靶基因,在EC组织中的表达水平高于正常组织,而在ESCC患者中的高表达水平表明预后不良。研究发现,SUV39H1通过影响铁代谢和脂质过氧化参与铁死亡过程,敲除或者抑制SUV39H1可以诱导铁聚集和脂质过氧化,引发铁死亡,进而导致癌细胞破坏[49]。研究证实,胶质毒素可通过下调ESCC细胞中SUV39H1的表达,诱导铁死亡,进而发挥对ESSC的抗肿瘤特性[50]。
富含亮氨酸胶质瘤失活蛋白1 (leucine rich glioma inactivated 1,LGI1) 是一种神经元分泌蛋白,去整合素-金属蛋白酶 23 (adisintegrin and metalloproteinase 23,ADAM23) 是其作用受体。相关性分析显示,ADAM23的表达与铁死亡相关基因GPX4、SLC3A2和SLC7A11显著相关,过表达的ADAM23通过抑制GPX4、SLC3A2和SLC7A11促进铁死亡。研究发现,LGI1可通过作用于ADAM23促进铁死亡,进而抑制ESCC细胞的增殖和迁移[51]。
5-氨基乙酰丙酸(5-aminolevulinic acid,5-ALA)是一种广泛用于癌症治疗的天然氨基酸,能通过GPX4和HMOX1诱导铁死亡,并在ESCC中具有抗肿瘤作用[52]。该团队证实5-ALA对ESCC有缩小肿瘤的作用,且可抑制肿瘤中GPX4的表达,并上调HMOX1的表达。铁死亡最重要的上游调节因子是GPX4,抑制GPX4可诱导肿瘤细胞中的铁死亡。目前,5-ALA已被临床应用于皮肤癌的光动力疗法和膀胱癌的光动力诊断,并被证明是安全的[53-54]。研究表明,5-ALA对ESCC细胞株的GPX4和HMOX1具有调节作用,可诱导铁死亡,发挥抗肿瘤效果[52]。GPX4和HMOX1这两个分子很可能是治疗ESCC的潜在靶点,5-ALA通过对二者的调节作用或将成为一种治疗ESCC的新药。
癌干细胞是导致肿瘤发生和治疗失败的癌细胞亚群,而Hsp27在保护癌干细胞免受铁死亡的影响中发挥重要作用。研究发现,食管癌干细胞铁水平升高,脂质过氧化,而Hsp27可通过抑制P53上调xCT-GPX4,以保护癌干细胞免受铁死亡,说明Hsp27和GPX4是食管癌患者重要的预后指标,靶向 Hsp27 或GPX4来阻断这种针对铁死亡的内在保护机制可作为ESCC中癌干细胞的潜在治疗策略[55]。
克服对辐射的抵抗力是癌症治疗的主要挑战。研究表明,硬脂酰辅酶A去饱和酶(stearoyl-CoA desaturase-1,SCD1)是负责油酸和棕榈烯酸形成的酶,SCD1的高表达与ESCC患者的不良生存率相关,靶向SCD1能够诱导铁死亡和免疫原性细胞死亡,从而提高ESCC的放射敏感性。SCD1可被视为ESCC生存的重要预后指标[56]。
有学者对铁死亡相关基因表达情况与ESCC预后之间的关联也做了进一步的研究,发现了一些关联程度较高的基因,在预测ESCC预后方面具有潜在的临床应用价值,为个性化治疗提供新的选择[57-58]。另外,铁死亡免疫治疗在EC中的运用也被报道,但其机制还有待探索[59]。
3.3 中医药方面目前,中医药被视为抗肿瘤药物的重要组成部分,近年来研究发现中药在抑制食管癌癌细胞增殖、促进细胞凋亡等方面发挥积极作用,与化学合成药物相比毒副作用少、耐药性小、远期疗效好[60]。部分中药含有的多种活性成分通过诱导肿瘤细胞铁死亡发挥了显著的抗癌功效。
冬凌草甲素是传统中药冬凌草的主要活性成分,是一种天然的四环二萜类化合物,研究发现冬凌草甲素除了具有降压、抗氧化、增强机体免疫力等功效外,还具有良好的抗肿瘤活性[61]。Zhang等[62]研究证实,经冬凌草甲素处理ESCC细胞TE1后,细胞内铁离子、MDA水平和ROS呈显著的剂量依赖性,出现脂质过氧化、细胞增殖抑制和细胞死亡,这一过程能够被特定的铁死亡抑制剂阻断。冬凌草甲素通过抑制TE1细胞中的γ-谷氨酰循环,能够使细胞内GSH/GSSG值降低,导致GPX4的酶活性显著降低,进而诱导铁死亡,发挥对食管癌细胞TE1的抗癌活性。
雄黄是一种含有砷化合物的中药。研究证实,雄黄具有镇痛、消炎、提高免疫力等作用,且对肿瘤细胞增殖、迁移和侵袭具有抑制效果,是一种潜在的抗肿瘤药物,已被用于治疗各种类型的肿瘤,并取得了积极的效果[63]。然而,雄黄对EC的干预效果尚未完全明确。研究发现,中药雄黄可显著抑制EC细胞系Eca109和KYSE150细胞的增殖,还可显著抑制Eca109和KYSE150细胞的迁移和侵袭,并影响p62、Keap1和Nrf2的mRNA和蛋白表达。作为对雄黄的反应,低p62表达抑制了Eca109和KYSE150细胞的增殖、迁移和侵袭,诱导铁死亡。结果表明中药雄黄可通过激活铁死亡,抑制p62-Keap1-Nrf2途径,进而诱导EC细胞铁死亡并抑制其迁移[64]。
4 结语与展望
铁死亡作为一种新型非程序性细胞死亡方式,已被证实与多种癌症的病理生理过程密切相关。通过铁死亡途径干预食管癌细胞的增殖为食管癌的精准治疗提供了新方法,具有一定的临床应用价值。然而由于铁死亡途径的调节通路较多,食管癌的发生发展亦有其复杂性,所以目前铁死亡在食管癌中的作用机制尚未完全明确。近年来,铁死亡在食管癌的相关研究中已经取得了重大进展,但仍有很多问题亟待解决。例如,在食管癌中,铁死亡与其他细胞死亡形式之间是否存在相互调控作用还有待评价;铁死亡对食管癌的具体干预阶段及靶点还需要探索;对一些预测食管癌的铁死亡相关基因的调控机制研究得还不够深入,与病情严重程度是否有关联也未探及;目前的研究多是理论或实验研究,真正把铁死亡理论用于食管癌临床治疗的还很少,研究证实使用胶质毒素、LGI1、5-ALA等能够抑制食管癌细胞的增殖和迁移,但确切机制仍有待进一步探索;目前已被证实的铁死亡诱导剂如柳氮磺吡啶、索拉菲尼等,还有待临床验证。下一步从代谢、免疫和遗传学的角度更全面地揭示和分析铁死亡在食管癌发病中的作用机制,探寻能够反映食管癌细胞对铁死亡敏感的生物标记物和治疗靶点,发现新的铁死亡诱导剂并进行临床评价,对提高食管癌诊治水平关系意义重大。中药干预铁死亡治疗食管癌的探索目前还处在起步阶段,需要进一步研究。铁死亡将成为治疗食管癌的一种新策略。
猜你喜欢
现代临床医学(2021年6期)2021-11-20 06:34:50
中成药(2018年9期)2018-10-09 07:18:36
中成药(2018年1期)2018-02-02 07:19:53
中成药(2017年12期)2018-01-19 02:06:48
中成药(2017年4期)2017-05-17 06:09:26
西南军医(2016年1期)2016-01-23 02:22:29
中国继续医学教育(2015年4期)2016-01-07 07:38:06
川北医学院学报(2015年5期)2015-12-05 08:22:43
中国洗涤用品工业(2015年4期)2015-02-28 19:02:15
中成药(2014年9期)2014-02-28 22:28:50
