
柑橘采后青霉菌的侵染及产毒机制的研究进展
2024-03-14 08:03:34黄欣娅戴宏杰张宇昊
食品工业科技 2024年6期
黄欣娅,郭 婷,2,3,4,戴宏杰,2,4,张宇昊,2,4,马 良,2,3,4,
(1.西南大学食品科学学院,重庆 400715;2.川渝共建特色食品重庆市重点实验室,重庆 400715;3.农业农村部柑橘类果品质量安全重点实验室,重庆 400715;4.发光分析和分子传感教育部重点实验室,重庆 400715)
柑橘属于荟香科植物,是我国种植面积最大、产量最高的水果。柑橘果实鲜美可口,富含丰富的维生素C,还含有多酚、黄酮、类柠檬苦素、辛弗林等许多有益于人类健康的活性成分,深受消费者喜爱[1]。据世界柑橘组织数据显示,自2021 年夏到2021 年冬,我国柑橘产量达到4460 万吨,占全球柑橘产量的28%,居全球第一。柑橘在我国有四千多年的种植历史,截至目前,已有20 多种不同的柑橘采后病害被报道,这些病害导致柑橘大量腐烂,造成了严重的经济损失和食品安全隐患。
柑橘之所以容易遭受采后病害,是因为我国柑橘主要以鲜销为主,在采收、运输、包装和贮藏过程中都可能产生机械损伤,使得病害大量发生,以青霉病为主[2-4]。但目前我国对于柑橘病害的防治重点还专注于采前的砂皮病、黑斑病、炭疽病、疮痂病等[5-6],对于采后青霉病的防治还没有足够的重视。柑橘的种植地主要分布在中国的长江中上游、湘南赣南桂北、鄂西和浙-闽-粤四个区域。长江中上游柑橘带,高湿多热,属于典型的亚热带季风湿润气候,年平均温度达到17.5~18.5 ℃,适合青霉菌生长(18~26 ℃最适)。宜昌地区柑橘携带微生物种类分析结果显示,其致病真菌主要是青霉属真菌[7]。此外,14 个主要柑橘产区采后病害分析发现,广东的沙糖桔由青霉病引起的腐烂达到总腐烂率的73%,宽皮柑橘感染青霉病的频率较其他品种高[2]。现有调查发现,由于健康的柑橘pH 呈酸性,柑橘更容易被真菌感染而发生腐烂[8],其中指状青霉(Penicillium digitatum,P.digitatum)和意大利青霉(Penicillium italicum,P.italicum)造成的采后损失分别高达90%和30%,而一般情况下果实腐烂率在10%~30%左右[9]。
柑橘是我国优势水果产业和长江库区农业增收、农民致富的支柱产业,对推动地方“乡村振兴”和“质量兴农”具有重要意义[2]。本文归纳了感染柑橘的三种青霉菌的侵染过程、产毒机制及其危害,分析其中的共性与差异,概述了近年来新兴的青霉病控制方法,为广泛探索柑橘收获后植物病原体的新控制方法,从而发现有前景的新方法提供参考,对促进农业经济产业、保障地区食品安全有重大意义。
1 青霉菌侵染机制及产毒机制
在农业生产中,真菌毒素污染存在于植物生长,果实收获、加工、运输以及贮藏等各个阶段。温度、水分活度、宿主抗性等环境因素的差异,导致真菌毒素的发生情况不同。青霉菌感染柑橘果实时,要突破角质层[10]、细胞壁和水果自身的免疫系统,已有研究证实角质层的成分会对P.digitatum的菌丝体生长和分生孢子的萌发有抑制作用[11]。
青霉菌的侵染过程可以大致概括为图1 所示。附着在柑橘表面或存在于环境空气中的病原菌产生的孢子在表皮萌发后通过伤口、菌丝穿破表皮的方式侵入果肉细胞,青霉菌再通过合成有机酸降低伤口pH 增强致病性,并通过基因上调表达分泌致病因子(细胞壁降解酶、蛋白酶、乙烯合成酶等)扩张侵染范围。在这个过程中,果皮受伤会引发柑橘果实的氧化应激免疫反应,但同时挥发性物质也会从伤口处破裂的油腺中释放出来,使果实表面的休眠青霉菌孢子变得活跃并迅速发芽定植于受伤组织,促进感染过程。

图1 青霉属真菌感染柑橘果实机制Fig.1 Mechanism of Penicillium fungi infection in citrus fruits
TAO 等[12]的研究认为在柑橘受侵染的过程中,主要的挥发性物质为柠檬烯,并且在侵染过程中始终保持稳定含量,否定了前人猜测柠檬烯可能参与碳源合成的假设。青霉菌还会分泌一些结构域蛋白(LysM)以及抗氧化酶如超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)抑制活性氧爆发,以抑制柑橘的免疫反应促进青霉菌的侵染。在侵染过程中真菌产生的致病因子(又称毒力因子)是植物病原体在病原体-宿主相互作用过程中用来破坏宿主和传播生化信号的次级代谢产物[13],也是霉菌毒素积累的途径。
1.1 P.digitatum 的侵染机制及产毒
P.digitatum属于专性寄生性、坏死营养型的病原真菌,只能通过果实伤口才能完成入侵,从而导致寄主细胞死亡,并且利用寄主营养完成自身生长[14]。
钱鑫[15]认为与柑橘伤口接触的最初48 h 是P.digitatum完成感染的关键时期。在这48 h 内,P.digitatum分生孢子在适宜条件下萌发,产生胚管,穿透果皮细胞,延伸至中果皮细胞,逐渐侵入周围组织,再通过宿主合成碳源,并产生柠檬酸、葡萄糖酸和富马酸,降低pH,创造细胞壁降解酶最适pH 环境[16],进一步扩展侵染范围[17]。为了获取自身三羧酸循环代谢所需的必需氨基酸,P.digitatum会分泌泛素、泛素偶连酶、蛋白酶等促进伤口蛋白的水解并软化创面。另一方面,TAO 等[12]认为在感染初期,受P.digitatum感染的影响,创面产生大量活性氧(ROS),并消耗大量抗坏血酸,使VC的含量迅速下降,P.digitatum为了避免大量ROS 对自身的影响,大量的抗氧化基因上调基因表达[18-19],除此之外萜烯柠檬烯通过调节P.digitatum孢子中的ROS 稳态诱导柑橘类水果的绿色霉菌的萌发。除了活性氧的爆发,柑橘伤口还会分泌茉莉酸和水杨酸来增加抗病性,但YANG 等[16]发现茉莉酸的合成途径可能会被P.digitatum分泌的效应因子阻断。水杨酸可以通过促进木质素在细胞壁中的积累来增加果实抗病力,但不能有效抵抗P.digitatum分泌的大量果胶酶和有机酸的攻击[20]。最终P.digitatum在48 h 后完成了代谢物积累,继续产生分生孢子并侵染整个柑橘[21]。P.digitatum在侵染过程中会产生毒素,作用于寄主果实的质膜、线粒体等细胞结构上,破坏寄主的膜完整性以及正常的代谢,加速果实衰老和感病[3]。P.digitatum与P.italicum只能产生桔霉素,缺乏合成展青霉素的完整基因[22]。基于对红曲霉产桔霉素的研究[23],目前得出的桔霉素合成步骤为:在聚酮合酶的催化下,一分子乙酰辅酶A(acetyl-CoA)与三分子丙二酰辅酶A(malonyl-CoA)通过反复缩合和延伸形成四酮体(tetraketide),四酮体与乙酰辅酶A 经甲基化缩合、氧化、脱氢、脱水等步骤生成桔霉素。目前对于桔霉素在柑橘类水果中的报道还较少[23-26],国标也未提及对于水果及其制品中桔霉素的限量值,这可能是由于桔霉素的毒性相对较弱未能引起大众的广泛重视。桔霉素已被证明具有抗菌活性[27],因此微生物之间的竞争可能是青霉菌产生桔霉素的诱因。
1.2 P.italicum 的侵染机制及产毒
P.italicum是一种坏死性植物病原体,其感染特征是白色菌丝体覆盖受感染的果实。随着白色菌丝体生长并深入受感染的组织,随后产生蓝色孢子。
虽然由P.italicum引发的蓝霉病病程发展较慢,但因为它能够直接攻击健康的水果[28],如表1 所示,与P.digitatum有明显的不同,并且由于P.italicum是筑巢型(nesting-type fungus)病原体,具有低温和低水可生长性[3],能较好适应恶劣天气,柑橘果实感染P.italicum后会造成更大的经济损失风险。WANG等[27]提到P.italicum可以由受感染水果迅速传染到与之相邻的健康水果。在感染过程中,非酶和酶因素引起的细胞壁降解是P.italicum感染砂糖橘的主要方式[28],这些致病性水解酶造成柑橘外观呈水浸状,变得柔软;GONG 等[29]研究表明P.italicum致病性与鞘脂和水杨酸信号通路有关,并且将诱导巴伦西亚橙(Citrus sinensis)果实中的碳水化合物代谢作为其感染策略的一部分,导致受感染的果皮组织失去光滑度并增加对机械损伤的易感性[32],但针对P.italicum穿透果皮感染柑橘类水果的过程仍需进一步研究。就目前来看,关于P.italicum合成桔霉素的报道较少,对于P.italicum在柑橘类水果中的产毒机制还不明确。

表1 侵染柑橘的重点青霉菌特征Table 1 Characteristics of key Penicillium which infecting citrus
1.3 扩展青霉(Penicillium expansum,P.expansum)的侵染机制及产毒
P.expansum是一种广泛存在于环境内的丝状真菌,最适生长温度在16~25 ℃,菌落呈黄绿色的小斑点状,它和P.digitatum一样都是伤口病原体[26,31]。赵利娜等[32]对感染P.expansumH1 处理的柑橘进行了转录组学分析,证实P.expansum通过分泌细胞降解酶穿透细胞壁,并产生桔霉素抵抗其它生物的威胁,降低宿主pH、增强抗氧化能力促进P.expansum的侵染。VILANOVA 等[33]检测发现感染了P.expansum的柑橘的伤口处主要有机酸是草酸和葡萄糖酸,以此给酶提供适宜的酸性环境。
P.expansum 是产展青霉素的主要菌种,具有更广泛的寄主范围[19]。不同于柑橘专一性侵染菌P.digitatum和P.italicum,P.expansum在苹果中侵染更为严重。展青霉素是一种四联醇霉菌毒素,可诱发免疫、神经和胃肠道疾病,比桔霉素毒性更强。中国规定苹果果汁中允许的展青霉素含量最高为50 μg/L,对于柑橘及其产品中毒素含量还未出台限量标准。董晓妍等[34]研究发现在苹果汁中P.expansum能产生大量毒素,并且随温度升高其生长速率和产毒速率加快,所以在柑橘类水果中同样有感染展青霉素的风险。P.expansum的展青霉素合成包含15 个基因簇,这些基因簇与P.expansum的色素沉积、菌落扩张、产孢和表性特征也有关。LI 等[19]采用基因组学分析了三种青霉菌致病性与展青霉素合成的关系,结果表明仅P.expansum有完整的展青霉素基因簇,并通过敲除合成展青霉素的关键基因(PePatK和PePatL)实验证明,展青霉素不作为致病因子促进P.expansum扩张,基于大多数产生展青霉素的真菌都不是植物病原体,但展青霉素对某些细菌和真菌具有抗菌活性可以推测出在与同环境中的微生物竞争中会促使该菌合成展青霉素,以提升竞争能力。展青霉素的生物合成包括大约10 步缩合反应和氧化还原反应,是一种聚醋酸酯衍生的次级代谢产物,由一个单位的乙酰辅酶A(CoA)和三个单位的丙二酰辅酶A 缩合而成。LI 等[35]的研究证明,在合成展青霉素的十个步骤中间,羟基苯甲醇先通过多羟基苯醇羟化酶(PatI)转化为龙胆醇,然后再转化为龙胆醛,解开了前人对于该步骤中中间体是龙胆醇还是苯甲醛的疑问,但在真菌中,展青霉素生物合成的分子调控机制以及与不同催化酶相关的各个基因在该途径生化步骤中的作用仍不清楚。
2 柑橘青霉病及其危害
2.1 柑橘青霉病发病情况
已知青霉菌侵染柑橘的方式主要是通过果实表面机械伤口入侵[7,9],有研究对武汉、宜昌、赣州三大柑橘产区的空气中微生物含量及优势菌进行了检测筛选,结果表明各产区空气中的活菌数以青绿霉最多,且三个产区优势病菌中均含有P.digitatum[8]。青霉菌生长温区为3~32 ℃,以18~26 ℃最适,相对湿度95%~98%。柑橘受真菌感染的过程如图2 所示,青霉病的发生与环境、果实和病原菌等因素有关,其中病原菌主要来源于果园、包装车间中自然存在以及贮藏环境中存在的青霉菌。环境中的病原菌除了自然存在外也可以由病果释放到空气中,这使得环境中的孢子数增加,进而提升发病率。采前环境因素造成的伤口、采摘中的机械伤口,或是储运中造成的伤口都使柑橘容易感染、传播青霉病[5],加剧青霉病感染带来的经济损失。LOUW 等[36]指出核果(如桃、杏、李等)出口季节与柑橘出口季节重叠,产品链上柑橘、梨和毛桃等具有交叉感染绿霉病的风险。
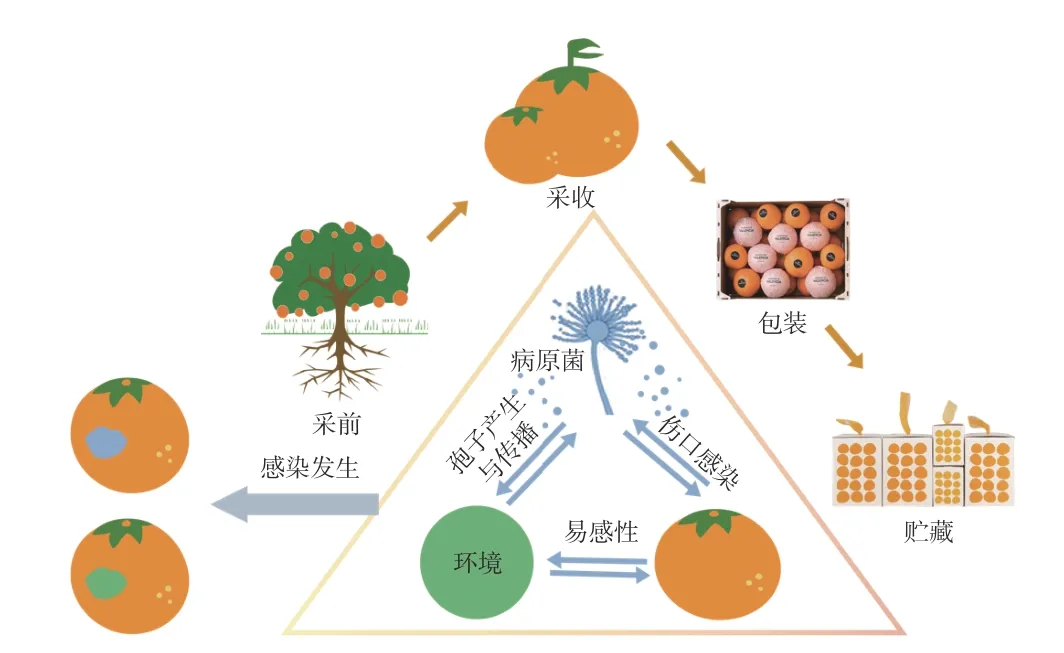
图2 柑橘真菌感染过程Fig.2 Process of citrus fungal infection
侵染柑橘的真菌中,P.digitatum引起的绿霉病、P.italicum引起的蓝霉病和P.expansum引起的青霉病都属于柑橘青霉病[37]。前两种真菌的发病率达到39.5%[38],P.expansum较前两者发病率相对较小[39]。P.digitatum引发的绿霉病是全球柑橘类水果采后危害最大的病害[40],占我国柑橘果实采后病害的80%[41],P.italicum对低温适应性强于P.digitatum,在冷藏条件下(3~5 ℃)青霉病仍然可以发生[42]。P.expansum感染果实范围更广,在其他水果中也被检出[43]。
2.2 柑橘感染青霉病对食品安全的影响
病原体感染会导致柑橘腐烂,造成柑橘质量和市场产量显著下降。除了造成的直接经济损失外,青霉菌还会产生对人类健康构成潜在风险的次级代谢产物。青霉菌属被认为是胞外分泌物(包括真菌毒素、潜在致癌物质和其他次生代谢物)的生产者[44],通常对机体的正常生长、发育和繁殖并不重要,但可以带来重要的生态优势、对人和动物有毒害作用。COSTA 等[45]指出P.digitatum侵染柑橘果实后能够产生生物碱A 和B。P.italicum侵染柑橘果实后能够产生deoxybrevianamide E、黄青霉素、脱氢短硫铵、脱氢脱氧短硫铵E 和PI-3[1,3],这些由真菌代谢产生的生物碱对哺乳动物细胞产生的细胞毒性可诱发炎症反应[46];张真真[47]发现P.expansum侵染柑橘果实后能够产生球毛壳菌素(Chaetoglobosins)和细胞松弛素(cytochalasins),它们属于细胞松弛素家族,对哺乳动物细胞是致命的,还会产生土臭素(Geosmin)、Expansolides A 和B。上述有毒代谢物还会在果实内、果实间扩散[46,48],误食病果会损害人体健康。
侵染柑橘的青霉菌还可以产生和增殖霉菌毒素,如桔霉素(CIT)、展青霉素(PAT)以及由丝状真菌产生的低分子量次级代谢物等,影响柑橘品质安全。青霉属真菌是桔霉素的生产者[49],研究发现青霉菌产生的桔霉素对人体、动物体具有致畸、致癌作用,还有肾毒性、胚胎毒性。展青霉素[50-52]主要由P.expansum产生,具神经毒性致癌性和诱变性,对人体有胃刺激性,摄入后可能会引起呕吐。毒素在水果中扩散可能渗透到腐烂部位周围几厘米处,扩散速度与水果自身以及环境有关。在植物生长阶段,真菌毒素的产生受宿主抗性、气候条件影响较大;作物收获后,农产品运输和存储环境会影响产毒真菌的发生以及真菌毒素的形成[53]。蒋黎艳等[24]人工接种宽皮柑橘,发现柑橘贮存时间较长或污染严重时,毒素会从腐烂部分向未腐烂部分渗透、扩散,导致未腐烂部分同样含有微生物的代谢物,通过简单的剔除腐烂病斑处,并不能完全有效防止毒素进入人体或加工流程中[54]。
2.3 柑橘青霉病的防治
为了减轻采后主要病害青霉病给柑橘产业带来的经济损失,柑橘果实采后常常采用一定的防治措施控制青霉病。防治措施主要包括化学和非化学的措施。其中,化学杀菌剂如咪唑、噻苯达唑、嘧霉胺和氟虫腈等[14]因方便高效成本低被广泛应用,但是化学杀菌剂可能会导致耐药真菌菌株的产生,并且存在残留问题,可能造成环境污染从而对消费者健康带来潜在危害。
近年来采用辐照[55]、天然化合物[56-59]、微生物[59]、生物防治剂[42,60]和热水处理等[61]生态友好型的生物防治、物理防治方法对柑橘果实采后病害进行防治的研究越来越多。物理防治虽然可以有效避免果实表面残留问题,但存在效果有限,作用持久力不够,成本高的局限性。目前生物防治方法因其绿色、高效的优势成为了研究热点,最新研究证明新鱼腥草素钠和淡紫灰链霉菌x33 发酵提取物对采后柑橘绿酶病菌有抑制作用[62],三峡地区特有灌木疏花水柏枝(Myricaria laxiflora)的12 株具较高抗氧化活性内生真菌可以较好抑制库区优势致腐菌橘青霉的生长[63],在其次级代谢产物中分离到的三峡肽素是一种潜在的生物抗真菌剂,对P.digitatum有较好的抑制能力。
3 结论与展望
近年来采后青霉菌造成的柑橘果实腐烂不仅给柑橘产业造成了显著的经济损失,而且引发的真菌毒素污染还会对果品安全和消费者健康产生严重威胁。目前国内研究主要集中于对P.digitatum的侵染机制以及物理化学方法防治的研究,对于柑橘其他两种青霉菌(P.italicum和P.expansum)的侵染机制及其真菌毒素的控制与降解方面的研究较少,并且对于鲜食水果的使用安全性的重视程度不够高,必须加大对柑橘水果安全的把控力度,以推进柑橘产业高质量发展,未来的研究趋势主要是绿色的生物防治方法和食品安全的控制。目前已探索出多种天然提取物对于青霉菌的抑制作用,但仍然存在成本高无法广泛投入生产以及实际应用的问题,所以应从更易获得的天然抑制剂入手,切断青霉菌的侵染、产毒进程,降低防治成本。建议可以从以下几个方面来推进各产区对柑橘青霉病防治以及真菌毒素的污染控制:研究P.italicum和P.expansum侵染与产毒机制的关系,如基因组测序、遗传转化系统的建立以及青霉菌在柑橘侵染过程中合成桔霉素和展青霉素的机制;研究将高新防治青霉病的技术与柑橘内毒素控制联系起来的联合控制办法,发掘有效抑制青霉菌和清除真菌毒素的生物拮抗菌、研究毒素降解酶等物质的表达基因等;针对感染的不同阶段开发适用的防治方法,将采前预防青霉菌污染方法与采后抑制青霉菌感染策略有机结合起来,开发联合防治方法。除此之外,对于柑橘毒素污染还应建立相应的检测标准以及毒素限量标准,进一步保障生鲜水果的食用安全。
猜你喜欢
课堂内外·初中版(科学少年)(2024年6期)2024-07-08 17:00:28
河南畜牧兽医(2021年3期)2021-01-06 04:43:52
祝您健康·文摘版(2019年10期)2019-10-14 02:07:23
天然产物研究与开发(2018年9期)2018-10-08 03:25:52
文理导航·科普童话(2016年7期)2017-02-04 20:21:18
食品与发酵工业(2016年4期)2016-05-24 09:04:20
中国当代医药(2015年26期)2015-03-01 02:06:51
现代检验医学杂志(2015年4期)2015-02-06 02:02:12
现代检验医学杂志(2015年2期)2015-02-06 02:00:51
食品工业科技(2014年15期)2014-03-11 18:17:42
